
Abstract
Cell-mediated immune response is critical for Mycobacterium tuberculosis (M.tb) control. Understanding of pathophysiology and role played by different cell mediators is essential for vaccine development and better management of patients with M.tb. A complex array of cytokines and chemokines are involved in the immune response against M.tb; however, their relative contribution in protection remains to be further explored. The purpose of this review is to summarize the current understanding regarding the cytokine and chemokine profiles in M.tb infection in order to assist research in the field to pursue new direction in prevention and control. We have also summarized recent findings on vaccine trials that have been developed and or are under trials that are targeting these molecules.
Graphical Abstract
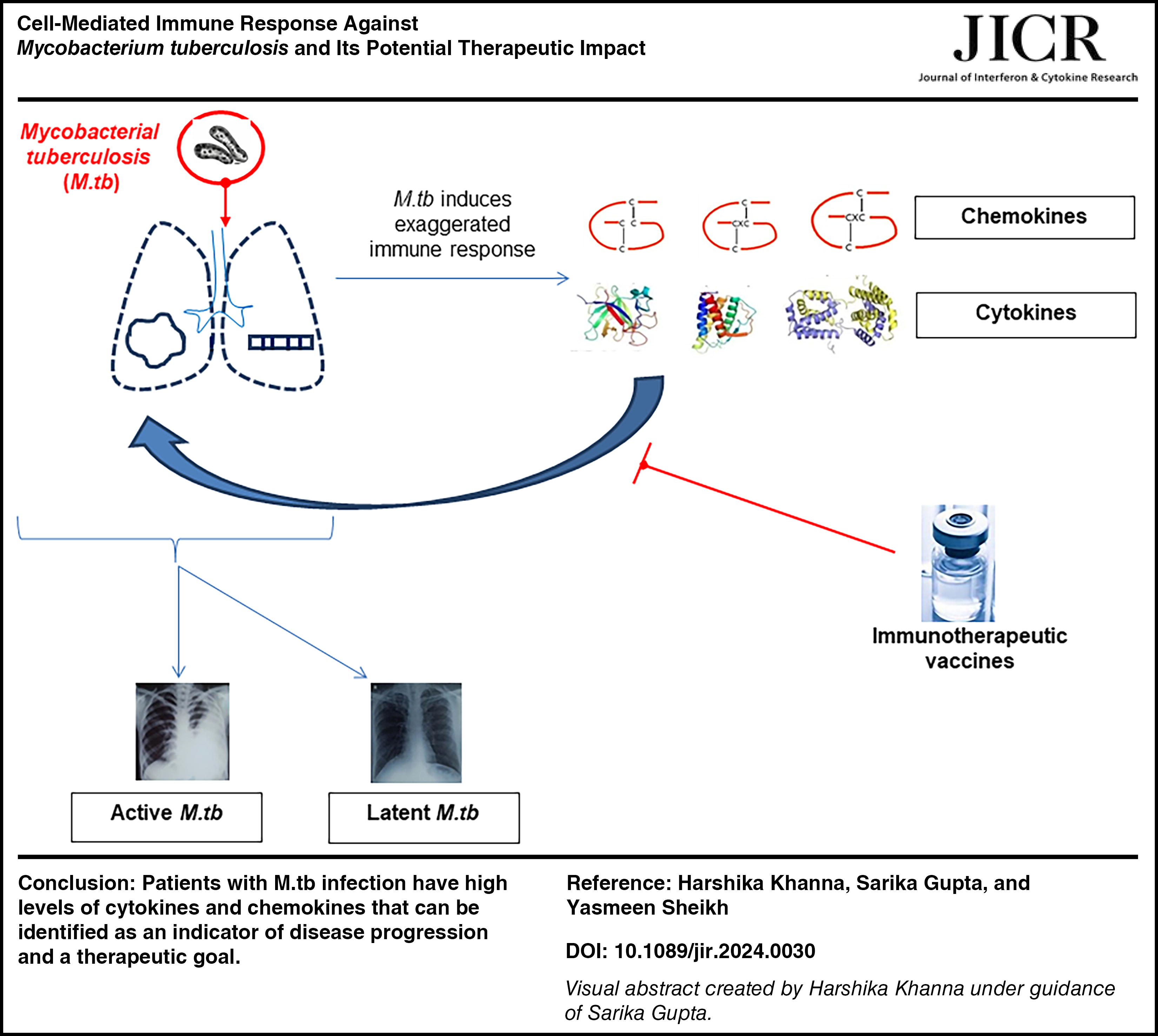
Introduction
M.tb is the 13th leading cause of death and the second leading infectious killer after COVID-19. As per the WHO report, 1.2 million children fell ill with M.tb across the world in 2021. India notified 1.9 million M.tb cases in 2021, out of which 6% were represented by children of age between 0 and 14 years (Kanabus, 2022). M.tb predominantly affects the lungs and accounts for 70% of the cases with pulmonary M.tb; however, it can disseminate to other organs too, including lymph nodes, bone, and meninges, and cause extrapulmonary disease (O’Garra et al., 2013). Even though M.tb is a curable disease, diagnostic methods for M.tb are imperfect and perform poorly in children because of difficulties in obtaining samples and low bacillary loads. Moreover, diagnosis of active M.tb in children poses a significant dilemma because clinical symptoms of M.tb in children are nonspecific and resemble other prominent pediatric disorders, including pneumonia and malnutrition, making clinical diagnosis questionable (Portevin et al., 2014). Because young children are unable to expectorate sputum, using respiratory specimens for the diagnosis of bacterial confirmation in these individuals is also challenging (Perez-Velez and Marais, 2012). Consequently, there exists an urgent necessity for a novel, nonsputum-based point-of-care (POC) diagnostic tool that could offer an early and accurate diagnosis of M.tb disease in children.
Accumulating evidence from recent clinical studies strongly indicate that cytokines and chemokines can promote the recruitment and activation of cells into M.tb-infected lungs. Particularly, T helper 2 (Th2) and Th17 cell cytokines are involved in the immunopathogenesis of M.tb by mediating alternative activation of macrophages and neutrophilic inflammation. Despite this, inflammatory markers are still largely overlooked in the management of M.tb. Neither inflammatory markers are routinely measured in patients with M.tb nor any agents targeting the inflammatory system are used in clinical practice for prevention or treatment of M.tb. Thus, with the purpose to combine basic science and clinical research to provide an updated overview of the role of cytokines and chemokines in the management of M.tb, we intend to write this review. In this review we will focus on the different cytokines of Th1, Th2, and Th17 cells as they make up the vast majority of the available data in clinical studies. We will also discuss the role of certain chemokines and their clinical relevance in management of M.tb, with potential future options.
Pathophysiology
M.tb is spread when a person who has pulmonary tuberculosis coughs, sneezes, talks, or sings; tiny airborne droplets known as droplet nuclei are produced. Transmission is influenced by the quantity of bacteria in the droplets, their virulence, their exposure to ultraviolet (UV) light, their level of ventilation, and the circumstances under which they get aerosolized (Dunlap et al., 2000). The respiratory system becomes infected when M.tb enters the lungs, but the organisms can also move to other organs such the lymphatics, pleura, bones or joints, or meninges and result in extrapulmonary M.tb (Fig. 1).

Innate immune response and its kinetic to M.tb: Alveolar macrophages which are located in the air spaces are the first cells to be infected from M.tb. This initiates a nuclear factor erythroid 2-related factor 2 (NRF2)-driven antioxidant transcriptional response leading to an impaired control of M.tb growth (Lavalett et al., 2017). Infected macrophages migrate out of the alveolar space in about 2 weeks and infect other phagocytic cells, including polymorphonuclear neutrophils (PMNs) and dendritic cells (DC). The DCs migrate from lungs toward the draining lymph node using both cytokine (IL-12p40) and chemokine (CCR2 and CCR7), in which the adaptive immune response starts by activation of naive T cells in around 4–6 weeks. Activated Th1 cells and Th17 cells contain M.tb by granuloma formation and protect the lungs from tissue damage. Th2 cells and Treg cells in contrast counter-regulate Th1 cells and allow resuscitation of M.tb, which leads to granuloma caseation and active disease. If an adaptive immune system is effective enough then the individual develops latent infection otherwise in around 8 weeks to years.
Infectious droplets that are inhaled settle throughout the airways. Alveolar macrophages, which are the most common immune effector cells in alveolar spaces, swiftly surround and absorb bacteria in droplets that enter the alveoli without passing through the mucociliary system (Korf et al., 2006). Briefly, alveolar macrophages are the first cells that are infected by M.tb during the first 2 weeks, which move out of the alveolar space into the lung interstitium and infect additional phagocytic cells, including polymorphonuclear neutrophils (PMNs), dendritic cells (DC), and a variety of tissue-resident and recruited macrophage populations (Aiello et al., 2023). Despite the ability of PMNs to eradicate widespread microorganisms through reactive oxygen species (ROS) and neutrophil extracellular traps (NETs), it creates a permissive niche for M.tb, because M.tb is strong enough to resist ROS-mediated and NET-mediated killing of PMNs (Chandra et al., 2022). Monocyte-derived DCs are mainly responsible for transportation of M.tb to lung-draining (also known as mediastinal/pulmonary/thoracic) lymph nodes where they present the antigen to naive T cells. M.tb appears in lymph node around 9–11 days after infection from where it is disseminated to spleen. The subsequent phagocytosis by macrophages initiates a series of events that either successfully controls the infection and resulting in latent M.tb or progresses to an active disease, known as primary progressive M.tb (Frieden et al., 2003). The effectiveness of the host defenses and the equilibrium between the host defenses and the invasive mycobacteria largely affect the outcome.
CD4+ T cells and CD8+ T cells mainly dominate the host adaptive immune system, which is required to prevent the progressive M.tb (Lu et al., 2021). CD4+ T cells release IFN-γ that mainly affects macrophages by enhancing their autophagy and destruct intracellular pathogens by expressing immunity-related guanosine triphosphatase (GTPase) (Chandra et al., 2022). Subsequently, CD8+ T cells also help in controlling M.tb infection by (1) cytokine release, (2) cytotoxicity through granule-dependent exocytosis pathway, (3) cytotoxicity mediated through Fas/Fas ligand interaction, and (4) direct microbicidal activity (Lazarevic and Flynn, 2002; Volpe et al., 2016).
T regulatory cells, or Treg cells, are an important subgroup of CD4+ lymphocytes that express the markers such as CD25+ and Fork-head winged helix family transcriptional repressor p3 (Foxp3) that lead to the suppressive immunity response to self-antigens. These cells mainly produce interleukin (IL)-10 and transforming growth factor (TGF)-β1 that restrain the activity of Th1 cells and inhibit the production of IFN-γ. It has been observed in the recent studies that the number of Treg cells is higher in patients with M.tb than in healthy people, and therefore, it has been hypothesized that the Treg lymphocytes are mainly responsible for disseminated M.tb (Keikha et al., 2018). Moreover, Treg cells express a surface protein CTLA-4, which interacts with CD80 and CD86 molecules (CD80 and CD86 are costimulatory molecules that are expressed on the surface of DC) and leads to the prevention of Th1 activation and progression of M.tb in host (Keikha, 2018). Since Tregs are also highly capable of expressing some exhaustion marker genes that help in progression of M.tb disease (Pan et al., 2023), more studies are warranted in this area, which can provide new insights into T cell signatures in M.tb-exhausted populations.
Vitamin D plays a vital role in innate immunity of host against M.tb. During the interaction of M.tb with Toll-like receptors on DC, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) gets activated, which in turn upregulates the expression of vitamin D receptor and vitamin D-1-hydroxylase genes that converts pro-vitamin D into the active form 1,25(OH)2Vit D3. This leads to generation of the antimicrobial peptides cathelicidin and β-defensin to kill intracellular mycobacteria (Fig. 2). In addition, vitamin D also regulates the expression of human β defensin 2, another antimicrobial peptide produced by epithelial cells, CD8+ T cell, and natural killer cell-derived granulysin (Kaufmann et al., 2014). Furthermore, calcitriol (1,25-(OH)2-Vit D3, 1,25-(OH)-Vit D3) is a hormonal agonist of IFN-γ induced autophagy through the activation of AMPK (AMP-activated protein kinase) (Deretic, 2014).

Vitamin D-mediated immune response against M.tb.
Diagnosis of M.tb
Recent researches have provided many diagnostic tests and tools, but none of them have reliable sensitivity and specificity to be able to consider as a gold standard test for diagnosis. Pediatric M.tb is paucibacillary in nature due to which its accurate diagnosis is difficult. M.tb diagnosis in young children mainly depends on contact history and clinical and radiological features. Table 1 describes the recent tests that are evaluated for their sensitivity and specificity. Despite promising developments, there are currently no reliable diagnostics for pediatric M.tb. As previously stated, the percentage of children with active M.tb who were misdiagnosed by current diagnostic tests is 40% by culture, 77% by microscopy, and 50% by gene Xpert (Schumacher et al., 2016). Because of the low reliability of current pediatric diagnostics, the diagnosis of childhood M.tb is almost entirely based on medical history, clinical symptoms, TST results, and chest radiography.
Recent Developments in Diagnostic Tests for M.tb with Their Limitations
Role of CD4+ T Cells and CD8+ T Cells
It has been shown in previous literature that differentiation state of Th1 cells has a profound effect on their ability to migrate into the lung and therefore mediate control of M.tb infection. It is already known that interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and IL-2 are the key components of immune response against M.tb. They alone however are not sufficient to provide protection, and a complex array of T cells is involved. T-betdimCXCR3+ CD4+ T cells, a subset of CD4+ T cells, rapidly migrate to infected lungs, whereas another subset resembling terminal effectors (T-beTBrightKLRG1+) and coexpressing CX3CR1 poorly migrate into the lungs and do not effectively help in controlling the infection caused by M.tb (Sallin et al., 2017). Thus, the number of circulating Th1 cells does not correlate with the protection against M.tb. Similarly, there is growing evidence that programmed cell death protein 1, also known as PD-1+, is more proliferative than terminally differentiated Killer cell lectin-like receptor G1 (KLRG1)+ Th1 cells. KLRG1 was earlier targeted by Bacille Calmette-Guérin (BCG) vaccine, as it was earlier known to produce more proinflammatory cytokines, such as IFN-γ and TNF-α. Recently, Moguche et al. (2015) have explained that a subset of PD-1+ is maintained in lung parenchyma during M.tb infection, which, when transferred to uninfected animal, provides better protection against by elevating T cell receptor signalling (Moguche et al., 2015). This potential role of PD-1+ cells that work similar to memory CD4+ T cells makes it a better candidate for vaccine development.
CD153, a molecule belonging to TNF-α superfamily, is also expressed by CD4+ T cells. In the animal studies, it was observed that CD153+ M.tb-specific CD4 response inversely correlated with lung bacterial load and antitubercular treatment partially restored CD153 expression on M.tb-specific CD4+ T cells. However, its role in humans is still not clear, and it was anticipated that its measurement may be useful to evaluate M.tb vaccine efficacy (Du Bruyn et al., 2021).
CD8+ T cells are generally referred to as cytotoxic T lymphocytes because of their potential to kill target cells. As discussed earlier, CD8+ T cells kill M.tb infected cells through granule-mediated function (using perforin, granzymes, and granulysin) or Fas-Fas ligand interaction to induce apoptosis or by producing granulysin which can kill M.tb directly (Lazarevic and Flynn, 2002). Like CD4+ T cells, CD8+ T cells also produce a variety of cytokines, including IFN-γ, IL-17, TNF, IL-10, IL-2, and TGF-β during M.tb infection (Table 2). CD8+ T cells also express PD-1+, T cell immunoglobulin and mucin domain-containing protein 3 (Tim-3), or CD122, which helps in increasing the susceptibility to chronic M.tb infection (Lin and Flynn, 2015). CD8+ T cells that express Tim 3 protein are not only capable of producing IFN-γ, IL-2, IL-22, IL-4, and IL-17A but also they were reported to function as anti-M.tb effector cells limiting intracellular M.tb growth (Qiu et al., 2012).
Cytokines Involved in M.tb and Their Mechanism
↑ upregulates; ↓ downregulates; ≠ inhibits.
IFN-γ
IFN-γ is the primary cytokine involved in the immune response to Mycobacteria, and its primary function is to activate macrophages, allowing them to carry out their microbicidal functions. Together with IL-12 and IL-6, IFN-γ promotes oxidative burst, which aids macrophages in their tuberculostatic function. It also promotes immune cell migration to the site of infection, which aids in the creation of granulomas, which slows the progression of the disease (Cooper and Khader, 2008) (Fig. 1). IFN-γ is a significant cytokine that functions in various capacities across the immune system. The ability of macrophages to destroy Mycobacteria through pinocytosis, receptor-mediated phagocytosis, and microbial death was boosted when IFN-γ activates them (Cooper, 2009). Type I IFNs, which are produced by fibroblasts and leukocytes, respectively, are stimulated and act efficiently in responses against viruses. Type II interferon, or IFN-γ, is mostly produced by T lymphocytes and NK cells in response to immunological and inflammatory stimuli, as opposed to being produced by viral infection (Gehring et al., 2003). More than 200 genes in macrophages are stimulated by IFN-γ, including those that produce antimicrobial compounds such as oxygen free radicals and nitric oxide, which are among the most effective effector mechanisms for killing M.tb (Cavalcanti et al., 2012).
IL-12
IL-12p40 is a subunit of IL-12 and IL-23, deficiency of which is predisposing the host to M.tb by altering the expression of NF-kB essential modulator (which regulates T cell-dependent and CD40L-triggered IL-12 production and IFN-γ production to mediate protection against Mycobacterial infections) and Chemokine (C-C motif) ligand 2 (CCL2) (Cooper et al., 2007). IL-12 receptor expression also helps in maturation of Th1 cytokine (IL-2 and IFN-γ) producing phenotype among T cells, generation of granulomas, and reduces the levels of IFN-γ–dependent chemokines such as RANTES, which is specific for activated or memory T cells (Cooper et al., 1997; Sampath et al., 2023a).
IL-2
IL-2 mainly regulates the differentiation of T cells and promotes the formation of CD8+ T cells. IL-2 also mediates vasodilatory effects and induces the chemokines to recruit leukocytes at the reaction site (Sakamoto, 2012). It has been also observed that during chronic M.tb infection antigen-specific T cells expressed high level of PD-1 and had limited capacity to produce IL-2. Many studies have shown that when combined with antibacterial drugs, IL-2 treatment against drug-resistant M.tb resulted in a higher rate of sputum conversion compared with drug-only controls (Liu et al., 2019; Zhang et al., 2012).
TNF-α
TNF-α is a cytokine that promotes inflammation and has a variety of biological consequences. TNF-α works in concert with IFN-γ to promote the synthesis of reactive nitrogen intermediates (RNIs), which in turn helps macrophages perform their tuberculostatic function. TNF-α also promotes immune cell migration to the infection site, which helps to produce granulomas that can slow the spread of the disease (Dambuza et al., 2008). The capacity of Mycobacteria to cause persistent infections was described as being accompanied by a decrease in TNF production in human peripheral blood mononuclear cells (PBMCs) (Manca et al., 2004). TNF-α, which is produced by macrophages, lymphocytes, neutrophils, and certain endothelial cells, orchestrates the inflammatory response by inducing other cytokines (IL-1 and IL-6), and it attracts immunological and inflammatory cells by inducing chemokines and upregulating adhesion molecules. According to experimental models, TNF-α is crucial for both the immunopathology of M.tb and the host response to M.tb (van Kooyk and Geijtenbeek, 2003). TNF-α enhances the ability of macrophages to phagocytose and destroy mycobacteria and stimulates macrophage apoptosis, which deprives bacilli of host cells and causes death, as well as the presentation of mycobacterial antigens by DC.
IL-4
Th2 lymphocytes and activated mast cells both produce IL-4. In addition to acting as an autocrine growth factor for Th2 cells and inhibiting the formation of Th1 and Th17 lymphocytes, this cytokine also promotes the isotype change of IgG4 and IgE and contributes to the activation of macrophages (Ordway et al., 2004; Rook et al., 2004). IL-4 has been proposed as a major player in the pathogenesis of M.tb by inhibiting inducible nitric oxide synthase, Toll-like receptor 2, and macrophage activation (Sugawara et al., 2000). Recent studies have shown a role of IL-4Rα-driven T helper 2 immunity in the progression of the disease as its secretion in peripheral blood mononuclear cell is elevated and involved in cavitary granuloma formation in patients with active M.tb disease (Parihar et al., 2021). Studies also indicated that downmodulation of IL-4 is required along with elevation of IFN-γ to exhibit a prolonged Th1 response against M.tb. This is because it was hypothesized that IL-4 suppresses macrophage derived production of IL-12, which inhibits differentiation of Th1 cells and inhibits cell mediated immune reactions by antagonizing the macrophage-activating effect of IFN-γ (Mihret et al., 2012).
IL-5
IL-5 is secreted by CD4+ T cells and macrophages. IL-5 is earlier known to activate eosinophil, antibody production, and B cell growth and promotion (Takatsu, 2011). In M.tb infection, IL-5 functions as a regulatory cytokine and disrupts the cytokine balance that is required for proper activation of macrophages (Diedrich et al., 2013). Recent studies have suggested that overexpression of Th2 cytokines, including IL-5, increases the severity of M.tb by suppressing the Th1 cytokines (Dheda et al., 2010).
IL-6
IL-6 is mainly produced by macrophages in M.tb infection, and higher level of IL-6 was reported in having advanced pulmonary M.tb. IL-6 inhibits both IFN-γ and starvation-induced autophagy (Hamilton et al., 2023). Virulent M.tb upregulates IL-6 production by macrophages through activation of signal transducer and activator of transcription 3 (STAT3) gene and regulates host immune response. It is also known to accelerate the growth on M.tb in peripheral blood mononucleated cells and inhibits the production of TNF-α and IL-1β, which negatively affects intracellular killing of microorganisms and development of granulomas which increases the disease severity (Buha et al., 2019).
IL-9
The biology of IL-9 underwent a paradigm change recently after the discovery of Th9 cells, a subset of CD4+ T cells that secrete IL-9, as earlier it was considered to be merely another Th2 cytokine (Ye et al., 2012). It has been observed that IL-9 expression impairs IFN-γ production by inhibition of Treg cells and is helpful in discriminating adult M.tb from latent TB infection (Wu et al., 2008). Li et al. have recently showed that the expression of IL-9 is significantly increased in children suffering from M.tb than in normal patients that makes it a potential biomarker for childhood M.tb diagnosis in high endemic settings (Li et al., 2015).
IL-10
IL-10 was initially referred to as a cytokine synthesis inhibitory factor (CSIF) because of its capacity to reduce the cytokine output from T lymphocytes. Several studies have investigated the role of IL-10 in M.tb infections, but the cumulative results are inconclusive. Elevated levels of IL-10 in patients have been suggested to be associated with more active or severe cases of M.tb, and blocking IL-10 improved Th1 and macrophage function in in vitro studies (Shin et al., 2005). IL-10 is generated during M.tb infection by macrophages and T cells. In contrast to TNFα and IFN- γ, IL-10 is primarily thought of as an inhibitory cytokine, which is crucial for maintaining a proper balance between inflammatory and immunopathological responses. The mycobacterial survival in the host, however, seems to be supported by the rise in IL-10 levels. Samples taken from patients with M.tb have greater levels of IL-10, and higher levels of IL-10 production are linked to higher illness incidence. The fact that IL-10 synthesis is higher in anergic patients with human M.tb suggests that M.tb stimulates IL-10 production, which suppresses an efficient immune response (Gao et al., 2011).
IL-13
IL-13 and IL-4 both are known to induce immunoglobulin (Ig)E and IgG production from B lymphocytes and augment antigen load capacity by increasing the cell surface molecules, including major histocompatibility complex (MHC) on B-lymphocytes and macrophages (Abedini et al., 2020). IL-13 expression increases in M.tb and correlates with disease severity. It increases the expression of arginase and downregulates TNF-α, IL-6, and IL-1 with concomitant upregulation of the IL-1 decoy receptor and the IL-1 receptor antagonist (IL-1RA) that oppose the actions of IL-1. All of these effects are the opposite of those brought on by IFN-γ (Rook, 2007).
IL-18
IL-18 is a potent immunoregulatory cytokine that regulates the mechanisms of both innate and adaptive immunity. It is secreted by monocytes, macrophages, DC, epithelial cells, keratinocytes, and synovial fibroblasts, as well as T and B lymphocytes. Since it has a capability to induce IFN-γ in T cells, it is considered as a member of Th1 family. However, IL-18 requires the presence of IL-12 to induce IFNγ production from Th1 cells. IL-18 upregulates the expression of IL-12Rß2 which is required for the efficient production of IFNγ from T cells (Robert and Miossec, 2021). It is known to promote activation of NK cells, polarizes T lymphocytes, enhances CD8+ T cell cytolytic activity, and induces the synthesis of nitric oxide (NO) and cytokines and chemokines (TNF-α, GM-CSF, IL-4, IL-5, IL-9, IL-13, IL-17) (Wawrocki et al., 2020).
IL-22
IL-22 belongs to the family of IL-10 whose role in M.tb is very controversial. Some studies showed an increased level of IL-22 in active patients with M.tb and suggest a pathological role of this cytokine (Matthews et al., 2011). However, recent studies have demonstrated a protective role of IL-22 in patients with M.tb by observing a higher level of IL-22 in serum of latent patients with M.tb and those on antitubercular treatment. Furthermore, a single nucleotide polymorphism (SNP) in the promoter region of the IL-22 gene (rs2227473) that increases IL-22 production was observed to be associated with reduced susceptibility to M.tb (Treerat et al., 2017).
IL-27
IL-27 is a heterodimeric cytokine of the IL-12 family, which is secreted by macrophages and DC. Its main function involves inhibition of anti-M.tb T cell effector functions and phagosomal acidification and compromises control of M.tb growth by macrophages. Apart from suppressing the production of antimycobacterial cytokines TNFα, IL-6, IL-1β, and IL-15, it limits the antimycobacterial activity of macrophages by reducing Cyp27B, cathelicidin (LL-37), and LC3B lipidation and increases IL-10 production. Recent researches have shown that neutralizing both IL-27 and IL-10 increased the expression of proteins involved in LC3-associated phagocytosis (LAP) pathway for bacterial clearance (Gollnick et al., 2023).
Chemokines
Chemokines (or chemotactic cytokines) are a broad family of proteins that communicates through heptahelical chemokine receptors on cell surfaces. Chemokines are classified into four major subfamilies: CXC, CC, C, and CX3C. Their corresponding receptors are CXCR, CCR, CR, and CX3CR1 (Zhou et al., 2022). They are mainly known for their capacity to promote cell migration, particularly that of leukocytes, regulating inflammation, angiogenesis, and antimicrobial immunity. By virtue of their chemotactic activities, chemokine serves as the catalyst for both innate and adaptive immune responses (Hughes and Nibbs, 2018). Similar to cytokines, chemokines also govern the containment of M.tb through immune cell recruitment, granuloma formation, DC migration, and priming of T cells. The catastrophic effects of M.tb are thought to be caused by neutrophil buildup with increased chemokine secretion and matrix metalloproteinases (MMP), which leads to decreased chemokine activity that causes lung cavitation (Sampath et al., 2023b).
As discussed earlier, M.tb enters the lower respiratory tract by inhalation and is ingested into the lung and taken up by alveolar macrophages. The infected macrophages actively release chemokines and cytokines that cause a number of immune cell types to be attracted and activated to the lung such as neutrophils, NK cells, T cells, and macrophages and later accumulation of adaptive effector cells such as CD4+ and CD8+ T cells (Slight and Khader, 2013). It has been suggested that the chemokines CXCL8, CXCL9, CXCL10, CXCL11, and CXCL13 control inflammation and immune response to M.tb through the migration of leukocytes, including neutrophils, monocytes, and macrophages. It has been observed that as early as day 12 following infection, increased expression of the chemokines CXCL-3 and CXCL-5 is shown in particular (Nouailles et al., 2014), and this is related to the early entry of NK cells and neutrophils, both of which likely express the CXCR2 receptor. Lung epithelial cells also have the ability to directly sense M.tb and secrete chemokines, which enhances the recruitment of immune cells. Human bronchial epithelial cells and a line of alveolar epithelial cells both release CCL-2 and CXCL-8 in response to M.tb stimulation (Mihret et al., 2014; Slight and Khader, 2013). Lung DC infected with M.tb enhance the expression of CCR7 and move in response to chemokines expressed within lymphoid organs to polarize T and B cells. Once activated, T cells and B cells upregulate chemokine receptors such as CXCR3, CCR5, and CCR6 and accumulate in the M.tb infected lung. Upon accumulation of immune cells in the lung, CXCL13 and CCL19 may then mediate precise spatial positioning of immune cells to create granulomas (Slight and Khader, 2013). M.tb-infected monocytes, macrophages, neutrophils, and respiratory epithelial cells produce IL-8, which attracts and activates white blood cells in normal immune response to M.tb and has been shown to be absolutely required for granuloma formation (Krupa et al., 2015). IL-8 secretion during pulmonary M.tb leads to the accumulation of neutrophils and the recruitment of T lymphocytes and monocytes. Likewise, the distinct role of chemokines during M.tb infection is summarized in Table 3.
Functions of Different Chemokines in M.tb
Cytokines from Bench to Bedside
Developing an effective vaccine or new diagnostic/prognostic tool in M.tb depends on understanding how the balance of the essential chemokines/cytokines and their receptors is maintained and how a disruption in that equilibrium can cause disease. Although antitubercular treatment (ATT) effectively cures M.tb, there are various aspects in M.tb that still remain to be managed, which include emergence of drug resistance, difficulty in diagnosing latent M.tb, longer treatment regimens which decrease patient compliance, delayed culture diagnosis, and low sensitivity assays. Hence, much effort has been exerted to understand the cellular basis of M.tb-induced immune responses, with the aim of identifying new biomarkers, diagnostic tool, and efficient vaccine and developing new therapeutic options. Table 4 describes how different interleukins are associated with tuberculosis. Cytokines and chemokines emerged as key player in adaptive immunity and inflammatory responses in the body during M.tb infection, and understanding the qualitative, quantitative, and temporal differences in cytokine expression is critical for the conquest of M.tb and for achieving the Millennium Development Goal (Goal 6; i.e., to combat HIV/AIDS, malaria, and other diseases).
Association of Different Cytokines with M.tb
In many chronic inflammatory and immune-mediated disorders, they already served as an excellent target for many biological therapeutics.
Cytokine-Based Therapies in TB
The host immune activation during TB infection highlights the need for cytokine-based therapy that can reduce the tissue or cell damage by controlling the inflammatory response, thereby strengthening the anti-M.tb defense mechanisms. Cytokines are pleiotropic proteins that elicit various pharmacological activities. Therefore, for an effective treatment of M.tb infection, a correct dose of administered cytokines along with route of administration should be considered carefully so that it must first reach its target cell, bind to the specific receptors, and, finally, activate an intact signal transduction pathway to elicit a cellular response. A randomized controlled clinical study conducted by Dawson et al. (2009) revealed that the use of an aerosolized recombinant IFN-γ (as an adjuvant) resulted in reduced bronchoalveolar lavage cytokines, rapid clearance of M.tb from the sputum, and reduced inflammatory macrophage-neutrophil alveolitis in TB patients (Dawson et al., 2009). Similarly, an open label, single-arm pilot clinical study has shown that Allogeneic Vγ9Vδ2 T-Cell therapy in Multi drug resistant (MDR) TB patients enhances the protection against tuberculosis by producing cytokines such as IFN-γ, TNF-α, and IL-17 (Liang et al, 2021). In contrast to this, a recent meta-analysis indicated that treatment with an anti-TNF agent could increase the risk of drug resistance in patients with pulmonary TB (Park et al., 2023). IL-2 is recently under trial as an adjuvant to standard chemotherapy regimen 2HRZE/4HR (NCT04766307). Recombinant IFN-γ is also in phase I/II trial for treatment of MDR tuberculosis (NCT00001407). Similarly, an active Hexose correlated compound, which increases the expression of IFN-γ and IL-2, is also being tested in tuberculosis–HIV infection along with standard regimen (NCT05100758).
Similarly, several molecules like Enbrel® (a soluble human TNFR2-Fc blocking TNF), vitamin D3 [binds to vitamin D receptor and induces antimicrobial peptide (AMP) expression], and supplementation of nebulized IFN-Y (increases CD4+ T cell response) have potential to regulate the immune response in TB. Details of such molecules can be read elsewhere (Kolloli and Subbian, 2017; Ahmed et al., 2020). The cytokine-based therapies can complement pathogen targeted approaches and have the potential to shorten TB treatment duration, prevent resistance, and reduce lung injury by modifying specific mechanisms that cause lung inflammation and matrix destruction (Yang et al., 2023). A large number of different cytokine-based therapies are in the preclinical phase, but the clinical translational models including the pharmacokinetics, biodistribution, and toxicity profiles of these cytokine-based therapies are still lacking for patients with tuberculosis (Deckers et al., 2023).
Vaccines against Tuberculosis
Vaccination against TB will probably provide the most effective interventional long-term means of preventing and controlling M.tb infection, and it has become an urgent global priority. Of more than 40 vaccines in clinical development, several have reached phase 3 testing, with interim efficacy results being already available. BCG vaccination offers partial protection for infants and young children against severe forms of TB, but it does not protect adolescents and adults, who account for the majority of TB transmission (Kwon et al., 2018). Therefore exploring the immunological correlates of M.tb will also aid in identifying new vaccine candidates or targets against TB. Data from animal studies have revealed that most of the TB vaccines induce Th17 cells that accelerate Th1 cell recruitment and increase the expression of IFN-γ, THF-α, IL-2, and IL-17. Recently many vaccines that are in the pipeline focused on increasing the expression of IFN-γ (Andersen and Scriba, 2019). Figure 3 depicts the crucial underlying mechanism of action of present vaccine candidates.
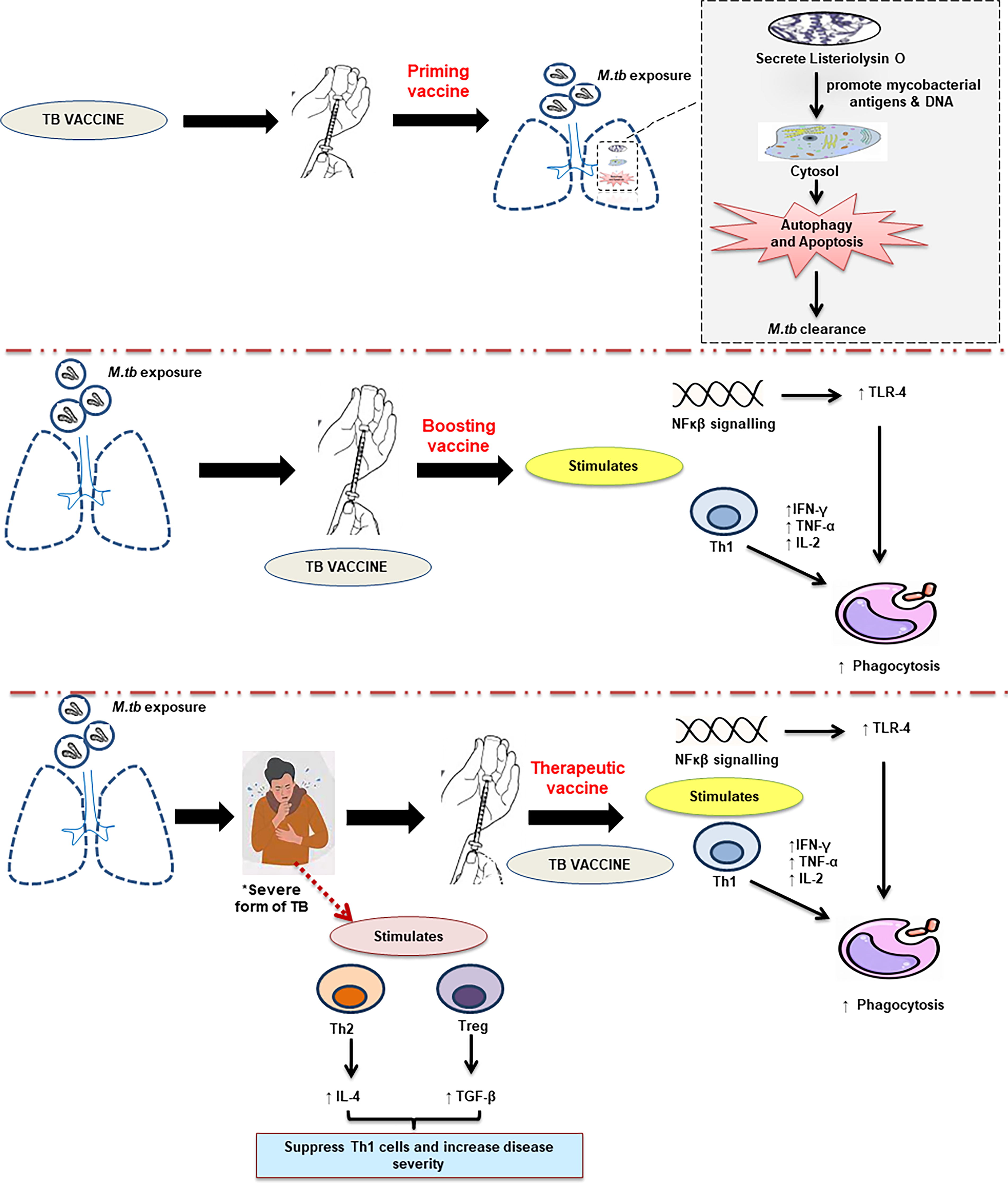
Schematic diagram of major strategies in TB vaccine development. *Target population of therapeutic vaccines in severe forms of tuberculosis like cases of MDR, XDR, or M.tb with HIV infection.
TB vaccines can be broadly categorized into two, namely, preventive vaccine and therapeutic vaccine. Preventive vaccine includes priming vaccines and boosting vaccines. Priming vaccines (preexposure) induce immune responses prior to M.tb infection such as BCG, whereas boosting vaccines that help in reinforcing the immune responses elicited after priming (Kwon et al., 2018). Currently, VPM 1002 (phase 3) and M.tbVAC (phase 1) are pursued as priming vaccines and ID93 + GLA-SE (phase 1) and M.tb72/AS02 (phase 1) are pursued as boosting vaccines against M.tb (Table 5). Therapeutic vaccines are administered in already-established disease, either to improve outcomes, shorten treatment duration, or prevent relapse (Bouzeyen and Javid, 2022). Therapeutic vaccines are now being developed based on two mechanisms, namely, bacilli-directed therapy and host-directed therapy. In the former, cellular immunity is increased to find those antigens linked to M.tb that are hidden to the immune surveillance, whereas in the latter, inflammatory responses are modulated to avoid extracellular growth of the bacilli fueled by the accumulation of neutrophils, or the Th2 response (or “Koch phenomenon”), that induce intragranulomatous necrosis (Cardona 2016). Therapeutic vaccines, for example, RUTI and DAR-901, are being developed to target the severe forms of TB in adjunct to chemotherapy, particularly in MDR TB and HIV coinfections (Kaufmann et al., 2014). Table 5 describes the summary of various M.tb vaccine candidates that are under evaluation, their composition, and their molecular mechanism. At present there are more than 20 new TB vaccines in clinical trials (Table 5), including (a) whole cell live vaccines (e.g., BCG revaccination, M.tbVAC, VPM 1002), (b) inactivated whole-cell vaccines or lysates (DAR-901, RUTI), and (c) protein subunit and adjuvant vaccines (GaM.tbvac, ID93+ GLA-SE), viral vectored vaccines (MVA85A), and DNA vaccines (GX-70).
Summary of New M.tb Vaccines under Pipeline
↑ upregulates.
One of the major challenges in development of effective TB vaccine is the selection of best antigen and how many antigens to include in a vaccine. This is because M.tb has approximately 4000 genes encoding thousands of proteins (Li et al., 2020). Identification of a reliable correlate of protection (CoP) capable of clearly identifying immune responses, associated with vaccine efficacy, is another issue. However, recent advancements in the study of immune cells or cytokines in the host’s immune protection against natural TB infection have solved this problem to an extent. Assessments of CoP in human trials is again challenging due to high costs, logistical requirements, and analytical challenges (Lai et al., 2023). Furthermore, striking a balance between longevity and efficacy is another issue as excessive induction of T effector cell vaccine modality may lead to high expression of effector cytokines, IFNγ or IL-17, that cause immunopathology (Andersen and Scriba 2019). Similarly, the lack of suitable animal model, interindividual variability observed among responders and nonresponders, and the emergence of MDR and prevalence of HIV-TB coinfection are new challenges for vaccine research.
Conclusion
In India, National Strategic Plan (2017–2025) has a national goal of elimination of M.tb by 2025. It requires increased awareness and understanding of M.tb at a molecular level, which pave the way for new vaccine/therapeutics/diagnostic tool. Patients with M.tb infection have high levels of cytokines and chemokines that can be identified as an indicator of disease progression and a therapeutic goal. Identifying the distinct cytokine profile and immune signature in M.tb has practical implications, as it can be helpful in development of vaccine and can be prognostic of clinical deterioration in M.tb patient.
Declaration of Generative AI in Scientific Writing
None.
Submission Declaration
The review article is original and has not been published anywhere.
Footnotes
Author Contributions
H.K.: Data acquisition, writing—reviewing, editing, and formatting. S.G.: Conceptualization, visualization, supervision, reviewing, and editing; Y.S.: Writing—original draft preparation, data acquisition, and formatting.
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
No funding was received for this article.