
Abstract
Recent studies have revealed that formononetin, a naturally occurring isoflavone found in kudzu root and licorice, has the potential to inhibit ferroptosis in intestinal epithelial cells. Inflammatory bowel disease (IBD) often involves oxidative stress-related pathways, making the modulation of ferroptosis a promising therapeutic avenue. We employed a combination of several techniques to explore how formononetin regulates the retinoid X receptor alpha/peroxisome proliferator activated receptor gamma (RXRA/PPARG) pathway to inhibit ferroptosis in Fetal Human Colonic Epithelial Cells (FHC) induced by RSL3. These techniques included propidium iodide staining, the levels of reactive oxygen species (ROS), Fe2+, malondialdehyde (MDA), and ferroptosis-inhibitory proteins glutathione peroxidase 4 (GPX4) and FTH analysis, Western blot analysis, and gene silencing. Our results demonstrate that formononetin significantly mitigated RSL3-induced ferroptosis as evidenced by reduced cellular levels of ROS, Fe2+, and MDA, alongside an increased expression of GPX4 and FTH. Silencing the RXRA gene reverses the protective effects of formononetin, highlighting that formononetin inhibits ferroptosis in FHC by upregulating the levels of RXRA. These findings provide new molecular targets for potential therapeutic intervention in IBD, suggesting that upregulating RXRA and PPARG expression via formononetin could be a viable strategy to mitigate ferroptosis-associated cellular damage. This could potentially lead to novel treatments for patients suffering from IBD.
Introduction
Inflammatory bowel disease (IBD) encompasses a group of gastrointestinal disorders characterized by chronic inflammation, primarily including Crohn’s disease (CD) and ulcerative colitis (UC). These diseases not only affect the digestive system but also involve multiple bodily systems, severely impacting patients’ quality of life (Dixon et al., 2012; Fakhoury et al., 2014; Wang et al., 2017). Although the precise causes of IBD remain unclear, research suggests that genetic factors, environmental influences, microbial imbalances, and abnormal immune responses may contribute to its pathogenesis (Adachi et al., 2006; Dubuquoy et al., 2006; Vandenabeele et al., 2010; Zhang et al., 2017). In recent years, researchers have shown a keen interest in the pathways of cell death in IBD, especially nonapoptotic forms of cell death such as ferroptosis (Dixon and Stockwell, 2014; Jiang et al., 2015; Liu et al., 2022). Ferroptosis is an iron-dependent form of cell death characterized by lipid peroxidation of cell membranes (Friedmann Angeli et al., 2014; Hassannia et al., 2019). In chronic inflammatory diseases such as IBD, ferroptosis could exacerbate inflammation due to the accumulation of iron ions and increased lipid peroxidation (Forrester et al., 2018; Gao et al., 2019).
Formononetin, a naturally occurring isoflavone extracted from kudzu root and licorice, exhibits significant anti-inflammatory and antioxidant properties (Chen et al., 2023; Kim et al., 2022; Li et al., 2016; Sun et al., 2016). It has demonstrated the ability to regulate cell survival across various cell types through pathways such as the NLRP3 inflammasome and other signaling pathways (Ding et al., 2024; Su et al., 2023; Wu et al., 2018; Wu et al., 2015). Moreover, its potential effects in inhibiting ferroptosis have garnered attention, particularly regarding its impact on iron-dependent cell death in model organisms (Tang et al., 2023; Zhou et al., 2023). Through the HERB database, a high-throughput experiment- and reference-guided database of traditional Chinese medicine, we identified potential targets for formononetin, peroxisome proliferator activated receptor gamma (PPARG) and retinoid X receptor alpha (RXRA). In IBD, the expression and function of nuclear receptors such as PPARG and RXRA are impaired, which are crucial in maintaining intestinal barrier function, regulating inflammatory responses, and influencing immune cell behavior (Almatroodi et al., 2023; Li et al., 2015). These receptors control cell fate by modulating antioxidative and anti-inflammatory responses (Li et al., 2021; Wang et al., 2023a).
First, RXRA and PPARG can form heterodimers and co-regulate the expression of downstream target genes. In IBD, reduced expression of RXRA might affect the formation of these dimers, thereby impairing PPARG function. Studies have shown that when RXRA levels decrease, PPARG-mediated anti-inflammatory pathways may be inhibited, exacerbating intestinal inflammation (Sæterstad et al., 2022). Furthermore, in IBD animal models and patient samples, RXRA and PPARG typically exhibit reduced expression compared with healthy controls, correlating with inflammation severity and disease activity. Exogenous enhancement of RXRA expression has been found to restore PPARG expression and function, reducing levels of inflammatory markers (Pan et al., 2023; Sæterstad et al., 2022). Transcriptional activity assays have shown that RXRA can enhance PPARG’s transcriptional activity. Co-expression of RXRA and PPARG significantly enhances the expression of PPARG target genes (Pan et al., 2023). In IBD treatment studies, enhancing the expression or activity of RXRA to boost PPARG function has shown potential in improving intestinal barrier function and suppressing inflammatory responses. This suggests that modulating the expression and function of these receptors could provide new therapeutic strategies for IBD.
Existing research indicates that PPARG can molecularly dock with formononetin (Du, 2023). The structure of formononetin contains multiple functional groups that can form hydrogen bonds or hydrophobic interactions with residues in the ligand-binding pocket of PPARG. These properties of isoflavones make them likely to interact effectively with PPARG. Based on this, the aim of this study is to explore the potential inhibitory effects of formononetin on ferroptosis in an IBD model by modulating the expression of RXRA. In various studies, RSL3 (1S,3R-RSL3) is an inhibitor of glutathione peroxidase 4 (GPX4) (ferroptosis activator), and it has been used to induce ferroptosis (Chen et al., 2021; Chen et al., 2022; Wang et al., 2023b; Xu et al., 2020). Using an RSL3-induced ferroptosis model, this study evaluates the impact of formononetin on the expression of these 2 nuclear receptors and further explores its protective effects on intestinal epithelial cell survival. Additionally, we have used specific short hairpin RNA (shRNA) technology to silence the RXRA gene to verify the role of RXRA in the protective effects mediated by formononetin. In conclusion, the effects of formononetin may provide a scientific basis for developing new IBD treatment strategies. By studying how formononetin modulates specific molecular pathways, more effective treatment options could be offered to IBD patients.

The impact of formononetin on ferroptosis in Fetal Human Colonic Epithelial Cells (FHC).
Materials and Methods
Cell culture
Fetal Human Colonic Epithelial Cells (FHC), obtained from American Type Culture Collection (CRL-1831, USA), were used in accordance with ethical guidelines provided by ATCC. This cell line was procured under conditions that comply with the ethical standards of both the provider and our institution concerning the use of cell lines derived from human tissues. FHC were cultured in McCoy’s 5A medium (16600082, Thermo Fisher Scientific, USA) supplemented with 10% fetal bovine serum (16000044, Thermo Fisher Scientific) and 1% penicillin–streptomycin (15070063, Thermo Fisher Scientific) at a temperature of 37°C and under 5% CO2 conditions.
Drug treatment
The chemicals RSL3 (SML2234, Sigma-Aldrich, USA) and formononetin (47752, Sigma-Aldrich) were used in the experiments. The group of Control (Con) was untreated cells; FHC of RSL3 group were treated for 8 h with 20 µM RSL3 (Xu et al., 2020); the For-L group was treated with 20 µM RSL3 for 8 h followed by 25 µM formononetin (Wu et al., 2018); the For-H group underwent the same initial RSL3 treatment but was subsequently treated with 50 µM formononetin; the For group received identical treatment to the For-H group; the For+short hairpin negative control (shNC) group involved FHC transfected with shNC vector and then treated with 20 µM RSL3 for 8 h, followed by 50 µM formononetin; similarly, the For+shRXRA group involved FHC transfected with pLKO.1-shRXRA and then subjected to the same RSL3 and formononetin regimen.
Propidium iodide staining for cell death detection
Post-drug treatment, cells were collected and resuspended in phosphate buffered saline (PBS, A610100-0001, Sangon Biotech, China). For apoptosis detection, cells were stained with propidium iodide (PI) (P3566, Thermo Fisher Scientific) at a concentration of 5 µg/mL for 10 min to label late apoptotic or necrotic cells. This step was followed by 4′,6-Diamidino-2′-phenylindole (DAPI, D1306, Thermo Fisher Scientific) staining to counterstain the nuclei. After washing off excess stain with PBS, cells were observed with an EVOS FL Auto 2 Microscope (AMF5000, Thermo Fisher Scientific). The microscopy was conducted at a magnification of 200× and scale of 100 μm.
Reactive oxygen species measurement
To assess the levels of reactive oxygen species (ROS) in FHC, we used the C11-BODIPY 581/591 staining method. Cells were incubated with C11-BODIPY 581/591 dye (D3861, Thermo Fisher Scientific) at a final concentration of 10 μM for 30 min at 37°C in the dark. After incubation, cells were washed with PBS and then observed with EVOS FL Auto 2 Microscope. The microscopy was conducted at a magnification of 200× and scale of 100 μm.
Fe2+ measurement
Fe2+ levels were determined using the Iron Assay Kit (ab83366, Abcam, UK). Briefly, cells were lysed and homogenized in iron release reagent, allowing for the release of Fe2+ ions into the solution. Subsequently, an iron probe provided in the kit was added, which reacts with the Fe2+ to produce a colored complex. The absorbance of this complex was measured at 593 nm using a microplate reader, which correlates directly with the iron concentration in the sample.
Malondialdehyde measurement
Lipid peroxidation levels in FHC were evaluated by measuring MDA content using an MDA Assay Kit (MAK085, Sigma-Aldrich). Cells were harvested, washed with PBS, lysed, and reacted with thiobarbituric acid (TBA) and then heated to 95°C for 60 min, with the absorbance of the MDA-TBA complex measured at 532 nm to quantify MDA concentration.
Protein expression analysis
Protein levels of GPX4 (17 kDa), FTH1 (21 kDa), RXRA (54 kDa), PPARG (58 kDa), and internal reference protein GAPDH (37 kDa) were assessed using Western blot technique. Cells were lysed in radioimmunoprecipitation assay (RIPA) buffer containing protease and phosphatase inhibitors (89900, Thermo Fisher Scientific) to preserve protein integrity. Following cell lysis, the lysates were centrifuged at 14,000g for 15 min at 4°C, and the supernatant containing the protein extract was collected. Protein concentrations were determined using a BCA Protein Assay Kit (23225, Pierce Biotechnology, USA). Equal amounts of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE), transferred to polyvinylidene fluoride (PVDF) membranes, and probed with specific primary antibodies against GPX4 (ab125066, Abcam, UK), FTH1 (ab75972, Abcam), RXRA (ab125001, Abcam), PPARG (ab272718, Abcam), and GAPDH (ab8245, Abcam), the incubation with primary antibodies was conducted overnight at 4°C. Following primary antibody incubation, membranes were washed and incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies (ab205718, Abcam) for GPX4, FTH1, RXRA, PPARG, and Goat Anti-Mouse IgG H&L (ab150113, Abcam, UK) for GAPDH at room temperature for 1 h. Chemiluminescent detection was performed using an ECL system (Amersham Pharmacia, USA) with chemiluminescence reagents from Millipore (WBKLS0500, Billerica, USA).
Cell transfection and quantitative reverse transcription polymerase chain reaction
FHC were transfected with the shRNA vector targeting RXRA, pLKO.1-shRXRA. This plasmid carries a sequence-specific shRNA designed to silence the RXRA gene. The sequence of the shRNA used for silencing is 5′-GCCAGTTATGATGAGATACAA-3′. Transfections were performed using Lipofectamine 2000 reagent (11668019, Invitrogen, USA) following the manufacturer’s protocol. Cells were incubated with the DNA-Lipofectamine 2000 complexes for 6 h, after which the medium was replaced with fresh growth medium. Forty-eight hours post-transfection, total RNA was extracted from the cells using the RNeasy Mini Kit (74104, Qiagen, Germany) according to the manufacturer’s instructions.
One microgram of the extracted RNA was reverse transcribed to cDNA using the High-Capacity cDNA Reverse Transcription Kit (4368814, Applied Biosystems, USA). For quantitative reverse transcription polymerase chain reaction (qRT-PCR), specific primers for RXRA (forward: 5′-CAAGAGGCGAACAGTGGTCT-3′, reverse: 5′-CGGAACATCTCCACCAGCAT-3′) and the reference gene GAPDH (forward: 5′-TGCACCACCAACTGCTTAGC-3′, reverse: 5′-GGCATGGACTGTGGTCATGAG-3′) were used. The qRT-PCR reactions were performed on an ABI 7500 Fast Real-Time PCR system (Applied Biosystems) using PowerUp SYBR Green Master Mix (A25742, Applied Biosystems). The cycling conditions were 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. The relative gene expression levels were calculated using the 2^-ΔΔCT method, normalizing RXRA expression to GAPDH and comparing it with control samples.
Statistical analysis
Data were analyzed using GraphPad Prism 8.0 software (GraphPad Software, USA). Multiple group comparisons were made using one-way analysis of variance followed by Tukey’s post hoc test. Statistical significance was set at P < 0.05. Results were expressed as mean ± standard deviation.
Results
The impact of formononetin on ferroptosis in FHC
Formononetin’s chemical structure is composed of 3 benzene rings: the first ring bears 2 methoxy groups (-OCH3) and 1 hydroxyl group. The central benzene ring is fused with a 6-membered ring that contains a ketone group (=O). A methyl ester group (-COOCH3) is attached at the para position of the third benzene ring. This unique compound structure potentially endows formononetin with antioxidative and anti-inflammatory properties (Fig. 1A). We systematically evaluated the effect of formononetin on ferroptosis in FHC. Previous studies have shown that 20 µM RSL3 induces death in FHC within 8 h (Xu et al., 2020). Based on this, we treated FHC induced to die by RSL3 with 25 µM and 50 µM formononetin and observed the cellular death, ROS content, Fe2+ concentration, MDA levels, and the expression of ferroptosis-related proteins GPX4 and FTH (Wu et al., 2018). Following PI staining, it was observed that compared with the Con group, RSL3 significantly induced apoptosis in FHC (P < 0.001, Fig. 1B). However, treatment with formononetin significantly reduced cell death, and higher concentrations of formononetin were more effective in reducing RSL3-induced apoptosis than lower concentrations (P < 0.05, Fig. 1B). Additionally, RSL3 increased intracellular levels of ROS (P < 0.001, Fig. 1C), Fe2+ (P < 0.01, Fig. 1D), and MDA (P < 0.05, Fig. 1E). Conversely, formononetin treatment resulted in a notable reduction in the levels of ROS, Fe2+, and MDA, with higher concentrations of formononetin proving to be more effective (P < 0.05, Figs. 1C–E). Lastly, Western blot analysis revealed that RSL3 downregulated the expression levels of the GPX4 and FTH within the cells (P < 0.001, Fig. 1F). Post formononetin treatment, however, these protein levels were significantly elevated, and high concentrations of formononetin even restored the levels of GPX4 and FTH to those of the Con group (P < 0.01, Fig. 1F). This series of experiments shows that formononetin significantly reduces cell death rates induced by RSL3 through impacting multiple intracellular signaling and metabolic pathways, effectively inhibiting iron-dependent cell death.
Protective effects of formononetin mediated by the RXRA/PPARG regulatory axis
To further explore the molecular mechanisms of formononetin in the prevention and treatment of inflammatory diseases, we utilized Western blot technology to delve into the regulatory effects of formononetin on the expression of RXRA and PPARG proteins. The results indicated that RSL3 significantly reduced the expression of RXRA and PPARG, whereas treatment with formononetin notably upregulated the levels of both RXRA and PPARG (P < 0.001, Fig. 2A). To confirm the mechanism of action of RXRA, we employed gene transfection techniques. The results indicate that after transfecting the silencing gene for RXRA, the levels of RXRA in FHC were significantly reduced compared with the shNC group, demonstrating the success of the transfection (P < 0.001, Fig. 2B). It was by silencing the RXRA gene that we were able to analyze its specific role in the protective effects mediated by formononetin through a series of experiments.
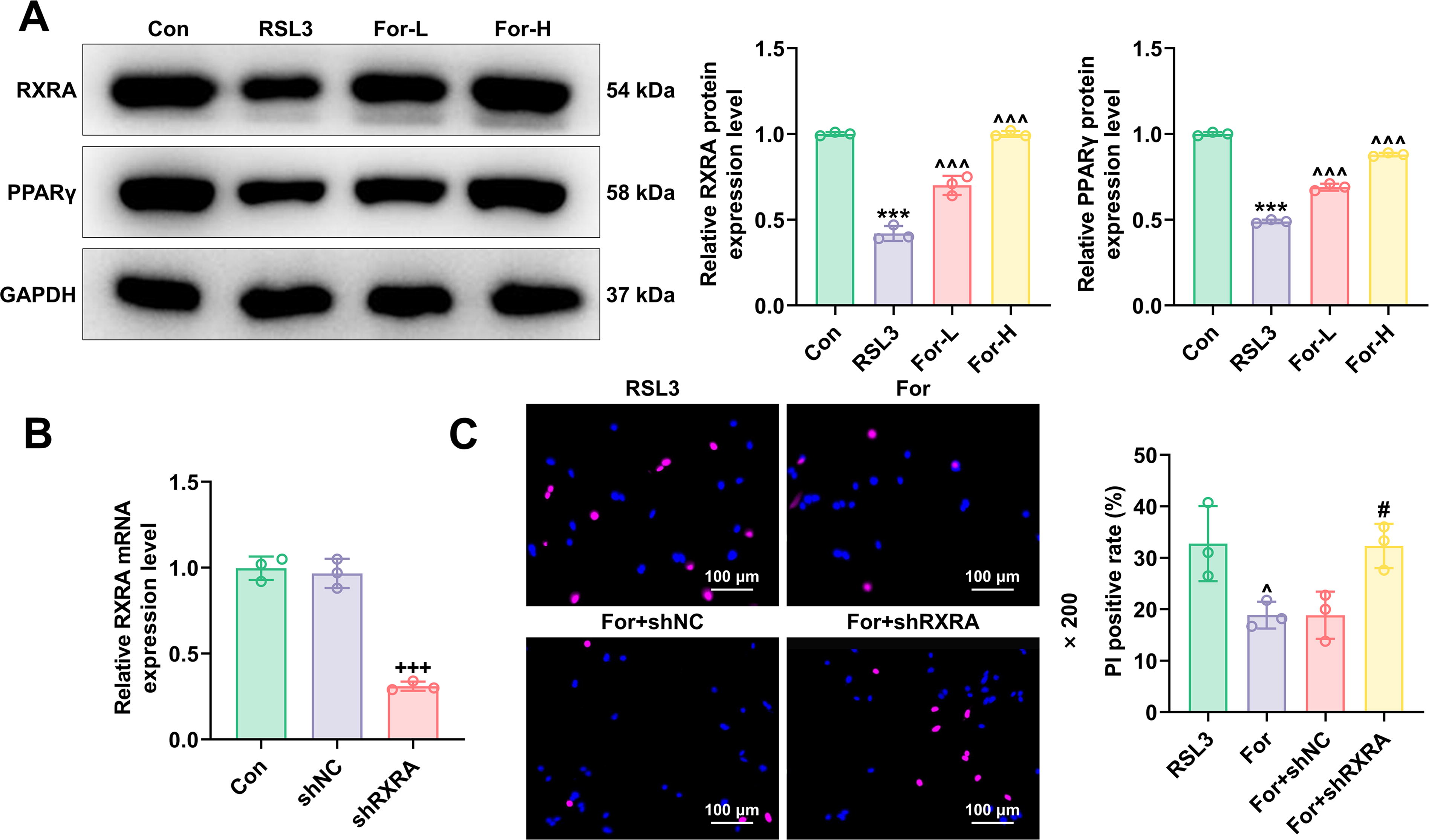
Protective effects of formononetin mediated by the RXRA/PPARG regulatory axis.
Analysis of the RXRA/PPARG signaling axis in formononetin-mediated suppression of ferroptosis
To deepen our understanding of the biological activity of formononetin, we validated the critical role of the RXRA/PPARG signaling axis in its mediation of ferroptosis suppression. Through a series of functional inhibition experiments, we detailed how formononetin regulates this signaling axis and its impact on ferroptosis. The results of PI staining showed that formononetin reduced cell death caused by RSL3, while silencing RXRA significantly reversed these effects (P < 0.05, Fig. 2C). Under the influence of formononetin, there was a significant reduction in cellular ROS levels (P < 0.05, Fig. 3A), Fe2+ concentration (P < 0.01, Fig. 3B), and MDA levels (P < 0.01, Fig. 3C), a significant increase in the expression levels of ferroptosis-related proteins (P < 0.01, Fig. 3D), RXRA, and PPARG (P < 0.001, Fig. 3E). Further experiments using shRNA technology to silence the RXRA gene, we observed that the reductions in cellular ROS (P < 0.01, Fig. 3A), Fe2+ (P < 0.01, Fig. 3B), and MDA (P < 0.01, Fig. 3C) levels achieved through treatment with formononetin were reversed. Moreover, the expression of RXRA/PPARG, which had been raised by formononetin, significantly decreased after RXRA gene knockdown (P < 0.001, Fig. 3E). This comprehensive approach clearly demonstrates that the protective effects of formononetin against ferroptosis are significantly dependent on the presence of functional RXRA. Based on this, we hypothesize that formononetin suppresses ferroptosis in intestinal epithelial cells by upregulating RXRA/PPARG.

Analysis of the RXRA/PPARG signaling axis in formononetin-mediated suppression of ferroptosis.
Discussion
This study fundamentally enhances our understanding of formononetin’s role in modulating ferroptosis through the RXRA/PPARG pathway, crucial for developing therapeutic strategies against diseases characterized by oxidative stress, such as IBD. The elevation of anti-ferroptosis proteins and the reduction in oxidative stress markers in response to formononetin highlight its potential as a therapeutic modulator, suggesting an intricate mechanism of action that extends beyond simple antioxidation.
Our study has elucidated that formononetin substantially mitigates ferroptosis in FHC, which is induced by RSL3. This modulation occurs through the upregulation of RXRA and PPARG, integral components of cellular defense mechanisms against oxidative stress. Significantly, formononetin not only reduces the classical markers of ferroptosis such as ROS, Fe2+, and MDA but also enhances the expression levels of GPX4 and FTH, pivotal in preventing oxidative cell death. This suggests a multifaceted role of formononetin beyond mere antioxidation, involving a strategic regulation of gene expression related to ferroptotic pathways and cellular survival. These findings highlight the therapeutic potential of formononetin in diseases such as IBD, where oxidative stress and ferroptosis contribute to disease progression and severity. This broad spectrum activity suggests that formononetin could be integral in developing treatments for diseases such as IBD, where oxidative stress and ferroptosis play critical roles in disease progression and severity.
When compared with existing treatments that primarily address symptoms rather than causes in IBD, the role of formononetin could be revolutionary, offering a mechanism-based approach to mitigate one of the underlying processes of the disease—ferroptosis. For example, studies on other natural compounds that inhibit oxidative stress often don’t show the same regulatory depth on gene expression as observed with formononetin, which potentially offers a more stable and sustained response against oxidative stress. The reversal of formononetin’s protective effects upon RXRA silencing, while illuminating, underscores a potential limitation of its therapeutic use: the effectiveness of formononetin may hinge critically on the baseline expression levels and functionality of RXRA/PPARG within cells. This dependency could influence treatment outcomes, suggesting that patient selection based on genetic profiling might be necessary to optimize therapeutic efficacy. Moreover, such findings prompt further investigation into the robustness of formononetin’s action across different biological contexts and genetic backgrounds. Future studies should also explore whether formononetin interacts synergistically or antagonistically with other treatments, particularly those already in use for conditions such as IBD. Understanding these interactions will be crucial for integrating formononetin into existing treatment paradigms and could lead to more personalized therapeutic strategies. Lastly, extended longitudinal studies are essential to monitor potential long-term effects or resistance patterns that may emerge with formononetin usage, ensuring sustained effectiveness and safety in a clinical setting.
The reliance on in vitro models in this study suggests a need for cautious interpretation when translating these findings to clinical scenarios. In vivo studies are required to confirm these results and to establish pharmacokinetic and pharmacodynamic profiles of formononetin. Furthermore, exploring formononetin’s interaction with other molecular pathways could unveil additional therapeutic targets and help in designing combination therapies. The implications of our findings are broad, with potential applications in designing preventative and therapeutic strategies for diseases beyond IBD, including various cancers and neurodegenerative diseases where ferroptosis plays a crucial role. Understanding the interplay between formononetin and the RXRA/PPARG axis could lead to biomarker-driven therapeutic approaches, enhancing the precision of treatment modalities in oxidative stress-related conditions. While promising, these results must be approached with caution, as the mechanisms observed may involve other, unexplored pathways. Future research should aim to define these pathways clearly, establish the long-term effects of formononetin, and explore its safety profile extensively.
In conclusion, this study not only broadens the horizon in the fight against diseases with an oxidative stress component but also opens up new avenues for the use of natural compounds in clinical settings, offering a hopeful perspective toward managing ferroptosis through dietary and pharmacological interventions. Further studies are necessary to move these findings from bench to bedside, ensuring that the potential of formononetin can be fully realized.
Footnotes
Authors’ Contributions
Conceptualization, Writing—original draft preparation, Writing—review and editing: H.H.; Data curation: X.X., Z.Y., F.X., and H.C.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by 2022 Hangzhou Medical and Health Technology Project (No. B20231277).