
Abstract
Lumbar disc herniation is primarily caused by intervertebral disc degeneration (IVDD). Nucleus pulposus (NP) cell dysfunction leads to pro-inflammatory cytokines secretion increase, causing extracellular matrix (ECM) degradation. ECM is essential for maintaining normal disc function. Glycoprotein (Transmembrane) Nmb (GPNMB) is strongly associated with inflammation, and its expression and effects in IVDD are unclear. We categorized 40 clinically collected IVDD samples using the magnetic resonance imaging (MRI)-based Pfirrmann grading system. GPNMB mRNA expression was notably suppressed in patients with severe IVDD compared with patients with mild IVDD. Increased GPNMB mRNA expression correlated with decreased Interleukin-6 (IL-6) expression and increased collagen type II (COL2A1) expression levels. We utilized lentivirus to overexpress GPNMB in IL-1β-induced NP cells to explore its function in IVDD. GPNMB overexpression inhibited pro-inflammatory cytokines Tumor necrosis factor-alpha and IL-6 secretion in IL-1β-induced NP cells, while anti-inflammatory cytokine IL-10 content was increased. In addition, GPNMB overexpression inhibited NP ECM degradation by decreasing ECM-degrading enzymes matrix metalloproteinases-3/13 and a disintegrin and metalloproteinase with thrombospondin motifs-4/5 in vitro. Mechanism studies revealed that GPNMB was bound to CD44, a receptor expressed on the NP cell surface. GPNMB overexpression inhibited nuclear factor-κB (NF-κB) p65 phosphorylation and nuclear translocation in vitro, possibly through CD44. In conclusion, GPNMB suppressed the expression of pro-inflammatory cytokines and ECM degradation in NP cells by inhibiting activation of NF-κB.

Introduction
Lumbar disc herniation (LDH) is a common condition that causes low back pain, radiating pain in the legs, and sensory deficits, and it mainly results from long-term intervertebral disc degeneration (IVDD) (Amin et al., 2017; Zhao et al., 2016). The intervertebral disc consists of the nucleus pulposus (NP), annulus fibrosus, and endplate cartilage (Pattappa et al., 2012). The gelatinous NP is the main component of the intervertebral disc. It is essential for maintaining the structural stability and biomechanical balance of the intervertebral disc (Adams and Roughley, 2006; Zhao et al., 2007), and NP dysfunction is considered to be the initiating factor of IVDD (Binch et al., 2021; Chou et al., 2011). The elevated pro-inflammatory cytokines Tumor necrosis factor-alpha (TNF-α), Interleukin-6 (IL-6), and IL-1β that NP cells secreted promote extracellular matrix (ECM) degradation, resulting in an imbalance of catabolic and anabolic processes, and degenerative disc disease (Liang et al., 2023; Risbud and Shapiro, 2014). Current treatments for IVDD include analgesics or surgery, which focus on alleviating symptoms and cannot cure IVDD radically (Reid et al., 2019). Nuclear factor-κB (NF-κB), a multifunctional transcription factor, is momentous for maintaining homeostasis in intervertebral disc. Activating the NF-κB pathway exacerbates local inflammatory response and ECM catabolism of cells in degenerative intervertebral disc (Elshamly et al., 2019; Wang et al., 2017; Zhang et al., 2021). The investigation of the molecular mechanism of NF-κB pathway activation is expected to alleviate IVDD and provide a reliable reference for LDH treatment.
The Glycoprotein (Transmembrane) Nmb (GPNMB) gene encodes a type I transmembrane glycoprotein, which is closely related to inflammation and neuroinflammation (Saade et al., 2021). GPNMB was found to attenuate the inflammatory response of astrocytes induced by cytokines (Neal et al., 2018). Overexpression of GPNMB exerted a protective effect on 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurodegeneration (Budge et al., 2020). GPNMB also reduced TNF-α and IL-6 expression, and increased IL-10 expression to inhibit inflammation in periodontal disease (Song and Lin, 2019). In addition, GPNMB protected against obesity-associated metabolic disorders by decreasing the inflammatory capacity of macrophages (Prabata et al., 2021). Several studies have elucidated that GPNMB levels are increased under pro-inflammatory or pathological conditions (Saade et al., 2021). However, whether GPNMB expression was altered in the NP of patients with IVDD required further investigation. Considering that inflammation played a crucial role in the development of IVDD, the above reports implied that GPNMB may be involved in the IVDD process.
It was reported that GPNMB reduced the inflammatory capacity of macrophages by inhibiting the NF-κB signaling pathway largely through binding to CD44 (Prabata et al., 2021). The overexpression of GPNMB decreased NF-κB expression in chondrocytes (Jie et al., 2025). In addition, GPNMB knockdown enhanced the expression of NF-κB in the ischemic brain (Ping et al., 2025). Considering the importance of NF-κB signaling in maintaining the homeostasis of intervertebral discs, we speculated that GPNMB could inhibit NF-κB signaling and decrease the production of pro-inflammatory factors and ECM degradation, resulting in alleviating IVDD.
In this study, we collected NP tissues from patients with herniated discs to measure GPNMB expression levels and analyze the potential relationship between GPNMB levels and certain pathophysiological factors associated with disc degeneration.
Materials and Methods
Clinical samples
Human degenerated NP tissues were obtained from 40 patients with LDH (17 females and 23 males; age 55.78 ± 12.74 years). The degree of NP tissues of IVDD was evaluated by the MRI-based Pfirrmann grading system (Pfirrmann et al., 2001). The study was conducted in accordance with the Declaration of Helsinki. Informed consent was obtained from each patient involved in the study. The ethics committee of Anhui No.2 Provincial People’s Hospital approved this study protocol.
Tissue histology
Clinical NP tissue samples were fixed in 4% paraformaldehyde and embedded in paraffin based on standard procedures. Next, the paraffin blocks were cut into 5 μm sections. For analyzing NP histopathology, the sections were dyed with hematoxylin and eosin (HE) and then observed by an optical microscope (Olympus, Tokyo, Japan).
Cell culture and treatment
Human primary NP cells (HUM-iCell-s012) and Human embryonic kidney 293T (HEK293T) cells (iCell-h237) were purchased from Cellverse (Shanghai, China). Human primary NP cells and HEK293T cells were cultured at 37°C in a humidified incubator containing 5% CO2 with the primary chondrocyte culture system (Cellverse, Shanghai, China) and Dulbecco modified Eagle medium (Servicebio, Wuhan, China), respectively. A lentiviral GPNMB overexpression vector (Lv-GPNMB-OE) was constructed by inserting the coding sequence of GPNMB into the XhoI/NotI site of Lv vector pLVX-IRES-puro. Then, HEK293T cells were co-transfected for 72 h with the Lv expression vector, pSPAX2 packaging vector, and pMD2.G envelope vector for Lv stocks. NP cells were harvested at 72 h after infection using lentivirus for subsequent testing. NP cells were treated with IL-1β (10 ng/mL) for an additional 24 h in another experiment after performing Lv infection. At the end of the experiment, cells and cell supernatants were collected for the following testing.
Cell counting kit-8 assay
The NP cells were seeded in 96-well plates at 5,000 cells/well. Cells were infected with the corresponding lentivirus for 72 h and then incubated with IL-1β (10 ng/mL) for 24 h. At the endpoint of the experiment, 10 μL of cell counting kit-8 (CCK-8) reagent (KeyGEN, Nanjing, China) was added to each well, and cells were incubated for 2 h in the incubator containing 5% CO2 at 37°C. A microplate reader (BioTek, Winooski, USA) was utilized to measure the Optical density (OD) value of each well at 450 nm.
Real-time quantitative PCR
Total RNA extraction was performed using the TRIpure reagent (BioTeke, Beijing, China) according to the manufacturer’s instructions. An All-in-One First-Strand SuperMix (Magen Biotechnology, Guangzhou, China) was used for transcribing the total RNA into cDNA. Real-time PCR was performed by amplifying 1 μL of cDNA using the SYBR GREEN (Solarbio, Beijing, China) and Taq PCR MasterMix (Solarbio, Beijing, China) on ExicyclerTM 96 (Bioneer Corporation, Daejeon, Korea). The 2−ΔΔCt method was used to determine relative mRNA expression, and the 2−ΔCt method for clinical samples. All samples were normalized to the housekeeping genes β-actin. Primer sequences are listed in Supplementary Table S1.
Co-immunoprecipitation
Radioimmunoprecipitation assay (RIPA) lysis buffer (PR20001, Proteintech, Rosemont, USA) containing 1% protease inhibitor was used to lyse NP cell samples. Cells were lysed on ice for 30 min and then centrifuged for 3 min (10,000g, 4°C). The supernatant was collected for the following experiment. The cell total protein contents were subjected using the bicinchoninic acid method (PK10026, Proteintech, Rosemont, USA). GPNMB proteins were immunoprecipitated by incubation with GPNMB antibodies and incubation buffer overnight at 4°C on a tube rotator. Eighty microliters of Protein G beads were then added to the lysates and incubated for 2 h at 4°C on a tube rotator. Next, the beads were washed six times with washing buffer, and samples were analyzed by Western blot (WB). For detailed antibody information, please see Supplementary Table S2.
Immunofluorescence staining
NP cells were fixed with 4% paraformaldehyde for 15 min and then treated with 0.1% tritonX-100 for 30 min. The cells were blocked with 1% bovine serum albumin for 15 min, followed by incubation with NF-κB antibodies overnight at 4°C. The secondary antibody used was Cy3-conjugated secondary. The nuclei were counterstained with 4',6-Diamidino-2-phenylindole. For detailed antibody information, please see Supplementary Table S2.
Western blot
Cells were lysed on ice with RIPA buffer (PR20001, Proteintech, Rosemont, USA) containing protease inhibitors (Proteintech, Rosemont, USA). After centrifugation, the supernatants were collected, and the samples were separated on a 10% sodium dodecyl sulfate–polyacrylamide gel. Proteins were electro-transferred to a Polyvinylidene fluoride membrane. The membranes were then incubated in skim milk solution (5% w/v) for 2 h at room temperature, and then overnight at 4°C with the primary antibodies. The membranes were washed and incubated with Horseradish peroxidase-conjugated secondary antibody for 40 min at 37°C. For detailed antibody information, please see Supplementary Table S2. Proteins were visualized using an Electrochemiluminescence reagent (Proteintech, Rosemont, USA) and imaged on a chemiluminescence detection system. The protein expression levels were normalized to β-actin.
Enzyme-linked immunosorbent assay
The concentrations of TNF-α (EK182), IL-6 (EK106), IL-10 (EK110), matrix metalloproteinases-3 (MMP-3) (EK1M03), MMP-13 (EK1M13), a disintegrin and metalloproteinase with thrombospondin motifs-4 (ADAMTS-4) (EH1940), ADAMTS-5 (EH2101), and Collagen Ⅱ (EH2865) in the supernatants of human NP cells were examined using enzyme-linked immunosorbent assay kits from MultiSciences Biotech Co. Ltd. (Hangzhou, China) and Wuhan Fine Biotech Co., Ltd. (Wuhan, China). The supernatants were assayed according to the manufacturer’s instructions.
Bioinformatics analysis
Gene expression profiling datasets were obtained from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/; accession numbers: GSE186542 and GSE147383). The screening criteria for differentially expressed genes (DEGs) are |log2FoldChange| > 2 and P value <0.005. A Venn diagram was used to show shared DEGs of the two datasets. The heat maps were utilized to illustrate the differential expression of the shared genes between patients with IVDD and control samples.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 9.5. The t-test was used to compare differences between two groups, and one-way analysis of variance was used to compare differences among multiple groups. The P values were indicated in the figures. P values <0.05 were considered significant.
Results
GPNMB expression and correlation between GPNMB and IL-6 or COL2A1 in clinical NP samples
To initially explore the role of GPNMB in IVDD, we collected degenerative NP samples from 40 patients with IVDD and determined the expression of GPNMB, IL-6, and COL2A1. Representative MRI results of three patients with mild IVDD and three patients with severe IVDD were shown in Figure 1A, and the degree of degeneration of the NP specimens was further confirmed using HE staining (Fig. 1B). The real-time quantitative PCR results showed that GPNMB mRNA expression was significantly suppressed in patients with severe IVDD compared with mild IVDD (Fig. 2A). Inflammation is one of the prominent pathological features of IVDD (Risbud and Shapiro, 2014). Compared with patients with mild IVDD, the mRNA expression of IL-6, a pro-inflammatory factor, was significantly up-regulated in patients with severe IVDD (Fig. 2B). Moreover, the mRNA expression of COL2A1, the important component of NP ECM, was significantly reduced in NP tissues from patients with severe IVDD (Fig. 2C). Correlation studies showed that, in patients with mild and severe IVDD, decreased GPNMB levels correlated with increased IL-6 expression and decreased COL2A1 levels (Fig. 2D and E). These results suggested that GPNMB may play an important role in IVDD.
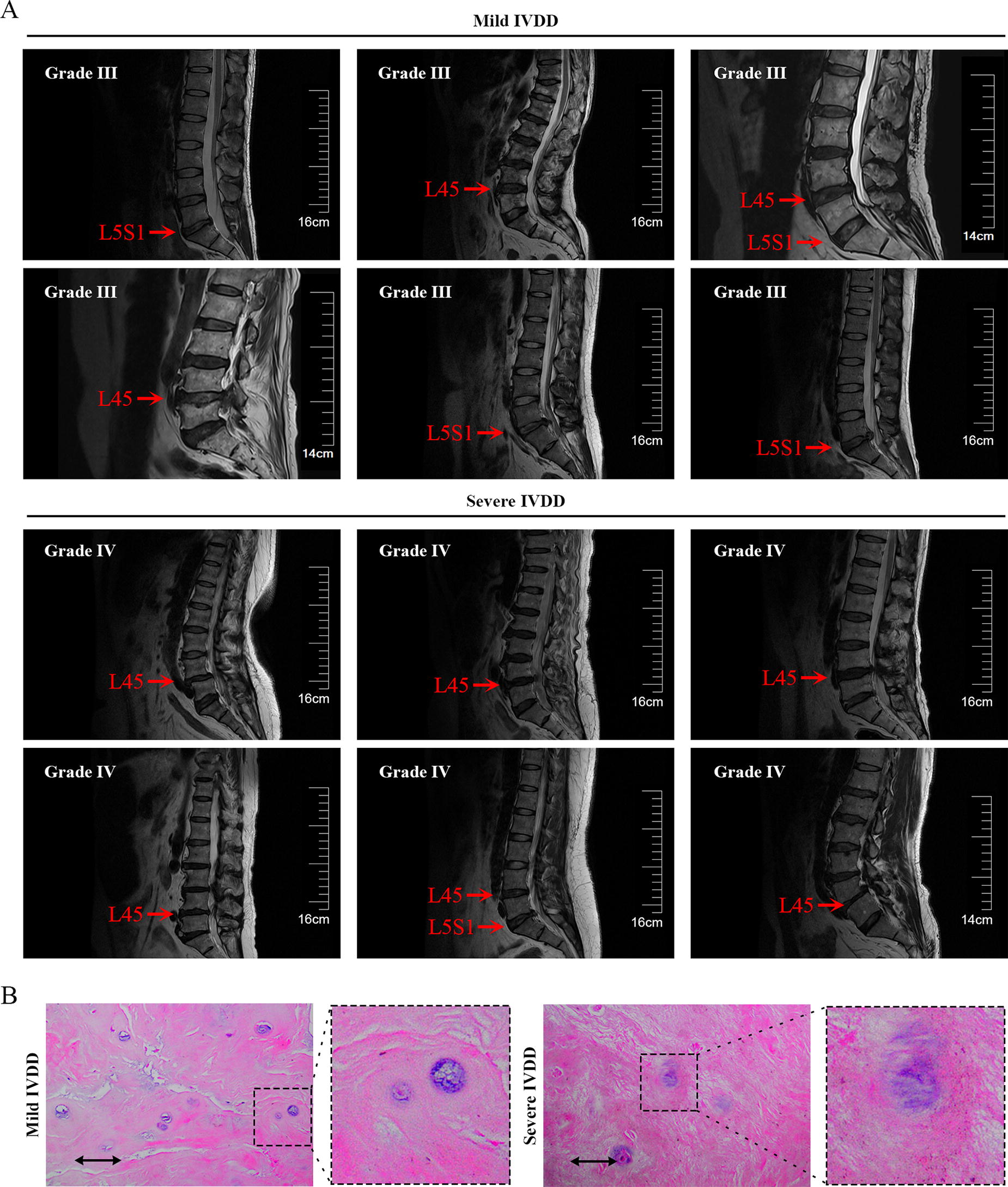
Cases of patients with intervertebral disc degeneration (IVDD) in mild or severe conditions.
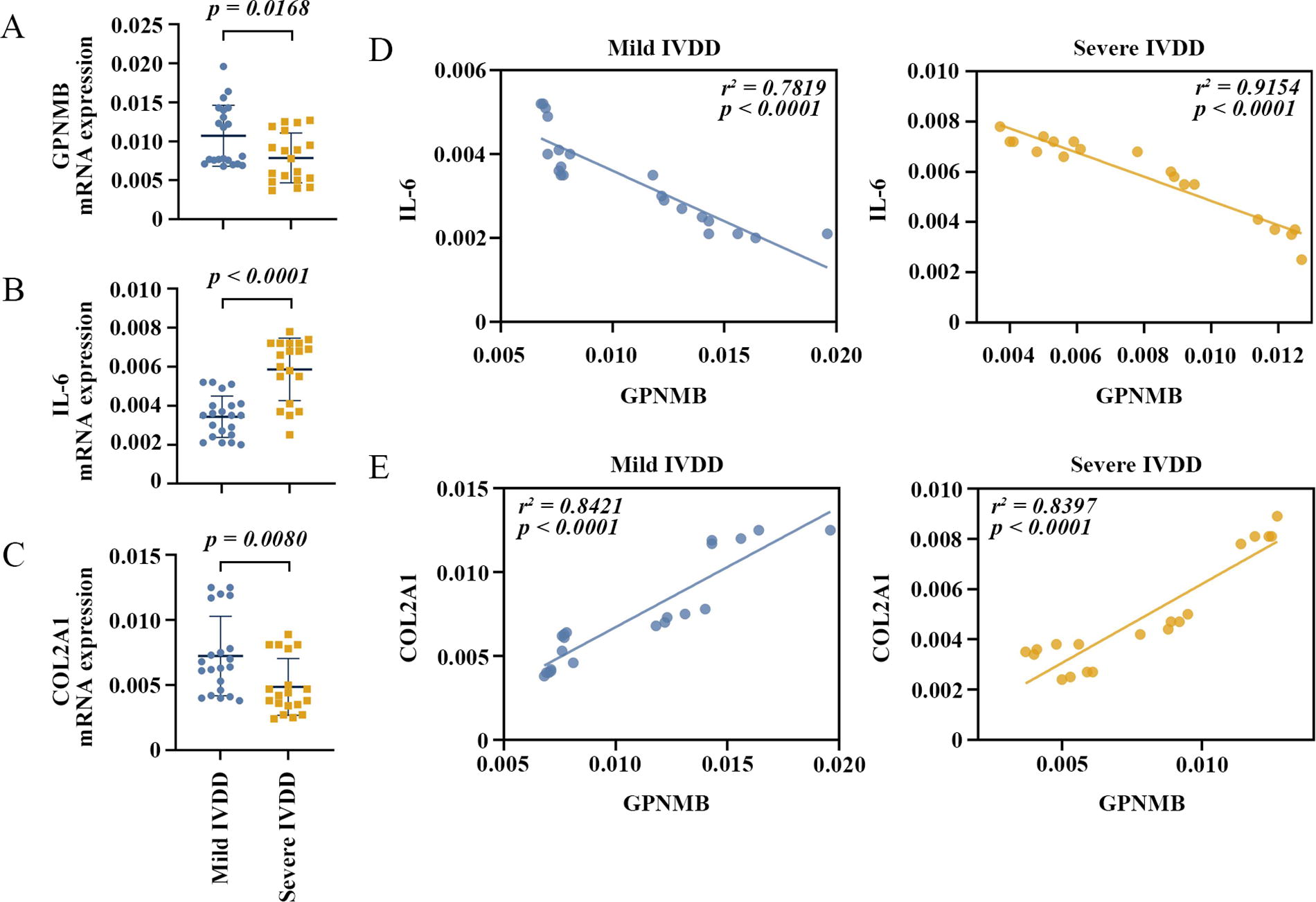
GPNMB expression and correlation between GPNMB and IL-6 or COL2A1 in clinical NP samples. GPNMB
GPNMB overexpression reduces the inflammatory response in IL-1β-induced NP cells
We overexpressed GPNMB in human primary NP cells and stimulated cells with IL-1β to explore the effects of GPNMB on NP cells (Fig. 3A). Efficiency of overexpression of GPNMB expression levels was verified (Fig. 3B). The protein and mRNA expression of GPNMB were decreased in NP cells exposed to IL-1β, while we showed that GPNMB was increased in IL-1β-induced NP cells after GPNMB overexpression (Fig. 3C). As shown in Figure 3D, IL-1β induction significantly decreased the OD value of NP cells, while GPNMB overexpression remarkedly promoted NP cell survival. In GPNMB overexpressed NP cells, the mRNA expression of pro-inflammatory cytokines TNF-α and IL-6 (Fig. 3E) and the content of secreted TNF-α and IL-6 (Fig. 3F) were significantly down-regulated. GPNMB overexpression notably increased anti-inflammatory cytokine IL-10 mRNA expression (Fig. 3E) and the concentration of secreted IL-10 (Fig. 3F) in NP cells under IL-1β stimulation. These results indicated that GPNMB overexpression may inhibit the progression of IVDD by suppressing the inflammatory response in NP cells.

GPNMB overexpression reduces the inflammatory response in IL-1β-induced NP cells.
GPNMB overexpression inhibits ECM degradation in IL-1β-induced NP cells
The effect of GPNMB on ECM degradation was explored in vitro by assaying collagen type Ⅱ and several key ECM-degrading enzymes including MMPs and ADAMTS. As presented in Figure 4A and B, the overexpression of GPNMB significantly increased the COL2A1 mRNA expression and secreted COL2A1 in NP cells incubated with IL-1β. The overexpression of GPNMB notably inhibited the IL-1β-induced increase in relative mRNA expression of MMP-3 and MMP-13 in NP cells, while the MMP-3 and MMP-13 concentrations were markedly decreased in the cell supernatant. In addition, overexpressed GPNMB significantly decreased ADAMTS-4 and ADAMTS-5 mRNA expression and production in NP cells treated with IL-1β. These results implied that GPNMB overexpression may inhibit IVDD by inhibiting ECM degradation.
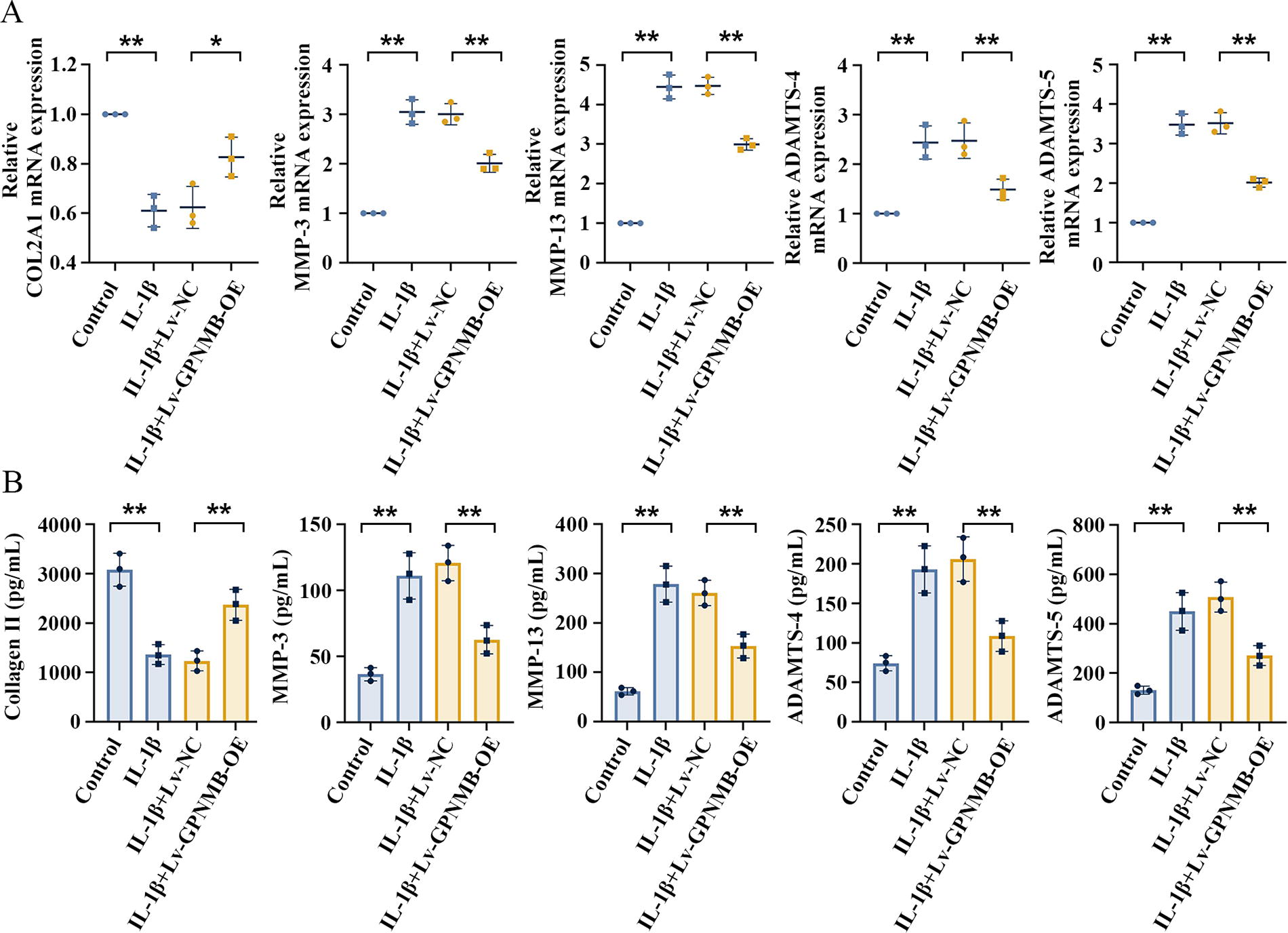
GPNMB overexpression inhibits extracellular matrix (ECM) degradation in IL-1β-induced NP cells.
GPNMB affects the NF-κB pathway by binding to CD44 in NP cells
GPNMB may bind to and signal through CD44 (Neal et al., 2018; Sondag et al., 2016). The co-immunoprecipitation results showed that GPNMB binds to CD44 in NP cells (Fig. 5A). The NF-κB signaling pathway is a downstream pathway of CD44. GPNMB overexpression decreased the NF-κB p-p65 relative protein expression in IL-1β-induced NP cells (Fig. 5B). As demonstrated in the immunofluorescence results (Fig. 5C), IL-1β induced NF-κB p65 translocation into the nucleus, and GPNMB overexpression suppressed this trend. These results indicated that GPNMB overexpression may inhibit the NF-κB pathway by binding to CD44.
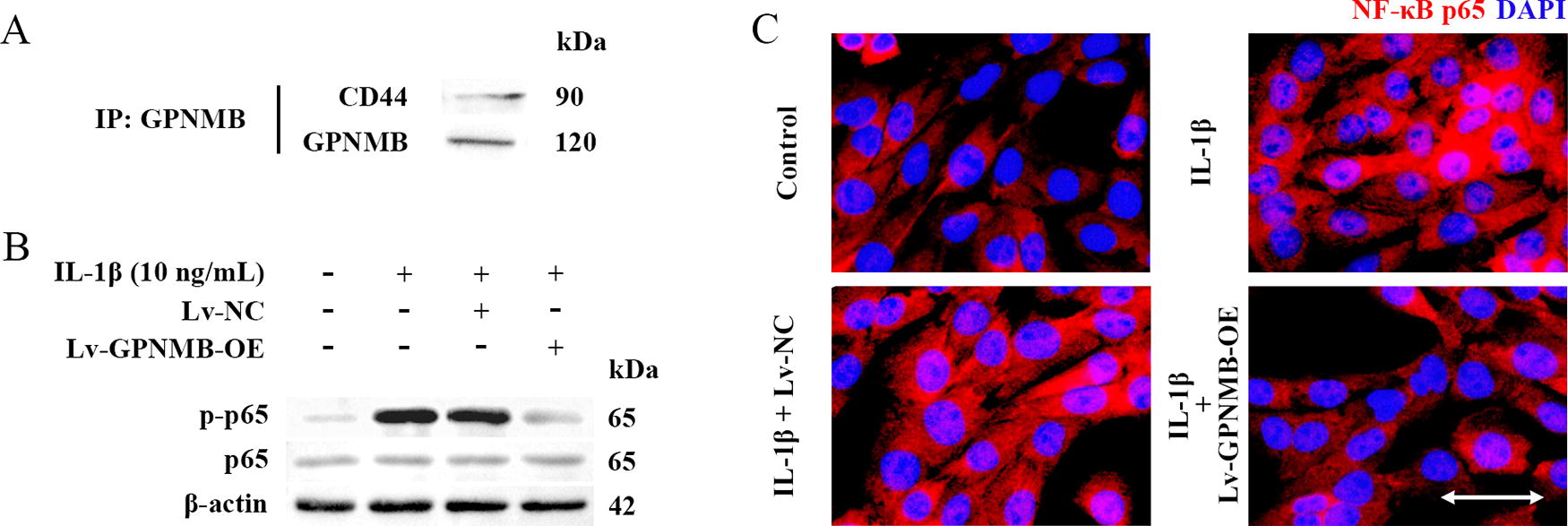
GPNMB affects the NF-κB pathway by binding to CD44 in NP cells.
Discussion
GPNMB was initially described as more highly expressed in low-metastatic melanoma cell lines (Weterman et al., 1995). Recent studies have identified the roles of GPNMB in neurodegenerative diseases, renal diseases, intestinal diseases, and cancers through its involvement in neuroinflammatory or inflammatory responses (Saade et al., 2021). GPNMB was investigated in IVDD for the following reasons. A bioinformatics analysis was performed using the GEO database, and we found that GPNMB was one of the shared DEGs in the datasets GSE186542 and GSE147383 (Supplementary Fig. S1A). The expression analysis of the shared DEGs showed that GPNMB gene expression was significantly down-regulated in the NP tissues of patients with IVDD, along with C3 and PDE1A (Supplementary Fig. S1B and C). The effects of C3 in IVDD have been explored (Kuhn et al., 2023). PDE1A has not been reported in IVDD, and its expression change was involved in the inflammatory response after traumatic brain injury (Oliva et al., 2012). However, more in-depth studies of PDE1A in inflammation are lacking. Therefore, GPNMB became our most interesting gene to be investigated in IVDD.
In this study, we investigated the expression of GPNMB in NP tissues from herniated discs of patients, analyzed its relationship with key pathophysiological factors causing disc degeneration, and explored its possible mechanism of action in NP cells. Specifically, GPNMB mRNA expression was inhibited in the NP tissues of patients with severe IVDD compared with patients with mild IVDD, which was consistent with the GPNMB expression trend using bioinformatics analysis. Markedly, GPNMB expression was negatively correlated with IL-6 expression and positively correlated with Collagen II in clinical IVDD NP samples. In addition, elevated pro-inflammatory cytokines secreted by NP cells will contribute to ECM degradation (Risbud and Shapiro, 2014). This process resulted in an imbalance between catabolic and anabolic responses of ECM (Liang et al., 2023; Risbud and Shapiro, 2014). The above evidence implied that GPNMB may exert a protective effect in NP tissues by inhibiting pro-inflammatory cytokine secretion and ECM degradation.
Previous studies have demonstrated that GPNMB attenuates the inflammatory response in a variety of inflammation-related diseases (Prabata et al., 2021; Saade et al., 2021; Song and Lin, 2019). We provided some evidence that GPNMB overexpression exerted an anti-inflammatory effect in IL-1β-induced human primary NP cells. GPNMB overexpression inhibited the expression of pro-inflammatory cytokines TNF-α and IL-6 while promoting the anti-inflammatory cytokine IL-10 expression in human NP cells. This finding is in harmony with the role GPNMB played in the inflammatory response in periodontal disease (Song and Lin, 2019). IL-10 as an anti-inflammatory cytokine tended to be up-regulated in IL-1β-stimulated NP cells in the study, which is consistent with the findings of Wang et al. (2018). The reasons for its up-regulation remain unclear, and possible explanations maybe its high responsiveness to pro-inflammatory stimuli and the elevated IL-10 showed a protective mechanism in response to IVDD (Wang et al., 2018).
Furthermore, GPNMB overexpression exerted an inhibitory effect on ECM degradation in human cells. Collagen II secreted by NP cells is one of the major components of the ECM in NP tissues (Amin et al., 2017). Overexpression of GPNMB significantly increased Collagen II secretion, and decreased matrix metalloproteinase MMP-3 and MMP-13, which are proteases involved in the degradation of Collagen II. The decreasing matrix metalloproteinase may account for the increased expression of Collagen II. ADAMTS-4/5 is the major enzyme for degrading aggrecan, which is another major component of the ECM (Amin et al., 2017), and ADAMTS-4/5 expression was also inhibited in our study. IL-1β treatment inhibited the survival of NP cells (Lin et al., 2019). This result is comparable with our research, and we found that GPNMB overexpression was able to promote IL-1β-induced NP cell survival. These findings suggested that GPNMB overexpression protected NP cells by inhibiting inflammatory responses and ECM degradation.
It has been reported that GPNMB is post-transcriptionally located on the cell membrane, where it can be cleaved by a metalloproteinase, resulting in the release of a soluble fragment (Rose et al., 2010). This fragment binds to many receptors, including CD44, Na+, K+-ATPase, epidermal growth factor receptor, and vascular endothelial growth factor receptor, and elicits cellular responses (Saade et al., 2021). GPNMB was found to reduce astrocyte-mediated neuroinflammation in a CD44-dependent manner (Neal et al., 2018). GPNMB inhibited NF-κB signaling by binding to CD44, which reduces the inflammatory capacity of macrophages (Prabata et al., 2021). We found that in NP cells, GPNMB bound to CD44, which is consistent with the findings of Yu et al. in osteoclasts (Sondag et al., 2016) and Sondag et al. in mesenchymal stem cells (Yu et al., 2016). Further, in our study, GPNMB overexpression inhibited NF-κB signaling in IL-1β-incubated NP cells, which may be achieved through the binding of GPNMB to CD44.
Conclusion
In conclusion, this study confirms that GPNMB overexpression inhibits the inflammatory response and ECM degradation in NP cells. This effect may be due to GPNMB binding to the receptor CD44, inhibiting NF-κB signaling in NP cells.
Footnotes
Authors’ Contributions
J.Z., and B.Y.: Conceptualization. J.Z., and Y.S.: Investigation, formal analysis. J.Z.: Writing—original draft, validation. Y.S.: Visualization. B.Y.: Writing—review and editing, project administration.
Ethics Approval
The study was conducted in accordance with the Declaration of Helsinki. Informed consent was obtained from each patient involved in the study. The ethics committee of Anhui No.2 Provincial People’s Hospital approved this study protocol.
Data Availability
All data generated or analyzed during this study are included in this published article.
Author Disclosure Statement
The authors have no competing interests to declare that are relevant to the content of this article.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2