
Abstract
Listeriosis is a foodborne disease caused by Listeria monocytogenes (Lm) that usually leads to serious adverse outcomes in pregnant women. Interleukin (IL)-33/serum stimulation (ST)2 axis has an important impact on infectious diseases, but its role in listeriosis is rarely studied. Here, the immunomodulatory effects of IL-33/ST2 axis during perinatal Lm infection were investigated. In our study, the perinatal Lm infection model was constructed by injecting Lm into the tail vein of C57BL/6J mice. IL-33/ST2 axis was blocked by intraperitoneal injection of anti-IL-33Rα/ST2 antibody. In vitro, mouse cytotoxic T lymphocyte cell line (CTLL)-2 cells were activated by using CD3/CD28 antibodies. Perinatal Lm infection caused massive necrosis of liver tissue. Blocking IL-33/ST2 axis in pregnant mice inhibited the infiltration of CD8+ T lymphocytes into the site of infection and further aggravated liver damage. We also found that IL-33 promotes mitochondrial autophagy in activated CTLL-2 cells in vitro. Mitochondrial autophagy was beneficial to the clearance of damaged mitochondria and reduced the production of reactive oxygen species. IL-33/ST2 axis affects the immune function of CD8+ T lymphocytes by regulating mitophagy, which plays a very important role in the occurrence and development of perinatal Lm infection. Immunomodulation targeting IL-33/ST2 axis may be an effective way to adjuvant treatment of perinatal Lm infection.

Introduction
Listeria monocytogenes (Lm) is a zoonotic gram-positive bacteria that can grow and reproduce at low temperatures and is commonly found in a variety of foods, including vegetables, fruits, meat, seafood, and raw dairy products. Pregnant women, newborns, the elderly, and immunocompromised people are susceptible to Lm and may suffer from gastroenteritis, meningoencephalitis, stillbirth, and spontaneous abortion, with mortality rates as high as 20% (Kayode and Okoh, 2022). Lm is an intracellular parasite that inhibits light chain 3 (LC3)-associated phagocytosis by regulating mitochondrial Ca2+ signaling to promote its intracellular survival (Li et al., 2021). Previous studies have shown that the number of infiltrated CD3+T cells and CD8+T cells in the intestinal lamina propria increased significantly in mice infected with Lm orally (Drolia et al., 2020), suggesting that T cells are a key factor in the process of fighting Lm infection. Host clearance of Lm mainly depends on cellular immune function (Ma et al., 2024). The most recent serious outbreak of Lm infection occurred in South Africa in 2017, caused by pork sausages contaminated with Lm. In this outbreak, a total of 937 cases of Lm infection were confirmed, of which 193 died (Thomas et al., 2020). The risk of Lm infection in pregnant women is at least 12–20 times that of the general population (Elinav et al., 2015). Globally, pregnancy combined with listeriosis accounts for 20.7%–43% of total cases (de Noordhout et al., 2014). A study investigated the incidence of listeriosis in pregnancy in Beijing Obstetrics and Gynecology Hospital, China, between January 1, 2013, and December 31, 2018, and found 12 cases of listeriosis among 87,644 pregnant women who delivered, with an incidence rate of 13.7 cases per 100,000 (Li et al., 2020). The outbreak of listeriosis can cause serious health problems and economic losses, which needs to be paid more attention to.
IL-33 is a multifunctional cytokine, acting either as a typical extracellular cytokine or as a nuclear transcription factor (Aggeletopoulou et al., 2022). In humans, IL-33 is constitutionally expressed in vascular endothelial cells, epithelial cells and stromal cells in various organs such as the liver and lung (Cayrol, 2021). In the homeostatic state, full-length IL-33 is expressed in the nucleus, but cell damage and necrosis caused by external stimulation enhances the expression of IL-33, leading to its release outside the cell. Inflammatory proteases synthesized by neutrophils and mast cells lyse full-length IL-33 into a shorter mature form, thereby fulfilling its biological function (Guo et al., 2022; Rasheed et al., 2022). IL-33 is closely associated with a variety of infectious diseases, such as Lm infection (Li et al., 2022), severe acute respiratory syndrome coronavirus 2(SARS-CoV-2) infection (Tsilioni and Theoharides, 2023), human bocavirus infection (Alkubaisi et al., 2023), enterotoxigenic Escherichia coli infection (Motyka et al., 2022), and tuberculosis (de Sá et al., 2022), where IL-33 is involved in the regulation of immune response. Via binding to the receptor complex composed of membrane-bound receptor (ST2L) and IL-1 receptor accessory protein (IL-1RAcP), IL-33 signals activate the nuclear factor kappa-B, mitogen-activated protein kinase, and phosphoinositide 3-kinase /protein kinase B pathways, which leads to an increase in cytokines such as tumor necrosis factor-α (TNF-α), IL-1, and IL-6 and induces appropriate immune responses (Brunetti et al., 2023; Fairlie-Clarke et al., 2018; Glineur et al., 2022). IL-33/ST2 signaling pathway plays a dual role in a series of infectious diseases, which not only promotes pathological immune response but also facilitates tissue repair.
Here, we studied the immune regulation effect of IL-33/ST2 axis on CD8+ T cells, and established a perinatal in vivo infection animal model by injecting Lm into the tail vein, exploring a new strategy for the treatment of perinatal Lm infection from a new perspective, and supplementing and improving the current antibiotic treatment regimen.
Materials and Methods
Materials
Protease and Phosphatase Inhibitor Cocktail for Mammalian (Applygen, P1261); Horseradish Enzyme-labeled Goat Anti-Rabbit IgG (ZSGB-BIO, ZB-5301); Horseradish enzyme-labeled Goat anti-mouse IgG (ZSGB-BIO, ZB-5305); Alexa Fluor® 488 Labeled Goat anti-mouse IgG (ZSGB-BIO, ZF-0512); Alexa Fluor 594 Labeled Goat Anti-Rabbit IgG (ZSGB-BIO, ZF-0516); rabbit anti-UNC-51-like kinase 1 (ULK1) antibody (Thermo Fisher Scientific, MA5-32699); rabbit anti-Beclin 1 antibody (abcam, ab207612); rabbit anti-p62 antibody (Thermo Fisher Scientific, MA5-42726); mouse anti-β-actin antibody (Thermo Fisher Scientific, MA5-15739); rabbit anti- LC3B antibody (Cell Signaling Technology, 43566T); Antifade Mounting Medium (Beyotime, P0126); mouse anti-TOMM20 antibody (Santa Cruz Biotechnology, sc-17764); rabbit anti-LAMP1 antibody (abcam, ab208943) rat anti-mouse IL-33Rα (ST2) antibody (Biolegend, 146604); Recombiant anti-CD8 alpha Rabbit mAb (Servicebio, GB15068).
Animals
All experimental procedures were approved by the Welfare Ethics Committee of Beijing Laboratory Animal Research Center (NO.BLARC-LAWER-202408001). We purchased 8-week-old female C57BL/6J mice and 7-week-old male C57BL/6J mice from HFK BIOSCIENCE CO., LTD (Beijing, China). C57BL/6J mice were fed standard laboratory chow and water in a standard temperature-controlled room with a temperature of 22 ± 1°C. Female mice were caged with male mice in a 2:1 ratio. After mating, the start of pregnancy is determined by examining the vaginal plugs. Successful mating was confirmed by checking for the presence of a vaginal plug the next morning, which was considered to be day 0.5 of gestation.
Lm (China General Microbiological Culture Collection Center, ATCC19115) stored at −80°C were inoculated onto solid plates of brain and heart infusion broth (BHI), incubated at 37°C for 20 h, and a single colony was inoculated into BHI liquid medium. After oscillating culture at 37°C for 16–18 h, the colonies were collected by centrifugation. Twenty pregnant mice at 14.5 days gestation were randomly divided into 4 groups: control, Lm, anti-ST2, and Lm + anti-ST2. Pregnant mice in the infected group were injected with 2.5 × 105 CFU of Lm through a tail vein to establish the perinatal Lm infection model. Pregnant mice in anti-ST2 group and Lm + anti-ST2 group were intraperitoneally injected with 100 μg anti-IL-33Rα(ST2) antibody every other day. The mice were sacrificed at day 17.5 of gestation, and serum, spleen, liver, and lungs were collected. The serum was used to detect the content of cytokines, the spleen was used to isolate and extract CD8+T cells, and the liver and lung were embedded in paraffin for pathological and immunofluorescence analysis.
Cell culture and activation
Mouse CTLL-2 cells (Meisen CTCC, China, CTCC-001-0171) were cultured in RPMI-1640 medium (C11875500BT, Gibco) supplemented with 10% fetal bovine serum (FBS), 1% penicillin-streptomycin solution, and 100 U/mL IL-2 (novoprotein, China, CK24) at 37°C in a humidified air containing 5% CO2. The cells were activated by incubating with 2μg/mL CD3(Biogems, 05112-25-500) and 2μg/mL CD28 (Biogems, 10312-25-500) for 2 days.
Cell viability assay
CTLL-2 cells at different concentrations were inoculated into 96-well plates, and cell growth curves were obtained by using cell counting kit-8 (CCK-8) (Glpbio, GK10001). In total, 3 × 104 cells with 100 μL culture medium per well in 96-well plates were treated with IL-33 (500 ng/mL) (abmole, M31329, E. coli, 95%), and/or treated by removing IL-2 in culture medium. The cell viability was detected at 12, 24, 36, and 48 h after treatment. At these time points, 10μL CCK-8 solution was incorporated into 96-well plates and incubated at 37°C for 2 h. The absorbance at 450 nm was then measured by GloMax Discover (Promega, New York, USA).
Transmission electron microscopy
CTLL-2 cells were fixed at 4°C with 2.5% glutaraldehyde and stored at 4°C. The next day, the cells were immobilized at room temperature with osmic acid for 2 h and gradiantly dehydrated with alcohol. The cell sample is then permeated and coated with epoxy resin. The sample was cut into slices of 100 μm thickness and stained with uranium acetate and lead citrate. Finally, whether autophagosome or autophagolysosome appeared in the cells was observed under an electron microscope.
Immunofluorescence staining
The cells were fixed with Immunol Staining Fix Solution (Beyotime, P0098) for 15 min, followed by blocking with 3% Albumin Bovine (BSA) (BioFroxx, 4240GR100) at room temperature for 1 h, incubation with primary antibody at 4°C overnight, and incubation with secondary antibodies at room temperature for 1 h. The nucleus was stained with DAPI (Solarbio, C0065) for 10 min. A laser confocal microscope (Olympus, Japan) was used to capture immunofluorescence.
The liver and lung sections were treated with dewaxing hydration, antigen repair, and endogenous peroxidase blocking, and then blocked with 3% BSA at room temperature for 1 h. The sections were placed in a humidified chamber and incubated with primary antibody at 4°C overnight, and then incubated with secondary antibody at room temperature for 1 h. The sections were stained with 4′,6-Diamidino-2-phenylindole (DAPI) for 10 min at room temperature in the dark. A scanner (3DHISTECH CaseViewer, Hungary) was used to detect immunofluorescence.
Autophagy staining assay with monodansylcadaverine
Monodansylcadaverine (MDC) is a fluorescent probe that binds specifically to membrane lipids to label autophagosomes. MDC (1000×) (C3018S, Beyotime) was diluted to MDC (1×) by Assay Buffer at a ratio of 1:1000 to obtain MDC stain solution. In total, 1 million cells were added to 1 mL MDC staining solution and incubated in a cell incubator at 37°C for 30 min without light. Next, cells were centrifuged at 250g for 5 min at room temperature to remove MDC staining solution and washed 3 times with assay buffer. Finally, fluorescence detection was carried out by GloMax Discover (Promega, New York, USA).
Assay of superoxide and ATP
The Superoxide Assay Kit (Beyotime, S0060) was used to measure the content of superoxide. Catalase solution, WST-1 solution, and detection buffer are mixed together in a ratio of 1:5:100 to form the detection working solution. The cells were collected by centrifugation, resuspended with the working solution, and incubated at 37°C for 3, 33, 63, 93, and 123 min. The cell suspension was then transferred to a 96-well plate with approximately 20,000 cells per well, and the absorbance at 450 nm was immediately measured by GloMax Discover (Promega, New York, USA).
ATP Assay Kit (Beyotime, S0026) was used to detect ATP content. 200 μL lysis buffer was added to every 6 × 105 cells to fully release intracellular ATP. After lysis, the supernatant was collected. 100 μL ATP test solution and 10 μL sample were added to each well of an opaque 96-well plate and mixed. After at least 2 s, RLU values were measured with GloMax Discover (Promega, New York, USA). In order to eliminate the error caused by the difference of protein content in the sample, the protein concentration was measured, and the obtained ATP concentration was calibrated.
Reactive oxygen species staining
The Reactive Oxygen Species Assay Kit (Yeasen Biotechnology, 50101ES01) was used to measure intracellular reactive oxygen species (ROS) levels. The intensity of green fluorescence is proportional to the level of ROS. Before loading the probe, 2,7-dichlorodi-hydrofluorescein diacetate (DCFH-DA) was diluted to 10 μM in serum-free medium. The cells were mixed with DCFH-DA working solution and incubated at 37°C for 30 min without light. The cells were washed twice with serum-free medium to remove the DCFH-DA that did not enter the cells. Finally, smears were prepared and observed under laser confocal microscopy (Leica, Germany).
Isolation and extraction of primary CD8+T cells
Spleen was disrupted in phosphate buffered saline (PBS) containing 2% FBS. The cell suspension was passed through a 70 μm mesh nylon strainer to remove aggregates and debris. After centrifugation, the cells were resuspended in PBS containing 2% FBS and 1 mM EDTA to a density of 1 × 108 cells/mL. EasySepTM Mouse CD8+T Cell Isolation Kit (STEMCELL Technologies, 19853) was used to isolate and extract CD8+T cells from spleen. Primary CD8+T cells were divided into 2 parts; one part was used for flow cytometry, and the other part was cultured in complete culture medium for 24 h, and the cell supernatant was collected to measure the content of cytokines.
Detection of the cytokines
The levels of cytokines (IFN-γ, TNF-α, IL-4, IL-10) in serum, primary CD8+T cell supernatant, and CTLL-2 cell supernatant were detected by ELISA (enzyme-linked immunosorbent assay) Kit (BOSTER, EK0375, EK0527, EK0405, EK0417). The 100 μL sample and 100 μL standard product were added to the enzyme-labeled plate hole coated with antibody and incubated at 37°C for 90 min. Next, Biotin-labeled antibodies were added and incubated at 37°C for 60 min and the board was washed 3 times with washing buffer. Then, Avidin-peroxidase complex (100X) (ABC) was added and incubated at 37°C for 30 min, and the board was washed 5 times with washing buffer. TMB Color Developing Reagent was continued to be added and incubated at 37°C for 15 min. Finally, the termination solution was added, and the optical density (OD) value was measured at 450 nm using GloMax Discover (Promega, New York, USA).
Flow cytometry
Primary CD8+T cell was collected and resuspended with D-PBS (1 × 107 cells/mL). First, Zombie Yellow™ Fixable Viability Dye (Biolegend, 423103) was added to the cell suspension, incubated at room temperature away from light for 20 min, and then centrifuged to remove the dye. In order to label T cell surface molecules (CD25, CD30, CD69, CD103, CD126), the following flow antibodies were mixed together into the staining system and incubated at room temperature in the dark for 20 min: PE anti-mouse CD25 Antibody (0.25 µg per million cells) (Biolegend, 113703), BV421 Hamster Anti-Mouse CD30 (0.25 µg per million cells) (BD OptiBuild™, 740031), PE/Dazzle™ 594 anti-mouse CD69 Antibody (0.25 µg per million cells) (Biolegend, 104535), PE/Cyanine5 anti-mouse CD103 Antibody (1.0 µg per million cells) (Biolegend, 121447), APC anti-mouse CD126 (IL-6Rα chain) Antibody (1.0 µg per million cells) (Biolegend, 115811). The cells were then fixed with fixation buffer (BD Cytofix™, 554655) at 4°C for 20 min in the dark. Finally, the cells were transferred to staining buffer, and mean fluorescence intensity of cell surface molecules was analyzed by flow cytometry.
Western blotting analysis
T cells were lysed with RIPA cell lysate (Solarbio, R0010), and the Enhanced BCA Protein Assay Kit (Beyotime, P0010) was used to determine total protein concentration. An equal amount of protein sample was separated on sodium dodecyl sulfate-polyacrylamide gel and then transferred to a Polyvinylidene Fluoride (PVDF) membrane. The membrane was incubated with 5% skim milk for 1 h, and then incubated with primary antibody at 4°C overnight. On the second day, the membrane was washed with Tris-buffer saline/Tween 20 (TBST) 3 times, incubated with secondary antibody at room temperature for 1 h, washed with TBST 3 times, and then reacted with chemiluminescent horseradish peroxidase substrate. The developed bands were detected by the ChemiDoc MP Imaging System from Bio-Rad (California, USA). The gray scale of the bands was analyzed by Image J.
Histopathological analysis
The mouse liver and lung tissues that had been embedded into paraffin blocks were cut into 4um sections. The nucleus was stained with hematoxylin and the cytoplasm with eosin. After the slices were dehydrated with 95% alcohol, anhydrous ethanol, and xylene until transparent, neutral gum was used to seal the slices. Finally, the tissue structure was visualized using a scanner (3DHISTECH CaseViewer, Hungary) to perform histological analysis of liver and lung tissue.
Statistical analysis
All data were presented as the mean ± standard error of the mean (SEM). Statistical analysis was performed using GraphPad Prism software (version 9.5.1). Statistically significant differences were determined by Student’s unpaired t-test or Kolmogorov-Smirnov test. P < 0.05 was considered statistically significant.
Results
Blocking IL-33/ST2 axis inhibited the infiltration of CD8+T cells into the liver and aggravated liver injury in perinatal Lm-infected mice
The middle lobe of the liver and the right lower lung of each group of mice were made into tissue sections for pathological observation. According to the nonalcoholic fatty liver disease (NAFLD) scoring standard, the pathological score of liver tissue was performed according to the steatosis and balloon-like degeneration of hepatocytes and the number of necrotic lesions in hepatic lobules. The results showed that the liver of Lm-infected mice was damaged, and the liver damage was further aggravated after treatment with anti-IL-33Rα/ST2. The lung tissue structure of Lm-infected mice was basically normal (Fig. 1A–C). The left outer lobe of the liver and left lung of mice in each group were made into tissue sections, and CD8+T cells were labeled by immunofluorescence. Compared with the control group, the number of CD8+T cells in the liver of Lm-infected mice was significantly increased, and this situation was reversed after intraperitoneal injection of anti-IL-33Rα/ST2. Although Lm infection did not involve the lung, the number of CD8+T cells in the lung tended to increase, and this trend was suppressed after anti-IL-33Rα/ST2 treatment (Fig. 1D–F).

Blocking IL-33/ST2 axis inhibited the infiltration of CD8+T cells into the liver and aggravated liver injury in perinatal Lm-infected mice. The pregnant C57BL/6J mice were infected with Lm (2.5 × 105 CFU/mouse). IL-33/ST2 axis was blocked by intraperitoneal injection of anti-IL-33Rα/ST2 (100 μg/mouse). Then mice were sacrificed 3 days after treatment.
Anti-IL-33Rα/ST2 inhibited the secretion of pro-inflammatory cytokines by primary CD8+T lymphocytes and affected the expression of CD25, CD30, CD69, CD103, and CD126 on their membrane surface
The cytokine content in serum and supernatant of primary CD8+T cells was measured in mice infected with Lm, with or without intraperitoneal injection of anti-IL-33Rα/ST2. Interferon-γ (IFN-γ) and TNF-α are pro-inflammatory cytokines, and IL-4 and IL-10 are anti-inflammatory cytokines. The levels of IFN-γ, TNF-α, and IL-4 in the serum of Lm-infected mice were increased. Blocking IL-33/ST2 axis in mice further increased IFN-γ and TNF-α, while the level of IL-4 decreased, suggesting that blocking this axis promoted systemic inflammatory response. Primary CD8+ T lymphocytes were extracted from the spleens of mice in each group and cultured for 24 h in vitro. The culture medium supernatant was collected to detect cytokine levels. We found that compared with the control group, the primary CD8+ T cells extracted from the spleens of mice in the Lm infection group secreted more IFN-γ, while the secretion of IL-4 and IL-10 decreased. Blocking IL-33/ST2 axis reduced the secretion of these 3 cytokines by primary CD8+ T cells, suggesting that blocking this axis inhibited the immune function of primary CD8+ T cells. (Fig. 2A–F). In addition, we found that the expressions of CD25, CD30, and CD69 on the surface of primary CD8+T cells in the Lm infection group were enhanced, while the expressions of CD103 and CD126 were decreased. After anti-IL-33Rα/ST2 treatment, the expression of CD25, CD30, and CD103 was reversed, the expression of CD69 was further enhanced, and the expression of CD126 was further decreased (Fig. 2G–P).

Anti-IL-33Rα/ST2 inhibited the secretion of pro-inflammatory cytokines by primary CD8+T lymphocytes and affected the expression of functional molecules on their membrane surface.
IL-33 promotes mitochondrial autophagy flow in CD3/CD28 antibody-activated CTLL-2 cells
The growth rate of CTLL-2 cells increased with the increase of density within a certain range (Fig. 3A). The survival of CTLL-2 cells was dependent on IL-2. If IL-2 was removed from the culture medium, almost all CTLL-2 cells were apoptotic after 24 h. Obviously, IL-33 had no significant effect on the proliferative activity of resting CTLL-2 cells (Fig. 3B). We analyzed the key regulatory molecules of mitochondrial autophagy by Western blotting. In CTLL-2 cells activated by CD3/CD28 antibody, IL-33 treatment increased the expression of ULK1 and Beclin1, as well as the LC3II/LC3I ratio, while decreasing the expression of P62 (Fig. 3C–G). These results suggest that IL-33 promotes mitochondrial autophagy in activated CTLL-2 cells.
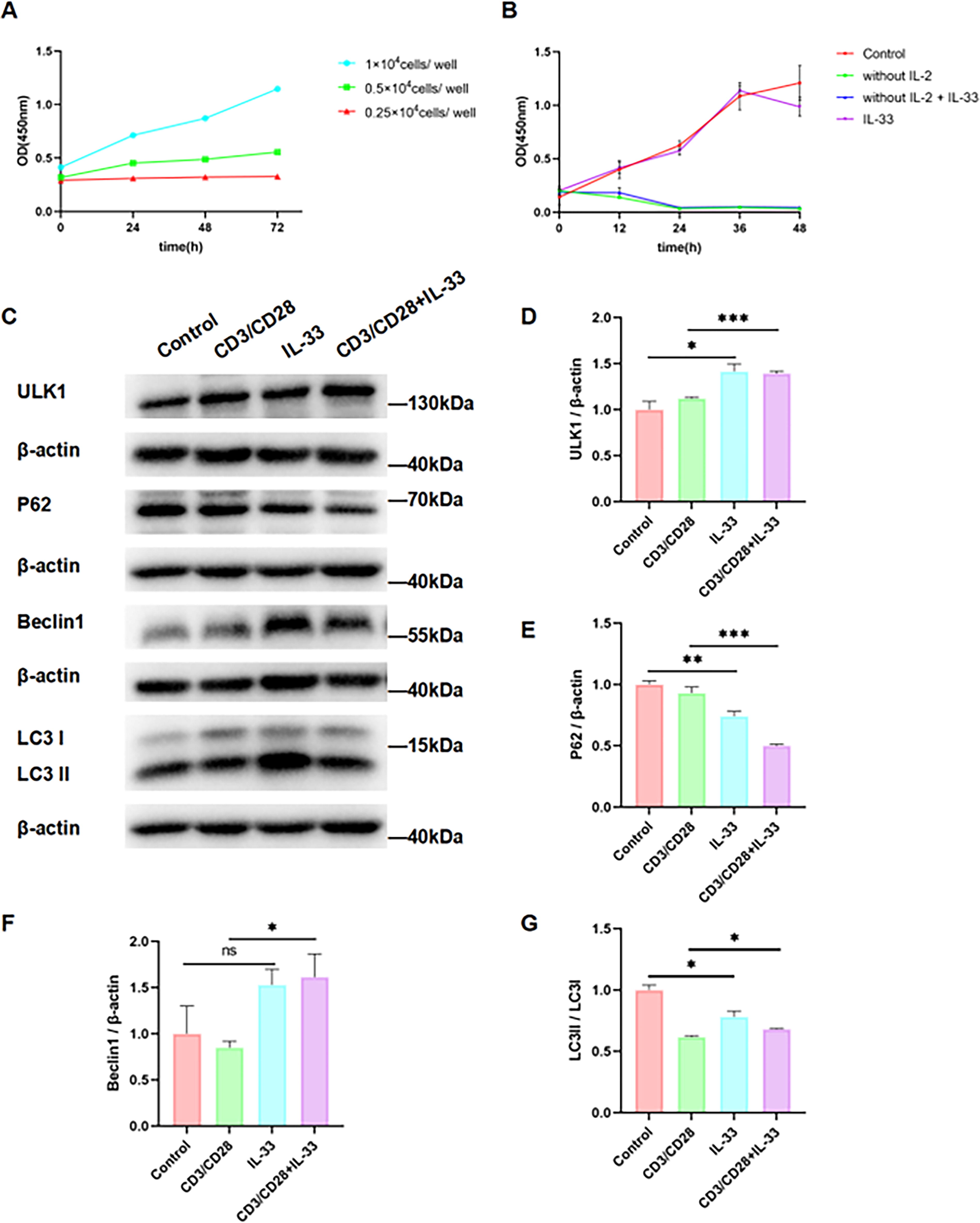
IL-33 promotes mitochondrial autophagy in CTLL-2 cells activated by CD3/CD28 antibodies.
IL-33 promotes the formation of autophagosome and autophagolysosome in CD3/CD28 antibody-activated CTLL-2 cells
Transmission electron microscopy was used to observe autophagolysosomes in each group of cells, and immunofluorescence was used to analyze whether there was co-localization between mitochondria and autophagosomes, and whether there was co-localization between mitochondria and lysosomes. It was found that CTLL-2 cells basically did not produce autophagolysosome in the resting state, and there was no co-localization relationship between mitochondria and autophagosomes, and no co-localization relationship between mitochondria and lysosomes. Autophagic lysosomes were observed in activated CTLL-2 cells, and there was a co-localization relationship between mitochondria and autophagosomes, as well as between mitochondria and lysosomes. When activated CTLL-2 cells were stimulated with IL-33, it was observed that a large number of autolysosomes were produced in the cells, and the co-localization relationship between mitochondria and autophagosomes, as well as mitochondria and lysosomes were enhanced (Fig. 4A–E). This suggests that CTLL-2 cells have little autophagy in the resting state. The level of autophagy was increased in activated CTLL-2 cells and further enhanced by IL-33 stimulation. We also used monodansylcadaverine to specifically label autophagosomes, and found that activated CTLL-2 cells produced more autophagosomes compared with resting CTLL-2 cells, and IL-33 stimulation further promoted this process (Fig. 4F), which was consistent with the conclusions drawn above.
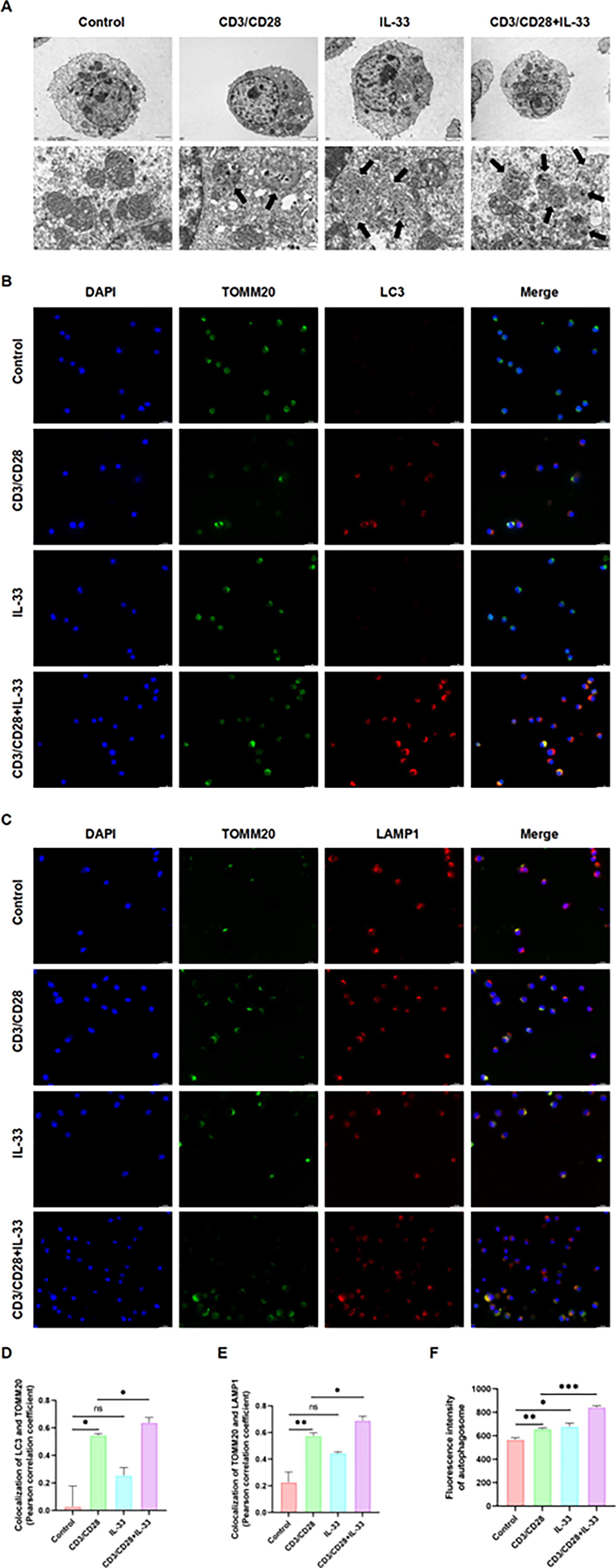
IL-33 promotes the formation of autophagosome and autophagolysosome in vitro.
IL-33 promotes the clearance of damaged mitochondria and the secretion of pro-inflammatory cytokines in CD3/CD28 antibody-activated CTLL-2 cells
Both resting and activated CTLL-2 cells produce low levels of superoxides and ROS during normal metabolism. The main source of superoxides and ROS is damaged mitochondria. After stimulating cells with IL-33 for 24 h, the levels of superoxide and ROS were reduced, suggesting that IL-33 promotes the clearance of damaged mitochondria (Fig. 5A–C). After the clearance of damaged mitochondria, in order to maintain the number of mitochondria, the healthy mitochondria will increase compensatively, so it was observed that the IL-33 stimulation group increased the production of ATP (Fig. 5D). We also measured the cytokine content in the supernatant of CTLL-2 cell culture medium. It was observed that the secretion of IFN-γ and TNF-α increased, and the secretion of IL-4 and IL-10 decreased after CTLL-2 cells were activated. When CTLL-2 cells were stimulated with IL-33 to activate the IL-33/ST2 signaling pathway, the secretion of IFN-γ and TNF-α was further increased, and the secretion of IL-4 and IL-10 was further decreased (Fig. 5E–H).

IL-33 promotes the clearance of damaged mitochondria and the secretion of pro-inflammatory cytokines in activated CTLL-2 cells.
Discussion
The liver and spleen are the main target organs for Lm infection and form the focus of infection (Vázquez-Boland et al., 2020). If the host immunity is strong, Lm is cleared by T cell-mediated immune responses. If the host immune system is weakened, Lm can spread and colonize throughout the body, attacking the brain or placenta and progressing to invasive listeriosis (Vázquez-Boland et al., 2001). In systemically infected mice, a small proportion of bacteria entered the spleen, and most bacteria entered the liver, where they were taken up by Kupffer cells (Ebe et al., 1999). It has been reported that 60% of intravenous Lm are rapidly cleared by the liver within 10 min (Gregory et al., 1996b), as a result of the combined action of Kupffer cells, neutrophils, migrating macrophages, and natural killer cells (Cousens and Wing, 2000). Natural killer cells can produce IFN-γ in response to IL-2 and IL-12, which induces macrophage activation (Gregory et al., 1996a; Humann and Lenz, 2010). Macrophages in the liver and spleen produce IL-6, IL-12, IL-1β, TNF-α, and nitric oxide, and inflammation ensues (Ehlers et al., 1992). The bacteria escape the siege and invade the central nervous system, resulting in hydrocephalus, cerebral hemorrhage, and brain abscesses (Charlier et al., 2018), which are extremely dangerous to the host. Because listeriosis has a high fatality rate and the incidence of sequelae, it is important to give appropriate treatment quickly. At present, the clinical treatment of listeriosis is mainly penicillin and other antibiotics (Hof et al., 1997). However, our results suggest that IL-33 can mitigate liver tissue damage caused by pregnant Lm infection by regulating mitochondrial autophagy and the function of immune cells, which has important implications for the prevention of tissue damage caused by listeriosis in pregnancy.
Pregnancy-associated listeriosis occurs mainly in the second and third trimesters. There are 2 main manifestations in pregnant women, one is non-specific obstetric signs (uterine contractions, labor, or fetal heart rate abnormalities), and the other is abortion. Lm rarely involves the central nervous system in healthy pregnant women, and only pregnant women with immunosuppression may have neurological changes (Charlier et al., 2020). Of course, maternal listeriosis may also be temporarily asymptomatic because pregnancy weakens the cell-mediated immune response (Khalil et al., 2023). During maternal infection, it is often difficult for the fetus or newborn to avoid infection. Fetal and neonatal mortality rates as high as 14.9% (de Noordhout et al., 2014). The main routes for newborns to acquire infection are vertical transmission, inhalation of infected amniotic fluid, or attack by vaginally colonized Lm (Dunphy et al., 2022). Studies have shown that maternal IL-33 levels increase during Lm infection (Li et al., 2022), which may be a new breakthrough point for changing maternal, fetal, or neonatal outcomes.
IL-33 has immunomodulatory effects on a variety of cells. It has been reported in many literature that IL-33/ST2 axis is an important factor affecting the process of sepsis. In a mouse model of sepsis, IL-33 is elevated in the blood circulation and acts on ST2 receptors on the surface of macrophages and type Ⅱ innate lymphoid cell (ILC2), promoting IL-10 secretion and Tregs cell expansion, ultimately leading to immunosuppression (de Lima et al., 2022; Nascimento et al., 2017). But at the same time, IL-33 promotes ILC2 to secrete IL-5, which can mediate the infiltration of neutrophils and monocytes in the lung during sepsis and is conducive to the clearance of bacteria (Xu et al., 2018).IL-33 can also activate mast cells and promote the secretion of IL-6 and TNF-α, thus having a chemotactic effect on neutrophils (Kumar, 2018). Extensive apoptosis of T lymphocytes occurs in sepsis, and IL-33 treatment can reduce apoptosis of T lymphocytes (Li et al., 2016). These findings strongly link the IL-33/ST2 axis to immune cells. In addition to the above immune cells, CD8+T lymphocytes are also important objects for IL-33/ST2 axis to play a regulatory role. It has been reported that in mice infected with lymphocytic choriomeningitis virus, IL-33 can promote the clonal expansion of cytotoxic T lymphocytes and exert an antiviral effect (Bonilla et al., 2012). Not only that, in melanoma, breast cancer, lung cancer, and other tumor diseases, IL-33 can also enhance CD8+T lymphocyte effector function to inhibit tumor growth (Gao et al., 2015; Xu et al., 2020). Our study aimed to investigate the impact of the IL-33/ST2 axis on the functional effects of CD8+T lymphocytes during perinatal listeria infection. We constructed a perinatal Lm infection model by injecting Lm through the tail vein and found that Lm infection would cause massive necrosis of liver tissue. Blocking IL-33/ST2 axis in mice by intraperitoneal injection of anti-IL-33Rα/ST2 antibody inhibited CD8+T cell infiltration and further aggravated liver damage. This is because CD8+T cells are the main immune cells that clear Lm. Blocking IL-33/ST2 axis inhibits the infiltration of CD8+T cells into the liver, preventing Lm in the liver from being cleared in time. As a result, Lm can damage the liver without restraint. In addition, our detection of cytokine levels in serum revealed that blocking the IL-33/ST2 axis could increase the levels of pro-inflammatory cytoines in serum and decrease the levels of anti-inflammatory cytokines, showing a pro-inflammatory effect. Inflammatory pathological reactions can further aggravate liver damage. This suggests that while IL-33 calls on one group of guards to kill the enemy bravely during Lm infection, it controls another group of guards not to hurt us.
The hypothesis that the IL-33/ST2 axis is associated with autophagy has been demonstrated in several studies. In allergic rhinitis, IL-33 released by nasal epithelial cells promotes mast cell degranulation by inhibiting autophagy. Blocking IL-33/ST2 pathway can activate mast cell autophagy, inhibit mast cell degranulation and inflammatory factor release, and alleviate symptoms (Nian et al., 2020). Another study on macrophages reached the same conclusion, finding that ST2-deficient macrophages had a significantly reduced number of mitochondria, while IL-33 overexpressing macrophages had a significantly increased number of mitochondria due to inhibition of mitophagy (Xu et al., 2021). However, a study in 2021 demonstrated that IL-33 inhibits mammalian target of rapamycin phosphorylation and promotes the expression of autophagy-associated proteins ULK1 and Beclin-1, thereby promoting autophagy in cardiomyocytes (Wu et al., 2021). At the same time, the conclusion that IL-33 promotes autophagy has also been verified in liver cells (Wang et al., 2021). Therefore, it is controversial whether IL-33 promotes mitophagy or inhibits mitophagy. Our current experimental study showed that IL-33 can promote mitophagy in CD8+T lymphocytes activated with CD3/CD28 antibody. In an animal model of listeria infection, CD8+T lymphocytes were activated in vivo, similar to CTLL-2 cells activated by CD3/CD28 antibodies in vitro. Based on the results of in vitro experiments, we speculated that IL-33 in vivo can act on activated CD8+T lymphocytes to promote the formation of ULK complex and initiate autophagy. Subsequently, a Beclin1-containing complex is formed to promote autophagosome isolation membrane formation. The cytoplasmic LC3I transforms into LC3II and integrates into the isolation membrane. The isolation membrane gradually extends to enclose the autophagy substrate P62 and the damaged mitochondria, forming the autophagosome. Next, the autophagosome fuses with the lysosome to form the autophagolysosome, where mitochondria and P62 proteins are degraded by proteolytic enzymes, and the final products are released into the cytoplasm for cell reuse. This is a complete mitochondrial autophagy process (Fig. 6).

Schematic diagram of mechanism by which IL-33/ST2 axis regulates mitochondrial autophagy in CD8+T lymphocytes.
Studies have shown that IL-33 is beneficial in bacterial infections such as Klebsiella, Clostridium difficile, and Staphylococcus aureus and can enhance host defense (Frisbee et al., 2019; Krishack et al., 2019; Ramirez-Moral et al., 2021). This is consistent with the protective effect of IL-33 in Lm infection. Immunomodulation targeting IL-33/ST2 axis may be an effective way to adjuvant treatment of perinatal Lm infection.
Footnotes
Acknowledgments
The authors thank Beijing Laboratory Animal Research Center for providing the experimental animal infection site and HFK BIOSCIENCE CO., LTD (Beijing, China) for providing C57BL/6J mice.
Authors’ Contributions
T.S.: Conceptualization, writing and methodology. Y.Y. and Y.L.: Methodology. N.X., F.C., and T.X.: Investigation and data analysis. H.W.: Supervision. Y.L. and C.Y.: Review and editing. All authors have read and agreed to the published version of the article.
Institutional Review Board Statement
The animal study protocol was approved by the Welfare Ethics Committee of Beijing Laboratory Animal Research Center (NO. BLARC-LAWER-202408001, approval date August 29, 2024).
Data Availability Statement
The data presented in this article are available.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research was funded by the Leading Talents in the Construction Project of High-Level Public Health Technical Talents in Beijing (2022–1-003), the National Natural Science Foundation of China (Grant No. 82371228).