
Abstract
Abstract
The objective of our study was to examine the pharmacology of the intraocular pressure (IOP)-lowering actions of the behaviorally inactive cannabinoids, abnormal cannabidiol (abn-CBD), and a cannabigerol analog, cannabigerol-dimethyl heptyl (CBG-DMH), in comparison to that of the nonselective cannabinoid 1 receptor (CB1R) and CB2R agonist, WIN55,212-2, in Brown Norway rats. The IOP was measured noninvasively using a hand-held tonometer in nonanesthetized animals. The IOP measurements were taken every 15 min for a period of 2 h after drug administration. All drugs were administered via intraperitoneal (i.p.) injections, and abn-CBD and CBG-DMH were also given topically. Both abn-CBD and CBG-DMH reduced IOP when administrated i.p. at doses of ≥2.5 mg/kg or topically at concentrations of 1%–2%. The IOP-lowering effects of abn-CBD and CBG-DMH were reduced by i.p. administration of O-1918 (2.5 mg/kg), a selective antagonist of the abn-CBD-sensitive cannabinoid-related receptor (CBx), but were unaffected by the CB1R antagonist, AM251 (2.5 mg/kg), or the CB2R antagonist, AM630 (2.5 mg/kg). In contrast, the IOP-lowering action of WIN55,212-2 was completely blocked by the CB1R-selective antagonist, AM251, and was unaffected by the CBx receptor antagonist, O-1918. However, similar to the nonpsychotropic cannabinoids, the ocular hypotensive actions of WIN55,212-2 were also insensitive to block by the CB2R antagonist, AM630. Consistent with this, the selective CB2R agonist, HU-308 (2 mg/kg) failed to reduce IOP in Brown Norway rats. Concurrent application of a dose of WIN55,212-2 that was subthreshold to reduce IOP (0.25 mg/kg), together with a topical dose of either abn-CBD (0.5%) or CBG-DMH (0.25%), respectively, potentiated the ocular hypotensive effect of either compound applied alone. This study demonstrates that the atypical cannabinoid, abn-CBD, and the cannabigerol analog, CBG-DMH, decrease IOP in the normotensive Brown Norway rat eye independent of CB1R or CB2R activation, via activation of CBx receptors. The enhanced decrease in IOP seen after coapplication of the CB1R agonist, WIN55,212-2, together with either abn-CBD or CBG-DMH, respectively, further suggests that the ocular pharmacodynamics of abn-CBD and CBG-DMH are mediated by receptor targets distinct from CB1R. These results indicate that both CBG-DMH and abn-CBD have the potential for further investigation as novel ocular hypotensive cannabinoids devoid of CB1R-mediated side-effects.
Introduction
In addition to CB1R and CB2R, studies using CB1R and CB2R knock-out mice have revealed the existence of additional putative non-CB1R/CB2R cannabinoid receptors present in the CNS,8–12 and at peripheral tissue sites, including the vasculature13,14 and immune cells. 15 Both the endocannabinoid, anandamide (AEA), and its synthetic analog, methanandamide, caused mesenteric endothelium-dependent vasodilation that was inhibited only by high concentrations of CB1R antagonist, SR141716A (>1 μM), and was not blocked by the specific CB1R and CB2R blockers, AM251 and AM630, respectively.14,16 Abnormal cannabidiol (abn-CBD), a synthetic analogs of the cannabinoid, cannabidiol, produces mesenteric vascular relaxation that, similar to AEA, is sensitive to antagonism by (-)-1,3-dimethoxy-2-(3-3,4-trans-p-methadien-(1,8)-yl)-orcinol (O-1918), an analog of cannabidiol. This abn-CBD-mediated vasorelaxation was insensitive to CB1R or CB2R selective blockers, and involved release of nitric oxide and activation of smooth muscle K+ channels, leading to relaxation of smooth muscle and vasodilation.9,11,14,16 Recent studies have suggested that the orphan receptors, GPR18 and GPR55, both of which can be activated by abn-CBD, may be candidates for the putative CBx receptor.17–19
Cannabinoids and endocannabinoids reduce intraocular pressure (IOP) in human and experimental animal models.20–22 This action is mediated locally in the eye and is independent of the CNS and effects on systemic blood pressure (BP). 23 Consistent with the pharmacological actions of cannabinoids on IOP, both CB1R and CB2R have been identified in tissues involved in aqueous humor secretion and outflow.24–26 Blocking of CB1R with specific antagonists eliminates much of the IOP- lowering effect of some cannabinoids, including the classical cannabinoids, such as Δ 9 THC, and the aminoalkylindole, WIN55,212-2.27–32 Apart from lowering IOP, cannabinoids have also been reported to exert neuroprotective actions in the retina and to increase ocular blood flow, consistent with additional retinal and vasculature sites of action. 33
Although most of the cannabinoid agonists tested so far are effective at reducing IOP, they have a relatively short duration of action, and chronic use of these compounds is limited by the behavioral and systemic side-effects they can produce, largely through actions at CB1R. Therefore, the use of behaviorally inactive cannabinoids that may act at receptor targets distinct from CB1R to exert therapeutically relevant pharmacological effects (e.g., lowering of IOP, vasodilation, neuroprotection, etc.) is desirable. The objectives of this study were (1) to examine the ocular hypotensive efficacy of 2 nonpsychoactive cannabinoids: abn-CBD, an atypical cannabinoid that is an agonist at the CBxR and cannabigerol-dimethyl heptyl (CBG-DMH), a dimethyl heptyl derivative of CBG 34 with ocular hypotensive and vasorelaxant properties 35 and (2) to compare the pharmacology of these cannabinoids with WIN55,212-2, which lowers IOP via actions at CB1Rs.28,31,32
Methods
Measurement of IOP
Brown Norway rats (300–400 g) were used for all experiments. Animals were housed under a 12-h light/dark cycle. Rats were maintained on chow and water at libitum except during experiments. All animals were treated in accordance with the standards and procedures set forth by the Canadian Council on Animal Care (CCAC) (www.ccac.ca) as well as the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. Protocols were approved by Dalhousie University Committee on Laboratory Animals. In accordance with the CCAC guidelines, all efforts were made to ensure that the animals experienced minimal suffering and pain and that the experimental design used the smallest number of animals required to obtain valid data.
The IOP was measured in nonanesthetized rats using a noninvasive method with a hand-held tonometer (Tono-Pen®XL; Solan Ophthalmic Products). Previous experiments using Brown Norway rats have shown an excellent correlation between IOP measurements made with a Tono-Pen XL and the more invasive transducer-measured IOP. 36 Before taking IOP measurements, the animals received 1 drop (40 μL) of 0.05% tetracaine eye drops (Dioptic Laboratories) for local anesthesia. Animals were then placed on a fixed bench and lightly restrained. For IOP measurements, the tonometer was placed gently against the anesthetized surface of the cornea. For each time point, IOP was measured 10 times with the mean value reported. Baseline or control IOP was measured before drug administration and all other IOP measurements were made every 15 min postdrug administration, for a total period of 2 h. All experiments were carried out at the same time in the morning to ensure that any changes in IOP readings were not related to diurnal variations in IOP. To achieve reliable IOP readings and to minimize stress, animals were handled and exposed to the IOP measurement procedure daily for at least 5 days before the experiments. In experiments using topical drug delivery, pressure measurements were made by using a TonoLab rebound tonometer (Innova) using the same experimental design as just described for the Tono-pen.
Drug preparation and administration
After control IOP measurements had been taken (time zero), animals were injected intraperitoneal (i.p.) with either abn-CBD (0.025, 0.25, 2.5, and 10 mg/kg) (Tocris), CBG-DMH (0.025, 0.25, 2.5, and 10 mg/kg), or WIN55,212-2 (2.5 mg/kg) (Tocris), dissolved in a 2-hydroxypropyl-β-cyclodextrin (HP-β-CD). The CBG-DMH was prepared by Y. Maor and A. Breuer (Hebrew University of Jerusalem, Jerusalem, Israel) based on published methods.34,37 The CB1R and CB2R antagonists, AM630 (Tocris) or AM251 (Tocris) and the antagonist for the CBxR, O-1918 (Cayman Chemical), were all dissolved in HP-β-CD and administered i.p. at 2.5 mg/kg, 30 min before the injection with any given agonists. For preparation of topical (0.25%–2%) cannabinoid solutions, abn-CBD or CBG-DMH was dissolved in Tocrisolve® (Tocris); and 5 μL of solution was applied to the cornea. Previous studies using Tocrisolve as a delivery vehicle for the psychotropic cannabinoid, WIN55,212-2, have demonstrated that topical ocular administration is well tolerated with no overt behavioral effects or systemic effects on BP or heart rate.31,38
Data analysis and statistics
The IOP data shown are presented as the IOP change (ΔIOP) calculated from the measured IOP minus baseline IOP at t=0. All data are presented as mean±standard error of the mean. Statistical analysis and curve fitting of the data were performed using Graphpad Prism version 4.0 (GraphPad Software Inc.). For calculation of ED50 for drug treatments, mean ΔIOP measured at 45 min for each drug treatment was fitted to a sigmoidal dose-response curve with a variable slope. Vehicle treatments were plotted at a concentration equal to one and a half log units less than the lowest drug treatment concentration tested. Unless otherwise indicated, statistical significance was determined by t-test when comparing means of only 2 groups; or by 1-way, or 2-way analysis of variance (ANOVA), as appropriate, when comparing the means of multiple treatment groups. Tukey's post-hoc analysis was used to determine differences among groups for one-way ANOVA, whereas Bonferroni's post-hoc analysis was used for 2-way ANOVA. P<0.05 was considered statistically significant.
Results
The ocular hypotensive actions of abn-CBD and CBG-DMH are independent of CB1R and CB2R activation
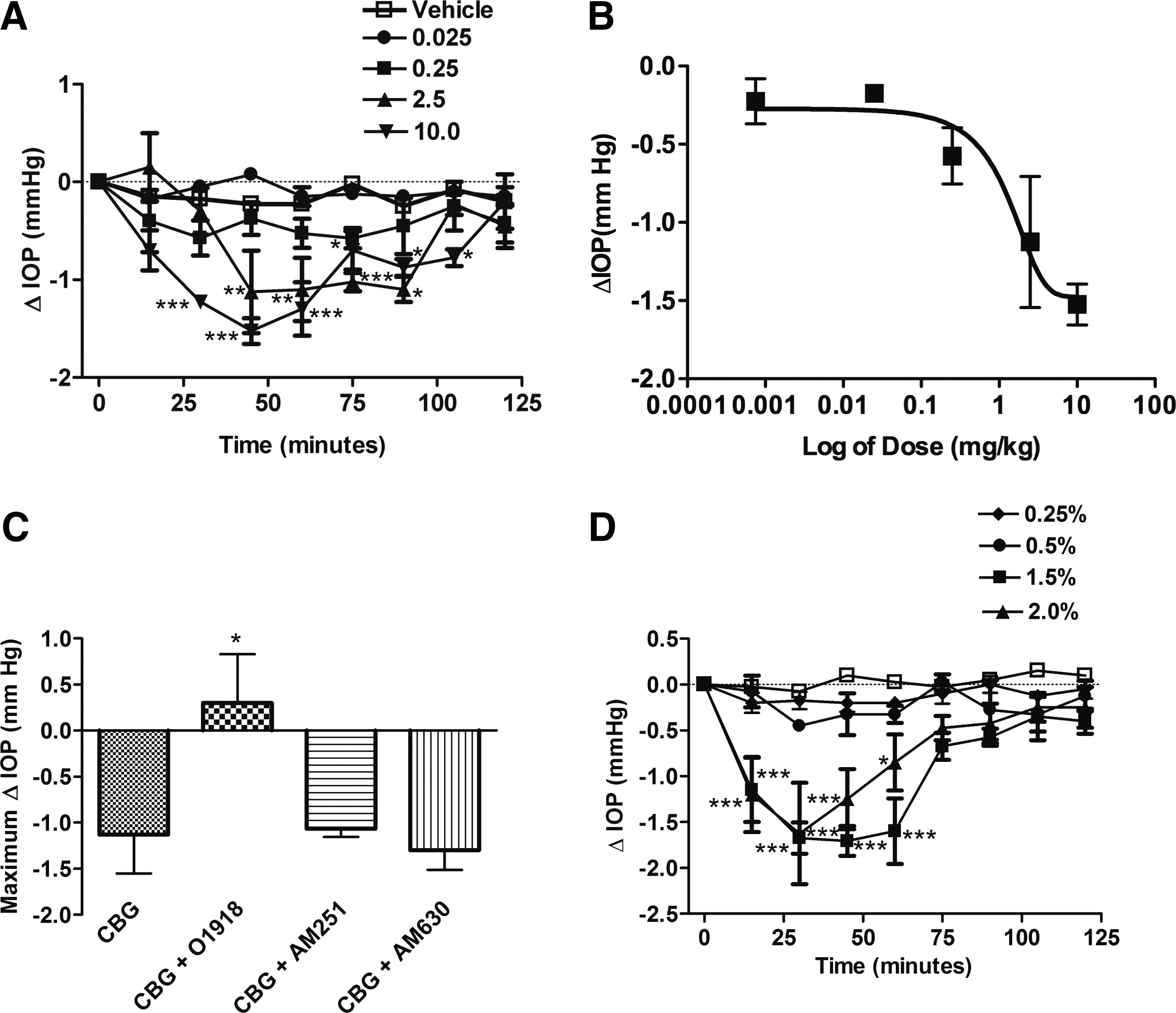
Figure 1A shows the effect of abn-CBD on IOP at doses of 0.025, 0.25, 2.5, and 10 mg/kg measured at 15 min intervals over a 2 h time-course after i.p. administration of a single dose of drug given at t=0. At doses of 0.025 and 0.25 mg/kg, abn-CBD did not produce a significant effect on IOP compared with vehicle control (P>0.05). However, at the higher doses of 2.5 and 10 mg/kg, there was a significant reduction in IOP at 15 (P<0.05), 30 (P<0.05), 45 (P<0.01), and 60 min (P<0.01) after the drug administration, as compared with vehicle control at the same time point. The IOP-lowering effect of abn-CBD was not significantly different from vehicle control (P>0.05) between 75 and 120 min after drug administration. Figure 1B shows the dose-response relationship for the change in IOP measured at 45 min after i.p. administration of abn-CBD (0.025–10 mg/kg). The calculated ED50 for the IOP-lowering effects of abn-CBD was 2.6 mg/kg. Figure 1C shows that the IOP-lowering effects of 2.5 mg/kg of abn-CBD were not blocked by i.p. preadministration of the CB1R or CB2R antagonists, AM251 (2.5 mg/kg) and AM630 (2.5 mg/kg), respectively. In contrast, administration of O-1918 (2.5 mg/kg, i.p.), an antagonist of CBxR, significantly blocked the IOP-lowering effect of abn-CBD. Figure 1D shows the effect of topical administration of 0.5%, 1.5%, and 2% of abn-CBD on IOP, measured at the same time intervals as for i.p. abn-CBD administration. However, topical application of 0.5% abn-CBD had no effect on the reduction of IOP. Higher concentrations of abn-CBD significantly reduced IOP at 15 (P<0.05 for 1.5%; P<0.01 for 2%), 30 (P<0.05 for 1.5%; P<0.01 for 2%), and 45 (P<0.01for 1.5%; P<0.05 for 2%) minutes after drug administration. Taken together, these results suggest that the pharmacological actions of abn-CBD on IOP may be mediated by distinct pharmacological targets independent of CB1R or CB2R that may include the CBx receptor.

We next examined the pharmacology of the cannabigerol analog, CBG-DMH, on IOP. Cannabigerol has been previously shown to produce a modest decrease in IOP after topical administration in cats, 39 but has minimal ocular hypotensive effects after topical administration in humans and nonhuman primates. 40 The DMH modification has generally been demonstrated to increase potency and efficacy of some cannabinoids. 41 We examined the effects of CBG-DMH (0.025, 0.25, 2.5, and 10 mg/kg i.p.) on IOP measured at 15 min intervals over a 2 h time course period as shown in Fig. 2A. As seen with abn-CBD, the lower i.p. doses of CBG-DMH (0.025 and 0.25 mg/kg) failed to produce a significant change in IOP compared with the vehicle control (P>0.05). However, i.p. administration of CBG-DMH at a dose of 2.5 mg/kg resulted in a significant reduction in IOP (P<0.01) within 45 min after drug administration. This decrease in IOP was sustained at 60 (P<0.01) and 75 min (P<0.001), and lasted up to 90 min (P<0.05) before returning toward control values. Higher i.p. doses of CBG-DMG were able to produce a significant drop in IOP (P<0.001) as early as 30 min after the drug administration, with a peak reduction in IOP occurring at 45 min (P<0.001). The drop in IOP produced by 10 mg/kg CBG-DMH i.p. was maintained up to 105 min (P<0.05). Figure 2B shows the dose-response relationship for the change in IOP measured at 45 min after i.p. administration of CBG-DMH (0.025–10 mg/kg). The calculated ED50 for the IOP-lowering effects of CBG-DMH was 3.15 mg/kg. To determine whether the IOP-lowering effect of CBG-DMH is mediated through the activation of CB1R and/or CB2R or, similar to abn-CBD occurred primarily via the CBxR, the CB1R antagonist, AM251 (2.5 mg/kg i.p.), and the CB2R antagonist, AM630 (2.5 mg/kg i.p.), were administered 30 min before administration of CBG-DMH (2.5 mg/kg i.p.). Similar to that observed with abn-CBD, both the CB1R antagonist, AM251, and the CB2R antagonist, AM630, failed to inhibit the ocular hypotensive action of CBG-DMH (Fig. 2C). However, when the animals were administered O-1918, the antagonist of the CBxR11,13,16 the IOP-lowering effect of CBG-DMH was completely abrogated (P<0.05). Figure 2D shows the effect of topical administration of 0.25%, 0.5%, 1.5%, and 2.0% of CBG-DMH on IOP, measured at the same time intervals as for i.p CBG-DMH administration. The lower concentrations, 0.25% and 0.5%, of topically applied CBD-DMH had no affect on the reduction of IOP (P>0.05). However, higher concentrations (1.5% and 2%) of CBG-DMG resulted in significant reductions in IOP at 15 (P<0.001), 30 (P<0.001), and 45 min (P<0.001). The reduction in IOP was still apparent at 60 min after administration of both 1.5% (P<0.05) and 2% (P<0.001) CBG-DMH. Taken together, these data indicate that CBG-DMH administered i.p. or topically can significantly lower IOP. Similar to abn-CBD, the IOP-lowering actions of CBG-DMH appear to be largely mediated independent of CB1R and CB2R and may involve the CBxR.
The IOP-lowering effects of WIN55,212-2 are mediated by CB1R
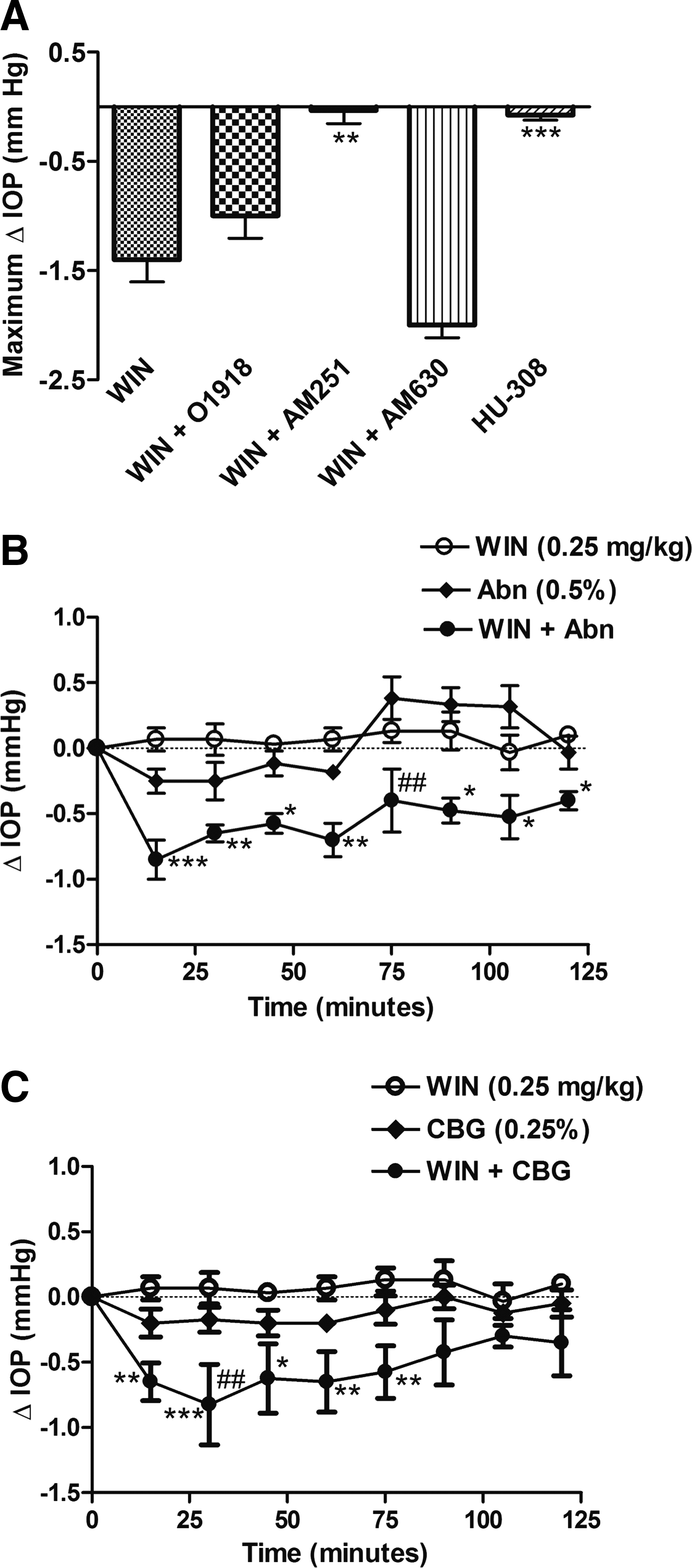
We next compared the pharmacology of the IOP-lowering actions of nonpyschotropic cannabinoids, abn-CBD, and CBG-DMH with that of the aminoalklindole, WIN55,212-2, a mixed CB1R/CB2R agonist that has been previously been reported to lower IOP in humans, 11 nonhuman primates, 42 and rodents.28,31,32 To confirm whether the WIN55,212-induced decrease in IOP observed in Brown Norway rats is mediated primarily by CB1R, the effects of the CB1R antagonist, AM251 (2.5 mg/kg), the CB2R antagonist, AM630 (2.5 mg/kg), and the CBxR antagonist, O-1918, were tested on the WIN55,212-2-induced IOP decrease. The effect of the selective CB2R agonist, HU-308 was also tested. Figure 3A shows that only the CB1R antagonist, AM251, significantly blocked the IOP decrease produced by i.p. administration of 2.5 mg/kg WIN55,212-2 (P<0.01). In keeping with the lack of block of the WIN55,212-2-induced decrease in IOP by the CB2R antagonist, AM630, the CB2R agonist, HU-308 (2.5 mg/kg), also failed to reduce IOP (P<0.001 compared with WIN55,212-2). These data support previous findings demonstrating that WIN55,212-2 lowers IOP via activation of CB1R,28,31,32 and are in contrast to the pharmacology of the nonpsychotropic cannabinoids, abn-CBD and CBG-DMH, which appear to mediate their IOP-lowering actions via a O-1918-sensitive receptor target distinct from both CB1R and CB2R.
The IOP-lowering effects of WIN55,212-2 are enhanced in combination with abn-CBD and CBG-DMH
To determine the effect of combination treatment, abn-CBD or CBG-DMG, were administered together with WIN55,212-2, at doses that individually failed to produce a significant decrease (P<0.05) in IOP. Figure 3B shows the effect of administration of WIN55,212-2 (0.25 mg/kg) and abn-CBD (0.5%) on IOP in Brown Norway rats. Administration of WIN55,212-2 or abn-CBD alone failed to produce a significant drop in IOP over the 120 min time course compared with i.p. vehicle or topical vehicle (P>0.05, vehicle data not shown), respectively. However, concurrent administration of both WIN55,212-2 and abn-CBD produced an enhanced drop in IOP at all time points from 15-120 min (P<0.001 at 15 min; P<0.01 at 30 and 60 min; P<0.05 at 45, 95, 105, 120 min), compared with WIN55,212-2 alone, and at 75 min (P<0.01) compared with abn-CBD alone.
Examination of the effects of CBG-DMH (0.25%) and WIN55,212-2 (0.25 mg/mg) (Fig. 3C) also demonstrated that although both compounds individually at these doses did not produce a significant drop in IOP compared with vehicle (P>0.05, vehicle data not shown), concomitant application of CBG-DMH and WIN55,212-2 together produced an increased drop in IOP at 15 (P<0.01), 30 (P<0.001), 45 (P<0.05), 60 (P<0.01), and 75 (P<0.01) minutes compared with WIN55,212-2 alone, and at 30 min (P<0.0001) compared with CBG-DMH alone.
Discussion
The primary findings of this study are that the behaviorally inactive cannabinoid analogs, abn-CBD, and CBG-DMH, given either via i.p. or topical routes, are able to reduce IOP in normotensive Brown Norway rats. The IOP-lowering actions of abn-CBD and CBG-DMH were independent of CB1R and CB2R activation, as indicated by lack of antagonism by CB1R or CB2R blockers, and were mediated via a receptor target that is susceptible to O-1918 antagonism. The pharmacology of abn-CBD and CBG-DMH in reducing IOP was distinct from that of the psychotropic synthetic cannabinoid agonist, WIN55,212-2, which acts at ocular CB1R to produce a reduction in IOP.28,31,32 The enhanced actions of the nonpsychotropic cannabinoids on the magnitude and duration of the IOP decrease when administered together with WIN55,212-2, at doses that individually failed to produce a significant IOP-lowering effect, supports distinct receptor targets. These results provide evidence for additional novel non-CB1R/CB2R receptor targets for nonpsychotropic cannabinoids in ocular tissue that could be exploited therapeutically for treating ocular disease.
The ocular endocannabinoid system and IOP
In keeping with the pharmacological effects of marijuana in the eye43–45 a number of studies have now demonstrated that cannabinoids including the classical phytocannabinoid Δ 9 THC,29,39,46–48 the synthetic cannabinoids CP5595027 and WIN55,212-2,30,31,42 the endocannabinoids, 2-arachidonoyl glycerol and noladin ether, 49 as well as inhibitors of AEA degradation, 50 all decrease IOP via actions that are mediated locally and independent of the CNS and may involve cannabinoid receptors.20,22 For example, the IOP-reduction produced by Δ 9 THC, 29 CP55940, 27 and WIN55,212-228,31,32 were all reduced by CB1R-selective antagonists. In support of this, CB1R mRNA and protein have been identified in humans, nonhuman primates, and rodents in anterior ocular tissues involved in aqueous secretion and outflow pathways,26,51 whereas CB2R mRNA has been reported in the rat retina. 52 Most cannabinoids tested to date appear to exert their IOP-lowering actions via CB1R activation.24,27,28,31,32,51,53 The mechanisms responsible for the CB1R decrease in IOP have been suggested to involve a decrease in aqueous humor production, 42 which may occur via CB1R-mediated vasodilation of blood vessels in the ciliary body 54 and/or an increase in outflow of aqueous humor mediated by contraction of the ciliary muscle affecting trabecular meshwork resistance. 55 In addition, the direct actions of cannabinoids on trabecular meshwork (TM) may modify outflow. For example, CP55,940 has been shown to relax bovine TM strips, an effect that was mediated by enhancement of a high conductance Ca2+-activated K+ current. 56 Further, the actions of WIN55,212-2 at CB1R in human TM cells results in activation of both a Gq-coupled pathway, leading to phospholipase C (PLC)-dependent increases in Ca2+, and increases in Gi-coupled MAPK signaling, both of which may affect trabecular outflow resistance via activation of Ca2+-dependent ion channels and alterations in TM extracellular matrix. 5 Cannabinoids have also been proposed to act via non-CBR mechanisms to decrease IOP. These actions may involve increased production of COX-2, together with the resultant products, prostaglandin E2 and matrix metalloproteinases-1, -3, 57 all of which have been implicated in outflow pathway remodeling. 58
Our studies have demonstrated that the atypical cannabinoid, abn-CBD, and the cannabigerol analog, CBG-DMH, are able to produce a reduction in IOP in rats that is independent of CB1R or CB2R antagonism. The IOP decrease seen with these cannabinoids was sensitive to blocking by O-1918, an antagonist at the CBxR; O-1918 has been shown to reduce the hypotensive actions of the endocannabinoid, N-arachidonoyl ethanolamide (AEA), as well as those of abn-CBD acting at abn-CBD-sensitive endothelial receptors in anesthetized rats in vivo, 59 and in vitro experimental models.60,61 The O-1918 has also been reported to block the actions of abn-CBD and the endogenous lipid, N-arachidonoyl glycine, on CNS microglial cell migration that are mediated by the orphan receptor, GPR18. 62
Although the specific receptor target(s) and site of action for the ocular hypotensive actions of abn-CBD and CBG-DMH are not yet identified, the enhanced decrease in IOP seen when these cannabinoids are combined with the CB1R agonist, WIN55,212-2, supports distinct non-CB1R targets. This may involve CBx receptors at conventional outflow tissue sites, including the TM as well as endothelial cells of the efferent blood vessels of the anterior uvea, to enhance aqueous humor outflow. In support of this, a preliminary in vitro study using a porcine anterior segment perfused organ culture model demonstrated that abn-CBD increases aqueous humor outflow through the TM pathway. 63 Additionally, although acute topical cannabigerol produced little effect on IOP in humans and rabbits, 40 an in vivo study of the actions of chronic topical cannabigerol administration in the eyes of cats demonstrated that this compound produced a reduction in IOP via an increase in aqueous humor outflow facility with no action on aqueous humor formation. 39 The nonpsychotropic cannabinoid, HU-211 (dexanabinol), a synthetic analog of the classical cannabinoid, Δ 9 THC, which exhibits little binding affinity for CB1R compared with Δ 9 THC, 64 has also been reported to lower IOP in rabbits independent of alterations in the rate of aqueous inflow. 65 Pharmacological studies carried out using HU-211 have indicated that, similar to Δ 9 THC, 66 part of the ocular hypotensive actions of this cannabinoid may involve interactions with the adrenergic system, although the exact mechanisms underlying this interaction as well as the involvement of cannabinoid, or other noncannabinoid receptors, remains to be clarified.
Nonpsychotropic cannabinoids as ocular therapeutics
Currently available treatments for glaucoma, an optic nerve neuropathy for which IOP is a primary risk factor, include β-adrenergic blockers, prostaglandin analogs, carbonic anhydrase inhibitors, and α2-adrenergic agonists. Many of the patients suffering from this disease frequently use 2 or 3 drugs to control their IOP. 67 Nonpsychoactive cannabinoids, administered as monotherapy or adjunctive therapy, may provide an alternative treatment for those who do not respond to available therapies, or who require additional drugs to control their IOP. Both abn-CBD and CBG-DMH were effective ocular hypotensives when acutely administered topically, and their ability to decrease IOP independent of CB1R avoids many of the systemic and CNS side-effects associated with those cannabinoids that activate CB1R. 68 In addition, an enhanced ocular hypotensive action and increased duration of drug effect is seen with a combination of a low doses of the potent CB1R agonist, WIN55,212-2, and either abn-CBD or CBG-DMH, which are devoid of CB1R activity. This suggests that nonpsychotropic cannabinoids may be useful in combination therapy with psychotropic cannabinoids or other glaucoma medications, 32 to produce ocular hypotension at lower doses with reduced side-effects.
This study used only a single dose of abn-CBD or CBG-DMH in normotensive rats, which resulted in a drop in IOP of <2 h duration. However, studies of psychotropic cannabinoids, such as WIN55,212-2, delivered chronically in glaucoma patients refractory to conventional therapy, 30 nonhuman primates with glaucoma, 42 or experimental animals with ocular hypertension,20–22,31,32 resulted in a sustained drop in IOP. Since WIN55,212-2, abn-CBD, and CBG-DMG had comparable durations of action in our study, this suggests that chronic dosing with the nonpsychotropic cannabinoids, such as WIN55,212-2, may be effective in producing a more sustained decrease in IOP in ocular hypertensives. 32
In addition to their IOP-lowering properties, cannabinoids exert other beneficial effects in the eye, which may be relevant in the treatment of glaucoma.20,22,69 For example, both behaviorally active Δ 9 THC as well as the nonpsychoactive cannabinoid, cannabidiol, have been shown to provide neuroprotection of retinal ganglion cells (RGCs) after glutamate-induced excitotoxicity and ischemia caused by elevated IOP.70–73 Further, the behaviorally inactive cannabinoid, HU-211, has a neuroprotective effect against injury-related optic nerve damage.20,74,75 Although abn-CBD and CBG-DMH have not yet been fully investigated for their neuroprotective potential in models of retinal neurodegeneration, abn-CBD has been reported to increase RGC survival and inhibit activated retinal microglia after axotomy. 76 In other studies, abn-CBD was shown to interact with abn-CBD-sensitive receptors on microglial cells in an organotypic hippocampal slice culture, after excitotoxic shock induced by N-methyl-D-aspartic acid (NMDA), to reduce microglial cell activation and neuronal degeneration. 77
Taken together, this study provides novel findings demonstrating that the behaviorally inactive cannabinoids: the atypical cannabinoid, abn-CBD, and the cannabigerol analog, CBG-DMH, can lower IOP via mechanisms distinct from CB1R or CB2R activation. These actions may involve a distinct cannabinoid-related CBxR localized to anterior ocular tissues involved in aqueous humor outflow. Presence of abn-CBD-sensitive receptors on endothelial cells and other retinal cells, including microglial cells, may provide additional therapeutic targets in the design of comprehensive drug treatments for glaucoma and chronic retinal disease.
Footnotes
Acknowledgments
This work was supported by the Canadian Institute of Health Research grants IAO-13484 and MOP-97768 (MEMK). The authors wish to thank Dr. Raphael Mechoulam for comments on the manuscript, Dr. Aviva Breuer for providing CBG-DMH, and Christine Jollimore for expert technical assistance.
Author Disclosure Statement
No competing financial interests exist.