
Abstract
Abstract
Purpose:
This study aimed to investigate the effects of anandamide or arachidonylethanolamide (AEA), an endogenous cannabinoid receptor agonist, on intraocular inflammation in an endotoxin-induced uveitis (EIU) model in rabbits.
Methods:
Forty New Zealand albino male rabbits were used (5 groups, 8 animals in each). After establishment of sufficient anesthesia, animals were taken under surgery for intravitreal injections. A maximum amount of 50 μL of solution was injected into the central vitreous with a 30-gauge needle. In the control group, sterile saline was injected into the right eyes of the animals. Likewise, AEA (10−5 M) in the second group, lipopolysaccharide (LPS; 100 ng) in the third group, and AEA (10−5 M) and LPS (100 ng) in the fourth group were administered. Fifth group received 0.1 mL subtenon injection of AM251 (10−5 M), a CB1-receptor antagonist, 30 min prior to intravitreal LPS (100 ng) and AEA (10−5 M) injection. At 24 h after the surgical intervention, clinical evaluation was performed and animals were then euthanized with 100 mg/kg intravenous pentobarbital injections. Immediately after the induction of pentobarbital anesthesia, the anterior chamber of the eyes was quickly punctured using a 30-gauge needle to drain aqueous humor (AH) and obtained specimens were used for cell count, protein measurement, and microbiological contamination tests. After AH collection, enucleation was performed and enucleated material was kept for the pathological evaluation.
Results:
AEA caused an overall worsening of EIU in studied eyes. It significantly increased the detrimental effects of endotoxin, as assessed by clinical investigation of ocular inflammation, AH leukocyte content, and AH protein concentrations. CB1-receptor antagonist AM251 administration reversed some components of this AEA-induced exacerbation to significant extents.
Conclusion:
AEA exacerbated EIU in rabbit eyes. AM251 has been found beneficial to prevent AEA's aggravating impact on EIU. As AEA is a treatment choice for lowering intraocular pressure in ophthalmology practice, concurrent use of CB1-receptor antagonists may be a questionable strategy in cases of secondary glaucoma, to avoid aggravation of the present inflammation.
Introduction
Endotoxin-induced uveitis (EIU) is an experimental model for human acute anterior uveitis, produced by intravitreal, intraocular, intravenous, intraperitoneal, or foot paw injections of lipopolysaccharide (LPS). 3 LPS is capable of inducing uveitis in a dose-dependent manner in a number of species including rabbits, rats, and mice, which is characterized by miosis, iris hyperemia, breakdown of the blood-aqueous barrier, subsequently leading to extravasation of vascular fluid, protein leakage, and massive inflammatory cell infiltration into the anterior chamber (AC), and vitreous of the eye.3–7
Cannabinoids, psychoactive constituents of marijuana, have psychotropic and pharmacological effects. 8 Known effects of cannabinoids are muscle relaxation, immunosupression, sedation, bronchodilation, neuroprotection, analgesia, stimulation of appetite, anti-emetic action, and induction of apoptosis in cancer cells.9–11 Many of these effects are due to activation of specific cannabinoid receptors, but some are related to receptor-independent mechanisms. 12
The identification of cannabinoid receptors was followed by the detection of endogenous ligands of these receptors. These ligands are arachidonylethanolamide or anandamide (AEA) and 2-arachidonylglycerol, mainly produced from the major precursors N-arachidonoyl-phosphatidylethanolamine and diacylglycerols with arachidonic acid (AA) on the 2-position, respectively. 13 The presence of ethanolamine and arachidonic acid, a family of endogenous lipids,14–17 at adequate amounts in various tissue types can yield to AEA synthesis by the activity of the enzyme formerly called AEA synthase, which later turned out to be AEA amidohydrolase.18–20 Similar to Δ9-tetrahydrocannabinol and other ligands of cannabinoid receptors, AEA is known to cause various cannabinomimetic actions; however, studies on the pharmacological effects of AEA on ocular structures are still not very high in number. 21 Therefore, we aimed to investigate the effects of AEA on intraocular inflammation by means of an EIU model in rabbits.
Methods
New Zealand albino male rabbits weighing 2–2.5 kg were used in this study. The animals were housed under a 12-h light–dark cycle at room temperature and were fed on a standard diet until the experiment date. The food was withdrawn from the cages at 24 h before the experiments, but the animals were allowed to reach tap water ad libitum. Criteria of the Association for Research in Vision and Ophthalmology (ARVO) resolution about the use of animals in research were applied to all animals. Rabbits were anesthetized with xylazine (5 mg/kg) and ketamine (37.5 mg/kg). After establishment of anesthesia, animals were given injections. All injections were performed under a biomicroscope with utmost attention to avoid any possible microbial contamination. Only 1 eye (right) of the animals was used for the study. In the first group (control), sterile saline was injected into the right eyes of animals. Likewise, AEA (10−5 M) in the second group, LPS (100 ng) in the third group, and AEA (10−5 M) and LPS (100 ng) in the fourth group were administered. In our previous trials, not any clinically observable variation was detected between lower (10−6 M) and higher (10−4 M) concentrations of AEA; therefore, 10−5 M concentration was used throughout the entire experiment. In the fifth group, animals received 0.1 mL subtenon injection of AM251 (10−5 M), a CB1 receptor antagonist, 30 min prior to intravitreal LPS (100 ng) and AEA (10−5 M) injections. All solutions were freshly prepared before the injections. AEA was first dissolved in anhydrous ethanol to be at a concentration of 10−2 M and further diluted with sterile saline. LPS was dissolved in phosphate buffer solution and sterile saline was used to achieve 2 and 4 mg/mL concentrations, to be used for injections of the LPS alone and AEA+LPS groups, respectively. AM251 was first dissolved in ethanol and diluted to 10−5 M concentration with sterile saline. A maximum amount of 50 μL of solution was injected into the central vitreous with a 30-gauge needle, taking care to avoid lens damage.
After macroscopic examination of the eyes, slit-lamp examinations were performed by 2 blinded investigators. At 24 h after the surgical procedure, animals were euthanized with 100 mg/kg intravenous pentobarbital injections. Immediately after the induction of pentobarbital anesthesia, the AC of the eye was quickly punctured using a 30-gauge needle to drain aqueous humor (AH) and obtained specimens were used for cell count, protein measurement, and microbiological contamination tests. After peritomy, enucleation was performed and enucleated material was kept in 10% formalin for the pathological evaluation.
The intensity of the intraocular inflammation was graded using a clinical scoring system as previously described by Mikawa et al. 21 (Table 1). To avoid clotting, 5% ethylenediaminetetraacetic acid was added to the AH sample at 9:1 ratio. To detect any possible contamination, AH sample was plated in blood agar, Eosin-methylene blue agar (EMB), and Sabouraud dextrose agar. One microliter of aqueous aspirate was placed on a glass slide and stained by Wright's method for the confirmation of cell nature. The total number of inflammatory cells in each 100 μL of samples was then counted using a Thoma glass under a microscope with 400× magnification. The remaining aqueous sample was reserved for protein assessment. Protein concentrations were determined by Lowry method. 22 The enucleated eye specimens were trimmed through anterior to posterior from the pupil toward the optic disc and then the lens was gently removed for pathological examination. Trimmed serial corneal eye sections were fixed in 10% formalin for 72 h and embedded in paraffin, and 5-μm tissue sections were taken and stained with hematoxylin–eosin staining. Pathological findings assessed in the uvea and cornea were scored according to Tilton's description 23 as follows: (0) normal tissue; (1+) dilatation of iris vessels and thickened iris stroma with exudate, protein, and/or a few scattered inflammatory cells in the AC; (2+) infiltration of inflammatory cells into the stroma of the iris and/or ciliary body with a moderate number of inflammatory cells within the AC; (3+) heavy infiltration of inflammatory cells within iris stroma and ciliary body and a heavy infiltration of inflammatory cells within the AC; and (4+) heavy exudation of cells and dense protein aggregation in the AC and inflammatory cells deposits on the corneal endothelium.
AC, anterior chamber.
Drugs used in this study were xylazine (Alfazyne; Alfasan Int.B.V.), ketamine (Ketalar; Eczacıbaşı İlaç.San.), AEA (Sigma), LPS (Escherichia coli, 0111, B4; Sigma), AM251 (Tocris), and pentobarbitale (Pental, I.E.Ulagay İlaç San.).
Kruskal–Wallis variance analysis and Mann–Whitney U tests were used to determine the variance among groups and statistical significance between the groups. P<0.01 was considered significant with regard to Bonferroni correction for post hoc tests.
Results
Microbiological culture examinations were negative in all groups; thus, infectious uveitis and probable contaminations during injections were ruled out. In the control group, no noticeable inflammatory sign, except for a mild response characterized by conjunctival hyperemia, was observed.
On clinical evaluation, the eyes obtained from AEA-injected animals showed a moderate response with a clinical score from 1 to 4. Ocular inflammation scores were significantly higher in the LPS, AEA+LPS, and AM251+AEA+LPS groups (Fig. 1). In these groups, hyperemia of iris and conjunctiva, accompanied by exudation in AC, was excessively observed during clinical examination (Fig. 2). Ocular inflammation scores and histopathologic grades for each animal are presented in Table 2.

Ocular inflammation scores based on clinical evaluation of the studied eyes. Data are presented as medians. Kruskal–Wallis variance analysis and Mann–Whitney U post hoc test were used. *P<0.01 was considered significant with regard to Bonferroni correction. AEA, anandamide; LPS, lipopolysaccharide.

Control (left) versus the AEA+LPS group. Conjunctival hyperemia, 4+ anterior chamber cells, and anterior chamber flare in the eye of the AEA+LPS group. Arrowheads indicate the 4+ anterior chamber flare.
Values are given as clinical grade/histological grade.
C, control; AEA, anandamide; LPS, lipopolysaccharide; AEA+LPS, anandamide+lipopolysaccharide; AM251+AEA+LPS, AM251 (CB1-receptor antagonist)+anandamide+lipopolysaccharide.
Leukocyte content of AH was reasonably low in both the control and AEA groups and cell numbers did not differ significantly (Fig. 3). Mean leukocyte content of AH was significantly high in the LPS (820±422.8 cell/mm3) group in comparison with the control and AEA groups. AEA caused a dramatic increase in leukocyte numbers in AH (1,543.8±295.7 cell/mm3) when administered with LPS (AEA+LPS group). CB1 antagonist AM251 pretreatment (AM251+AEA+LPS group) completely reversed this intense increase in cell numbers (498.8±134.6 cell/mm3).

Aqueous humor leukocyte counts of the groups. AEA did not cause a remarkable change in cell numbers. AM251 completely withdrew AEA's impact on LPS-induced cell accumulation in AH to levels even lower (not significant [ns]) than that of the LPS group. Data are expressed as medians. Kruskal–Wallis variance analysis and Mann–Whitney U post hoc tests were performed. *P<0.01 was considered significant with regard to Bonferroni correction. AH, aqueous humor.
The mean protein concentration of AH in the control group was 0.97±0.9 mg/mL. In the AEA group, AH protein levels were elevated with respect to control values. LPS administration induced a remarkable increase in AH protein content (32.38±3.18 mg/mL), whereas AEA significantly intensified the effect of endotoxin up to 3-fold. In a similar manner to cell counts, AM251 completely reversed this augmentation produced by AEA. The LPS and AM251+AEA+LPS groups did not significantly differ from each other (Fig. 4).

AH protein concentrations. AEA caused an increase in protein levels when applied alone. The drastic effect of AEA on LPS-induced protein increase was completely reversed by AM251. Data are expressed as medians. Kruskal–Wallis variance analysis and Mann–Whitney U post hoc tests were performed. *P<0.01 was considered significant with regard to Bonferroni correction.
When histopathologically examined, there was no indication of inflammation in control eyes, except for focal unilateral neutrophil leukocyte infiltrations at the iridocorneal angle nearby the site of injection. This finding has been detected in all groups and interpreted to be a result of surgical intervention. In the AEA group, vascular changes including widespread edema in the stroma and congestion of arterioles and venules were mainly observed in the iris, ciliary body, and ciliary processes. In some of the cases, capillaries located at the top of the ciliary processes were severely congested (Fig. 5). A limited number of inflammatory cells in the AC and posterior chamber (PC) were present as well. In the LPS group, the lesions consisted of focal neutrophil leukocyte and macrophage infiltrations from moderate to severe degree and mild hyperemia in the iris and ciliary body (Fig. 6). Numerous inflammatory cells occupying AC were observed. In one of the eyes, PC, vitreous, retina, and optic disc were also affected with intense neutrophil leukocyte infiltration. In the AEA+LPS group, both vascular and inflammatory changes throughout the iris, ciliary body, and processes were typical for uveitis. Hyperemia, perivascular edema, multifocal hemorrhagies, and intense focal neutrophil leukocyte accumulations in iris and ciliary body were the main findings (Fig. 7). Inflammatory reaction seemed to be more severe in this group when compared with the LPS group. Capillary dilatations were not as prominent as those observed in the AEA group. The middle arteries were usually hyperemic and many clusters of neutrophil leukocytes with 50 or more cells in each were detected in spaces between the 2 reciprocal ciliary processes in AC. Eyes obtained from the AM251+AEA+LPS group also had neutrophil leukocyte infiltrations in AC and PC. In this group, vascular changes and the signs of inflammation in the uvea were less prominent when compared with the AEA+LPS group by microscopic examination. The results of histopathological assessment based on Tilton's description 23 are shown in Fig. 8 for all groups.
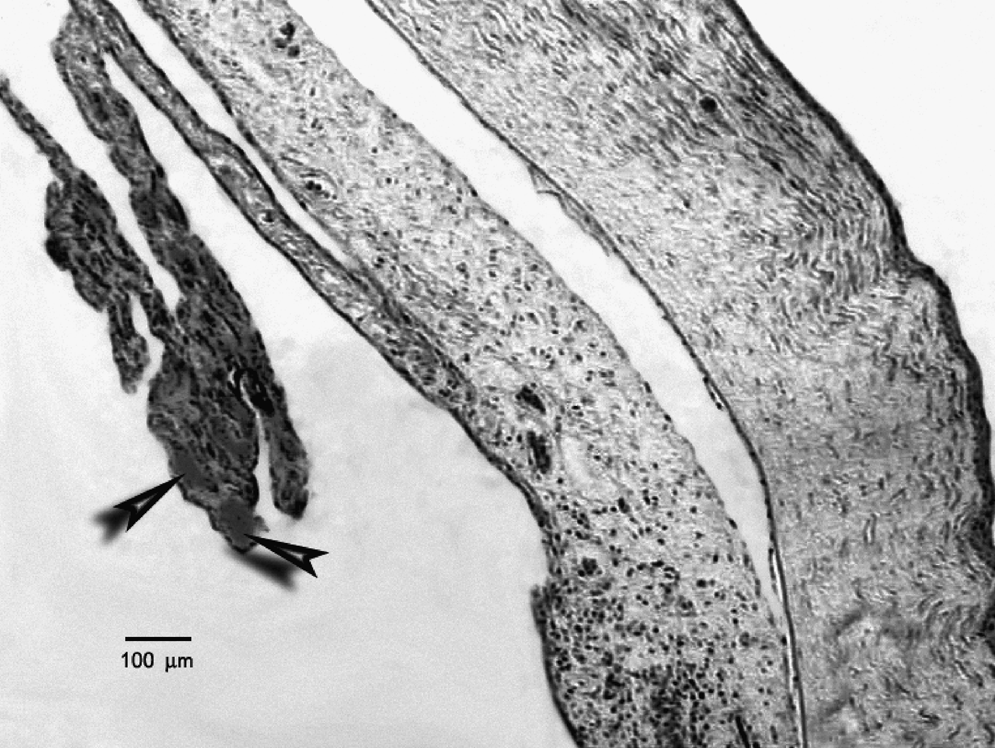
Pathological section of an eye from the AEA group. Arrowheads indicate the dilated and severely congested vessels. Note widespread edema in the iris, ciliary body, and ciliary processes. Hematoxylin–eosin staining; scale bar= 100 μm.

Pathological section of an eye from the LPS group. Dense neutrophil leukocyte infiltration (arrowheads) in choroidea is seen. Hematoxylin–eosin staining; scale bar=100 μm.
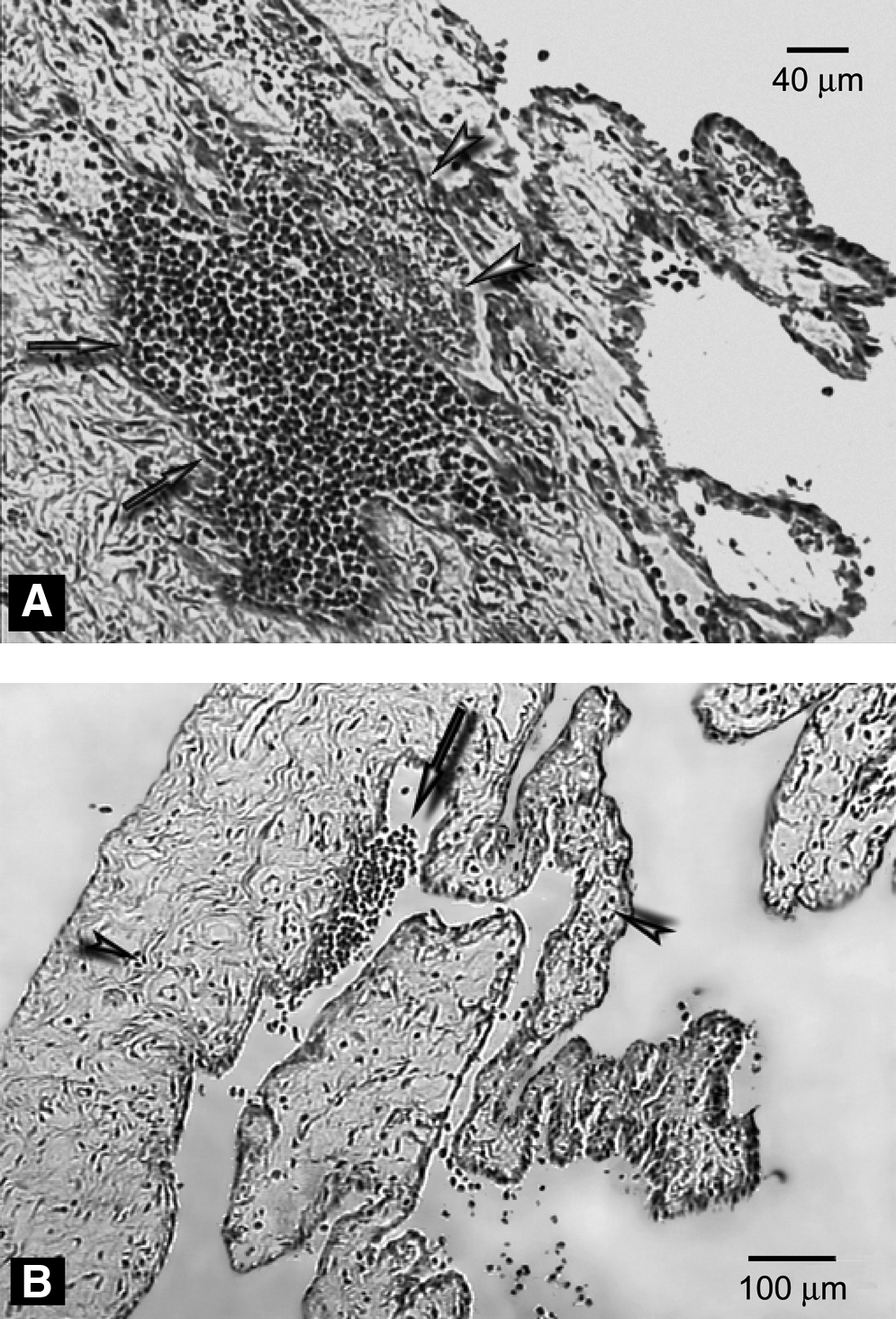
Pathological section of an eye from the AEA+LPS group.
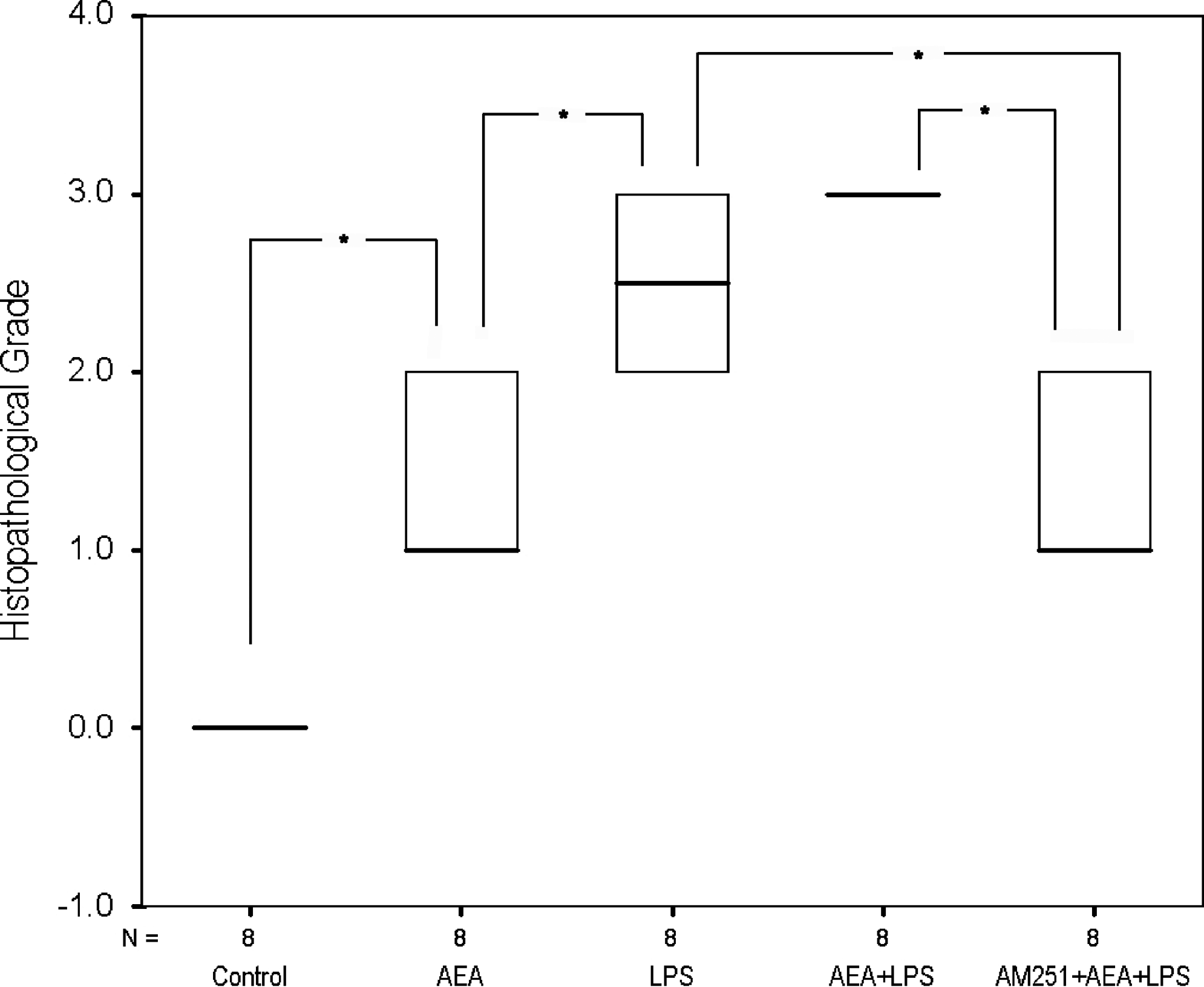
Histopathological examination results as evaluated with Tilton's scale. 23 Data are presented as medians. Kruskal–Wallis variance analysis and Mann–Whitney U tests were performed. *P<0.01 was considered significant with regard to Bonferroni correction.
Discussion
AEA, the endogenous cannabinoid receptor ligand, has a cannabinomimetic action profile.2,18 Cannabinoids have ophthalmologic effects as well, such as decreasing intraocular pressure (IOP) and causing conjunctival hyperemia without pupillary dilatation.11,21,24,25 Topical application of AEA also has ophthalmologic effects. 21 Cannabinoid receptors have been identified in the eye. 26 Additionally, enzymes playing a role in the biosynthesis and inactivation of endocannabinoids were also isolated from ocular structures. Widespread distribution of cannabinoid receptors in the retina and the presence of enzymes responsible for the synthesis and metabolic processes of cannabinoids indicate that endocannabinoids may play an important role as modulator substances in retinal activity.27,28
Selective and highly potent CB1 antagonist AM251 was shown to alter decreased IOP following noladin administration, a putative novel endogenous cannabinoid, in rabbit eyes.29,30 It has also been shown that topical administration of AEA causes conjunctival hyperemia without flare. 21
In the present study, we observed that intravitreal AEA application caused an identifiable inflammation, which had a clinical score varying between 1 and 4. The inflammatory process induced by application of intravitreal AEA may be either due to its conversion to AA and ethanoamine by ocular amidases and/or nonspecific esterases30,31,32 or mediated directly by cannabinoid receptors. Attributable to the administration route of AEA, intravitreal application might have failed to mimic the crucial features of a complex in vivo system, such as corneal penetration limitations, ocular distribution, and metabolism; thus, AEA might have reached supraphysiological concentrations in the ocular tissues. When histopathologically examined, not severe but a noticeable inflammatory reaction, mainly defined by vascular changes and exudative lesions, was observed in the irises, and inflammatory cell infiltration to a mild extent was usually present in anterior and posterior portions of the eyes in the AEA group, which most probably resulted from an increase in vascular permeability of the dilated capillaries, particularly at the top of the ciliary processes.
AEA drastically enhanced LPS' effect on AH protein content. Protein concentrations were significantly higher in the AEA+LPS group in comparison with all other groups, possibly because of the breakdown of blood-ocular barrier (BOB) by AEA's effect, leading to an increase in leakage. Likewise, AEA administration with LPS significantly facilitated the inflammatory effects of this compound and caused a 2-fold augmentation in cell numbers counted in AH, accompanied with an overall increase in clinical and histopathological evaluation results. This potentiation, if not due to the action of AEA as an adjuvant, can be attributed to the vascular effects of AEA. LPS is known to induce the release of various proinflammatory cytokines.33,34 When vascular damage and BOB breakdown occurs, chemotactic mediators allow cells, as well as macromolecules, to leak into the AC, PC, vitreous, and subretinal space. Besides, cytokine and chemokine production recruits more cells and herewith sustains the inflammatory response. Indeed, in the AEA+LPS group, iris and ciliary body vessels exhibited prominent hyperemia and exudative changes in pathological examinations, which supported our thought that AEA exerted its effect mainly through vascular changes35–38 and appears to be a molecule that enhances the immune response regarding its receptor-dependent vasoactive modality, rather than being a proinflammatory substance itself.
In this study, CB1-receptor antagonist AM251, when applied via subtenon route before the administration of AEA and LPS, significantly decreased the protein concentration and cell counts of AH to the levels similar to that of the LPS group, together with an improvement in clinical and histopathological outcome. This finding indicates a prominent role for ocular CB1-receptors to take place in AEA-mediated augmentation of the LPS-induced inflammatory reaction. Although the recovery obtained by AM251 in histopathological grades was significant, it has failed to provide a statistically significant improvement in clinical evaluation scores. Given the fact that clinical evaluation protocol used for the assessment of studied eyes is pretty much based on vascular changes, it can be speculated that AM251's reversing effect on AEA-induced vascular changes is not as strong as its modulator effect on endotoxin-induced immunological processes. Indeed, in a different set of experiments (n=5), AM251 administration prior to LPS injections produced a remarkable inhibition of LPS-induced cell increase in AH, but it did not change the AH protein concentrations to a significant extent (Fig. 9). This finding may also propose an independent role for AM251 in the regulation of cell recruitment driven by LPS challenge. Roche et al. 39 showed that AM251 significantly attenuated the increase in circulating plasma levels of tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 induced by LPS administration to rats in an in vivo study. Similarly, Mnich et al. 40 recently claimed that CE-178253, a selective CB1-receptor antagonist, reduced the concentrations of TNF-α, IL-1β, and IL-6 as measured in rat sera following an LPS challenge. ILs and TNF-α are known to induce the production of chemoattractant factors such as IL-8 and monocyte chemotactic and activating factor, as shown by Kasahara et al. 41 In this perspective, any possible impact of AM251 on LPS-triggered cytokine production might have altered the overall chemotactic and cell infiltration profiles of the studied tissues in our uveitis model. This can also explain the significant observation of AM251's effect, as assessed by histopathological examinations, even on endotoxin-induced inflammatory courses alone, when administered prior to LPS injections (Fig. 10), but it somehow failed to produce a statistically significant impact on clinical evaluation scores when employed prior to LPS alone or AEA+LPS injections. Therefore, our results appertaining to the effects of AM251 should be interpreted in relation to its pure antagonist effect on AEA's action together with any possible interaction of the antagonist with the mentioned cytokines in a manner involving CB1-receptors.
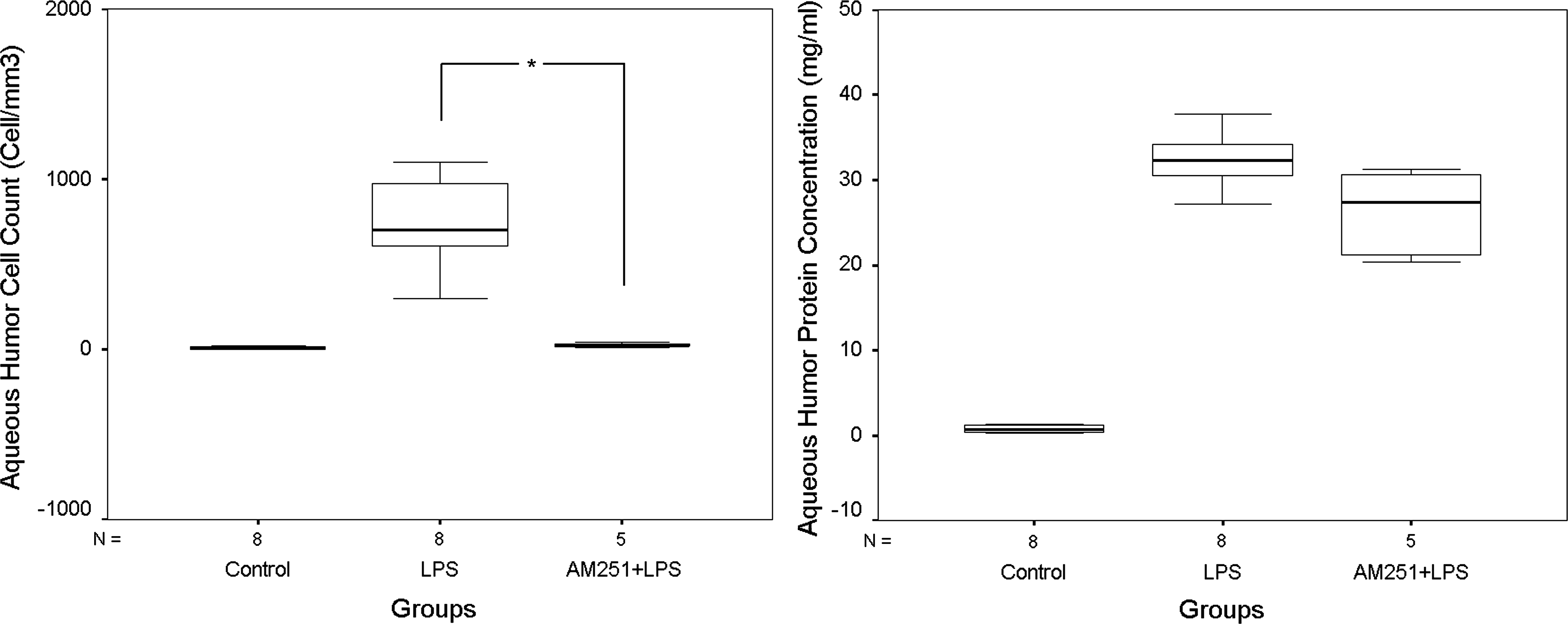
AH leukocyte count (left panel) and protein concentration data (right panel) obtained from eyes treated with AM251 at 30 min prior to LPS injection (AM251+LPS group, n=5). Data are expressed as medians. Kruskal–Wallis variance analysis and Mann–Whitney U post hoc tests were performed. *P<0.016 was considered significant with regard to Bonferroni correction.

Histopathological (left panel) and ocular inflammation scores (right panel) of eyes from the LPS and AM251+LPS groups in comparison with the control group. Data are expressed as medians. Kruskal–Wallis variance analysis and Mann–Whitney U post hoc tests were performed. *P<0.016 was considered significant with regard to Bonferroni correction.
In conclusion, to the best of our knowledge, this is the first in vivo preliminary report indicating the effects of AEA on intraocular inflammation tested with an EUI model in rabbits. AEA obviously exacerbated EIU in rabbit eyes, and CB1-receptor antagonist AM251 has been found beneficial to prevent this aggravation. As AEA is a clinical choice for lowering IOP, concurrent use of CB1-receptor antagonist in glaucoma cases secondary to intraocular inflammation should be subject to further research as a reasonable strategy for avoiding the aggravation of the present inflammatory course.
Footnotes
Author Disclosure Statement
None of the authors has any competing commercial and/or financial association that might raise a conflict of interest in relation to this manuscript.