
Abstract
Abstract
Purpose:
To clarify the mechanism of prostaglandin (PG) analogue-dependent relaxation in ciliary arteries from wild-type (WT) and prostanoid receptor-deficient mice.
Methods:
The intracellular-free calcium concentration ([Ca2+]i) in isolated WT mouse ciliary arteries was measured by fluorescence photometry. Reduction of [Ca2+]i leading to vascular relaxation by PG analogues latanoprost, isopropyl unoprostone, or tafluprost was compared to the maximum increase of [Ca2+]i by 50 mM KCl. The cyclooxygenase inhibitor indomethacin and the NO synthase inhibitor NG-nitro-L-arginine methylester (L-NAME) were added to investigate the involvement of vascular endothelial factors. Moreover, PG analogue-dependent reduction of [Ca2+]i was measured in ciliary artery strips from FP, EP1, EP2, and EP3 receptor-deficient mice.
Results:
The 3 PG analogues reduced K+-dependent increase in [Ca2+]i in a concentration-dependent manner. Indomethacin (10 μM) had little effect. The reductions of [Ca2+]i induced by 10 μM PG analogues were not significantly affected by the treatment with the NO synthase inhibitor L-NAME (10−4 M). The effect of all 3 PG analogues in FP and EP3 receptor-deficient arteries was similar to the effect in WT arteries. Latanoprost significantly enhanced the reduction of [Ca2+]i in ciliary arteries from prostanoid EP1 and EP2 receptor-deficient mice compared to WT mice. Tafluprost had a similar effect in arteries from EP2 receptor-deficient mice.
Conclusions:
PG analogues latanoprost, isopropyl unoprostone, and tafluprost reduced the K+-dependent increase in [Ca2+]i in isolated mouse ciliary arteries. Endothelial-derived factors and FP and EP3 receptors were not involved in the responses. The increased effectiveness of latanoprost and tafluprost in reducing [Ca2+]i in EP1 and EP2 receptor-deficient arteries suggests that the PG analogues may act, at least partially, through nonprostanoid receptor pathways. For glaucoma patients, PG analogues can be selected to reduce the intraocular pressure and increase the ocular blood flow.
Introduction
Prostaglandin (PG) analogues lower IOP and are widely used as ocular hypotensive drugs for the treatment of glaucoma and ocular hypertension. Interestingly, topically applied PG analogues increase the ocular blood flow. Latanoprost increases the optic nerve head blood flow in humans,4,5 and travoprost increases the ocular blood flow in both humans and animals.6,7 In addition, we reported that the PG analogues latanoprost, isopropyl unoprostone, and tafluprost induce concentration-dependent relaxation of isolated rabbit ciliary arteries.8–10 Using laser speckle flowmetry, we reported that tafluprost, latanoprost, and travoprost prevent the endothelin-1 (ET-1)-induced decrease of optic nerve head ocular blood flow in rabbits. 11 At the same time, we used an isometric tension recording system to show that these PG analogues relax the ET-1-induced ciliary artery contraction, 11 and this relaxation is accompanied by a decrease in the concentration of intracellular-free calcium. 10 Therefore, we hypothesized that the increased ocular blood flow to the optic nerve head induced by PG analogues is the result of a decrease in the concentration of intracellular-free calcium that, in turn, leads to relaxation of the ciliary artery smooth muscles. 11 However, the underlying mechanism has not been clarified yet.
Several prostanoid receptors contribute to the IOP-lowering effect of the currently available PG analogues. For example, the prostanoid FP receptor plays a crucial role, 12 and the EP3 receptor plays an additive role in FP receptor-induced IOP reduction. 13 Other PG receptors, such as EP1 and EP2, are not involved. 13 However, the role of these receptors on PG analogue-induced vascular smooth muscle relaxation has not been determined. In this study, we investigated the effects of PG analogues on isolated mouse ciliary arteries by measuring the intracellular Ca2+ concentration ([Ca2+]i), which is a triggering signal for smooth muscle contraction. 10 Next, the involvement of endogenously produced PGs or endothelial-derived NO in the changes of [Ca2+]i were assessed using the cyclooxygenase inhibitor indomethacin and the NO synthase inhibitor NG-nitro-L-arginine methylester (L-NAME). Furthermore, FP receptor- and EP1-3 receptor-deficient mice were used to clarify which prostanoid receptors are involved in the PG-induced [Ca2+]i reduction.
Methods
Animals
All experiments were performed in compliance with the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research and with the approval of the Animal Research Laboratory of Bioscience Education-Research Center of Akita University. FP, EP1, EP2, and EP3 receptor homozygous knockout (FPKO, EP1KO, EP2KO, and EP3KO respectively) mice were provided by Prof. Shuh Narumiya, Kyoto University, and the genotypes were determined by polymerase chain reaction.12,13 C57BL/6 mice were used as the wild-type (WT) control. Mice were bred and housed in clear cages covered with gauze. In all experiments, we used 8- to 21-week-old mice.
Preparations
Mice were killed by inhalation of diethyl ether. The eyes were immediately enucleated, ensuring that a maximum length of optic nerve was removed, and placed in the Krebs solution of the following composition (mM): NaCl 135, KCl 5.0, MgCl2•6H2O 1.0, HEPES 10, glucose 10, and CaCl2•2H2O 2.5. The pH of the solution was adjusted between 7.2 and 7.4. With the aid of a dissecting microscope, the ciliary arteries were carefully isolated from the optic nerve and surrounding connective tissue. The arteries were cut longitudinally to form strips (50–100 μm in width, 1.2 mm in length) that were immediately mounted on silicone film and stretched with pins at both ends. They were incubated at room temperature in the dark for 90 min in a solution composed of 10 μM fura-2 acetoxymethyl ester (Wako Chemical, Osaka, Japan) in the Krebs solution. The solution was aerated with 95% O2 and 5% CO2. 10
Drugs and chemicals
The following drugs and chemicals were used: carbachol hydrochloride, indomethacin, latanoprost (all from Sigma Chemical Co., St. Louis, MO), NG- L-NAME, isopropyl unoprostone (R-Tech Ueno Ltd., Hyogo, Japan), and tafluprost (Santen Pharmaceutica, Osaka, Japan). Latanoprost and tafluprost were in the isopropyl ester form.
Fluorescence photometry
After fura-2 incubation, the strips were rinsed several times with the Krebs solution to remove extracellular fura-2, and then equilibrated in a fresh Krebs solution. The strips were warmed to 37°C. The [Ca2+]i was measured using the Aquacosmos System (Hamamatsu Photonics K.K., Shizuoka, Japan) equipped with a Nikon epifluorescence microscope (TE2000-U; Nikon, Tokyo, Japan) and band-pass filters for wavelengths of 340 and 380 nm. After correction for individual background fluorescence, the ratio of the fluorescence at both excitation wavelengths (F340/F380) was monitored simultaneously to determine the [Ca2+]i. 14 During the experiment, the solution flow rate was 5 mL/min. 11 At this rate, it took one minute for each new solution to reach the mounted arterial strip.
The baseline [Ca2+]i was determined in the Krebs solution and defined as the 0% response. The Krebs solution was then replaced with a high potassium solution. The high potassium ion solution was composed of (mM) NaCl 90, KCl 50, MgCl2•6H2O 1.0, HEPES 10, glucose 10, and CaCl2•2H2O 2.5. The pH of the solution was adjusted between 7.2 and 7.4. We defined the maximum increase of [Ca2+]i response just before addition of PG analogues as 100%. At 4 min after the solution was replaced, latanoprost, isopropyl unoprostone, or tafluprost was added. The change in [Ca2+]i was expressed as the percent against 100%. In experiments applying just 10 μM PG analogues, we defined the [Ca2+]i at 90 s after the PG analogue application as the percent reduction. Afterward, the solution was replaced with the Krebs solution again.
Statistical analysis
Results are expressed as means±standard deviations with n representing the number of preparations studied. We simultaneously examined 3 - 4 preparations from 1 mouse. Differences between groups were determined by the unpaired Student's t-test. All statistical analyses were performed using the Statistical Packages for the Biosciences, 15 and the 2-sided P-value significance level was P<0.05.
Results
The high potassium solution increased [Ca2+]i in the isolated WT mouse ciliary arteries (Fig. 1). Latanoprost (Fig. 1A), unoprostone (Fig. 1B), and tafluprost (Fig. 1C) reduced the [Ca2+]i increase. For latanoprost, the percent reduction achieved at 3 μM, 10 μM, and 30 μM was 21.3%±3.8%, 53.5%±5.3%, and 83.5%±9.7%, respectively (Fig. 2). For isopropyl unoprostone and tafluprost, the percent reductions in [Ca2+]i were similar to those for latanoprost at each concentration (P>0.05).

Latanoprost-, isopropyl unoprostone-, and tafluprost-induced reduction of [Ca2+]i elevated by high potassium solution in isolated wild-type (WT) mouse ciliary artery strips. These representative concentration–response curves showed that

Effect of prostaglandin (PG) analogues on [Ca2+]i elevated by high potassium solution in isolated WT mouse ciliary artery strips. LAT, n=10, isopropyl UNO, n=8, and TAF, n=10 reduced [Ca2+]i elevated by high potassium solution in isolated WT mouse ciliary artery strips in a concentration-dependent manner. There were no significant differences among the 3 PG analogues at any concentration (P>0.05).
To determine the involvement of endogenously produced PGs, we investigated the effect of 10 μM indomethacin, a cyclooxygenase inhibitor, on the concentration-dependent reduction of [Ca2+]i induced by the 3 PG analogues. The reduction of [Ca2+]i by latanoprost at any concentration was not significantly changed by indomethacin (P>0.05 for all, Fig. 3A). Indomethacin did not inhibit the reduction in [Ca2+]i by 3 μM unoprostone, but it did inhibit that by 10 and 30 μM unoprostone (P=0.046 and P=0.031, respectively, Fig. 3B). In contrast to the effect on the other PG analogues, indomethacin increased the reduction in [Ca2+]i by 3 μM tafluprost (P<0.001, Fig. 3C), but not that at the higher doses.

Effect of indomethacin on PG analogue-induced reduction of [Ca2+]i.
Based on the concentration-dependent reduction of [Ca2+]i by the PG analogues (Fig. 2), the 10 μM concentration of each provided about 50% inhibition of the increase in [Ca2+]i elicited by the high potassium solution. Therefore, we chose that concentration for further studies with each of the PG analogues. L-NAME, a NO synthase inhibitor, abolished relaxation of rabbit ciliary artery segments induced by 1 μM carbachol (data not shown). To determine the role of NO in the PG analogue reduction of [Ca2+]i, we added 100 μM L-NAME to the high potassium incubation solution and to the PG analogue solutions. L-NAME itself had no effect on [Ca2+]i (Fig. 4). In the presence of L-NAME, the reduction of [Ca2+]i achieved by 10 μM latanoprost, isopropyl unoprostone, and tafluprost were each about 50% and were unaffected by the presence of L-NAME (P>0.05 each, Fig. 5).
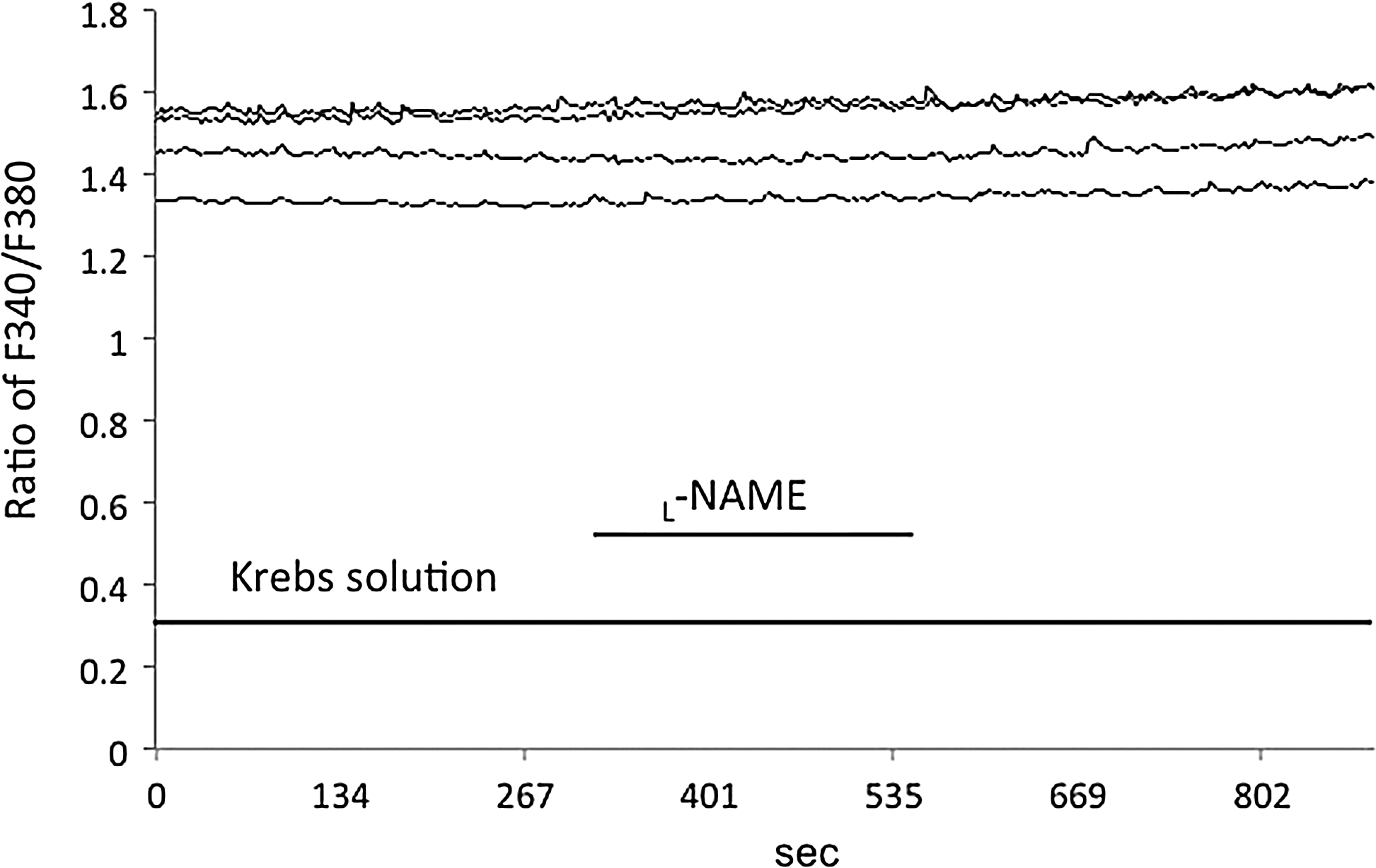
Effect of L-NAME on [Ca2+]i levels. L-NAME (100 μM) in the Krebs solution for 4 min had no effect on [Ca2+]i. Horizontal bar, time in seconds; vertical bars, ratio of F340/F380 that was monitored to determine the [Ca2+]i.
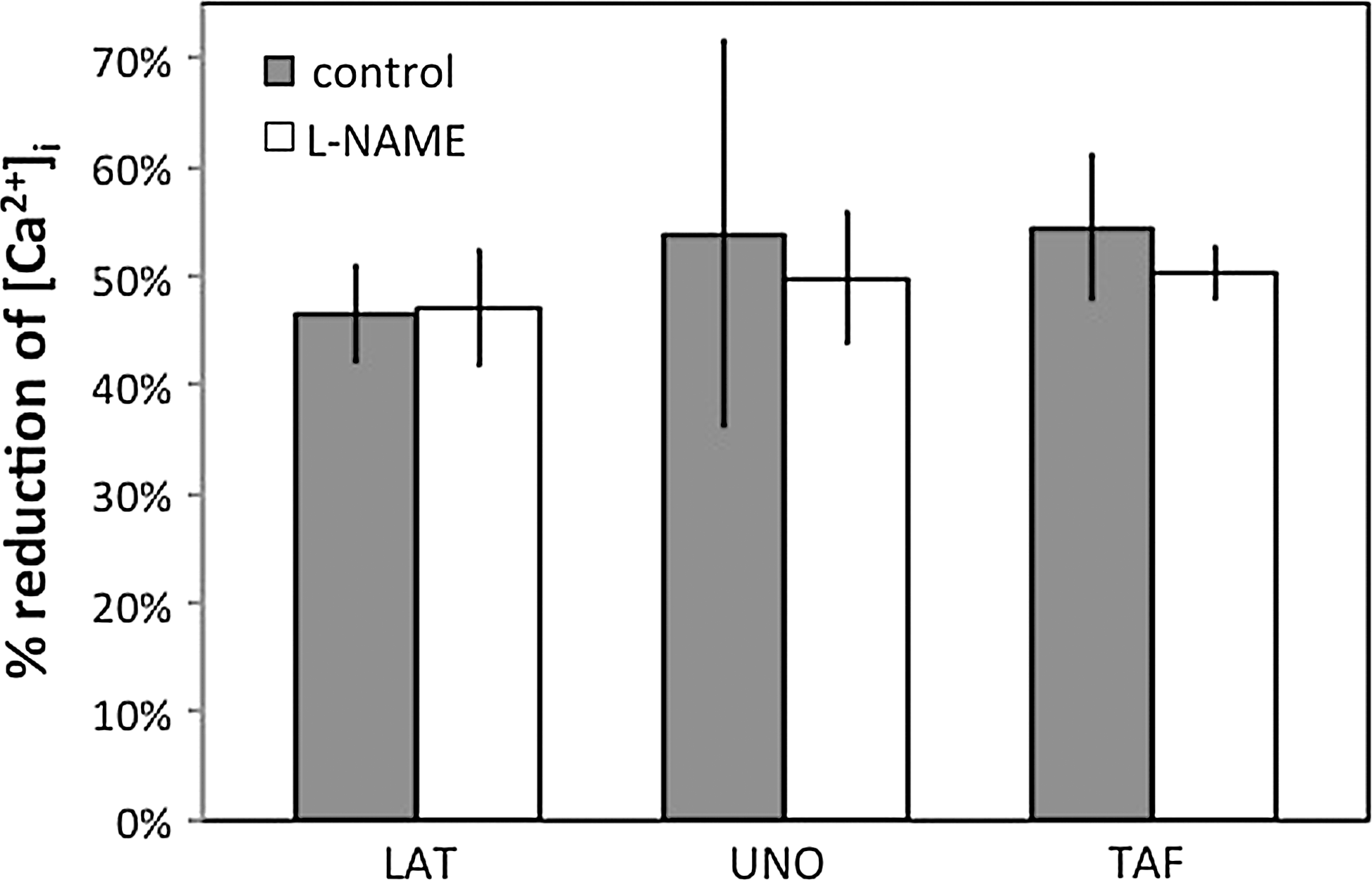
Effect of L-NAME on PG analogue-induced reduction of [Ca2+]i. L-NAME (100 μM) had no significant effect on the decrease of [Ca2+]i caused by 10 μM LAT, n=8, isopropyl UNO, n=8, or TAF, n=8 in WT mouse ciliary arteries (P>0.05 each).
To determine if the mechanism of [Ca2+]i decrease depended on prostanoid receptors, we investigated the effect of the PG analogues in prostanoid receptor-deficient mice (Fig. 6). For ciliary artery strips from EP3KO and FPKO mice, the reductions in [Ca2+]i induced by the 3 PG analogues were similar to those for the arteries from WT mice (P>0.05). The reduction of [Ca2+]i by 10 μM latanoprost in strips derived from EP1KO, EP2KO, EP3KO, and FPKO mice ranged from 41% to 57%. The [Ca2+]i reduction in artery strips from EP1KO and EP2KO mice was significantly more than in WT arteries (P=0.020, P=0.011, respectively, Fig. 6). The reduction achieved by 10 μM isopropyl unoprostone in ciliary artery strips from the knockout mice ranged from 42% to 68%, with no significant differences between the WT and prostanoid receptor-deficient arteries. The [Ca2+]i reduction achieved by 10 μM tafluprost in the preparations from the knockout mice ranged from 49% to 67%. The reduction in arteries from EP2KO mice was significantly more than in arteries from WT mice (P=0.011).

Effect of PG receptor knockout on PG analogue-induced reduction of [Ca2+]i. There were no significant differences in PG-analogue induced reductions of [Ca2+]i in ciliary arteries from EP3KO and FPKO mice compared to those from WT mice (P>0.05). Latanoprost (10 μM, LAT) increased the reduction in [Ca2+]i significantly more in ciliary artery strips from EP1KO and EP2KO mice compared to WT mice. For 10 μM TAF, the increase was significant only for ciliary arteries from EP2KO mice. For 10 μM isopropyl UNO, none of the changes in arteries from the knockout mice were significantly different from the WT arteries. None of the differences induced by isopropyl unoprostone among the WT and knockout arteries were significant (UNO, P>0.05).
Discussion
Previously, we reported that tafluprost, isopropyl unoprostone, and latanoprost, all PGF2α-related compounds used for glaucoma therapy, relax isolated rabbit ciliary arteries in vitro.8–10 The present study is the first report that these 3 PG analogues decrease [Ca2+]i of isolated mouse ciliary arteries in a concentration-dependent manner. The results are consistent with them causing relaxation of ciliary arteries in vivo and increasing the optic nerve head blood flow as shown by laser speckle flowmetry in rabbits.11,16
Our previous study showed that tafluprost ester relaxed precontracted rabbit ciliary arteries in vitro, and the potency was greater than tafluprost acid. 10 Moreover, isopropyl unoprostone relaxed precontracted rabbit ciliary arteries, but the degraded form of unoprostone had no effect on these preparations. 9 It is the acid form of tafluprost and the degraded form of isopropyl unoprostone that are responsible for IOP reduction.17–19 The fact that the relaxation of ciliary arteries was induced by the ester forms, but not the acid forms, 10 suggests that the mechanism for reducing the [Ca2+]i in precontracted ciliary arteries may be different from the mechanism of IOP reduction. Our recent studies also support the notion that [Ca2+]i reduction may be one of the mechanisms of ciliary artery relaxation by isopropyl type PG analogues, but not by acid types. 9 Both tafluprost and isopropyl unoprostone may reach the ciliary artery without any degradation by the loose connective tissue around the eyeball as occurs for nipradilol. 20 Upon arrival at that location, the unaltered drugs could successfully increase the ocular blood flow.
In this study, to reveal the underlying mechanism of [Ca2+]i reduction in response to these 3 PG analogues, we used the cyclooxygenase inhibitor indomethacin to determine if endogenously produced PGs were involved. The indomethacin had no effect on latanoprost-induced [Ca2+]i reductions; however, it inhibited [Ca2+]i reduction induced by isopropyl unoprostone and exaggerated [Ca2+]i reduction induced by tafluprost (Table 1). Because these PG analogues stimulate FP receptors, it seems as though the effect of indomethacin on [Ca2+]i reduction should be similar for all 3 analogues. However, they were not. Thus, the mechanism of [Ca2+]i reduction may be independent of binding to FP receptors, and it may be caused by other activities of each drug. Further studies are required to clarify the mechanisms of [Ca2+]i reduction and ciliary artery relaxation by ester-type PGs.
NS, not significant; ↑, increased reduction. ↓, decreased reduction; LAT, latanoprost; UNO, unoprostone; TAF, tafluprost.
Next, we determined if the endothelial-derived vasodilator NO 21 was involved. The NO synthase inhibitor L-NAME 22 had no effect on PG analogue-induced [Ca2+]i reductions. This suggests that endothelial-derived NO has little to do with the [Ca2+]i reduction induced by the 3 PG analogues.
In mice, the EP2 receptor is highly expressed in the cornea, iridial sphincter muscle and blood vessels, nonpigmented ciliary epithelium, trabecular meshwork, Schlemm's canal area, and all of the retinal layers. 23 The EP1 receptor in mice is predominantly expressed in the ganglion cell and the inner nuclear layers, while the EP3 receptor is rarely expressed. 23 In humans, the distribution of EP receptor subtypes is similar to that of mice. 23 Stimulation of EP2 receptors is thought to cause the relaxation of smooth muscle cells by activating adenylate cyclase, whereas EP1 and EP3 receptors initiate contraction by stimulating intracellular Ca2+ mobilization. 24
In humans, the prostanoid FP receptor is highly expressed in the corneal epithelium, ciliary epithelium, circular portion of the ciliary muscle, and iridial stromal and smooth muscle cells. 25 The FP receptor, 12 but not the EP1 and EP2 receptors, 13 plays a crucial role in the early lowering of IOP in mouse eyes. We found that in the absence of the prostanoid FP, EP1, EP2, or EP3 receptors, the PG analogues latanoprost, isopropyl unoprostone, and tafluprost reduced the high potassium solution-induced [Ca2+]i in isolated strips of mouse ciliary arteries. In FPKO and EP3KO mice, [Ca2+]i reduction by the 3 PG analogues was not significantly different from WT mice. Latanoprost and tafluprost reduced [Ca2+]i in ciliary artery strips from the EP2 receptor-deficient mice significantly more than those from WT mice. Thus, the effects of latanoprost and tafluprost on [Ca2+]i may be partially achieved through the EP2 receptor. Because the acid form of tafluprost has no binding affinity for the EP2 receptor, 26 the involvement of EP2 receptor on the reduction of [Ca2+]i may be caused by an indirect mechanism associated with the PG analogues. Latanoprost reduced [Ca2+]i in ciliary artery strips from the EP1 receptor-deficient mice significantly more than those from WT mice, but the effect was small. These results indicate that the EP2 receptor may be involved in a minor way in the reduction of [Ca2+]i in ciliary artery smooth muscle.
Taken together, our results indicate that PG analogues reduce [Ca2+]i and increase blood flow through a FP receptor-independent mechanism. Nevertheless, the FP receptor is clearly important because it is necessary for PG-mediated IOP reduction. 12 In choosing PG analogues for glaucoma patients, now, we can consider not only the IOP reduction, but also increasing the ocular blood flow. We need further investigation of the precise mechanisms by which the PG analogues act to reduce the IOP and increase the blood flow to the optic nerve. Such studies may lead us to develop new compounds for treating glaucoma.
Upon topical application of 100 μL of 1% nipradilol to monkey eyes, approximately 0.15 μM nipradilol may have penetrated to the periocular tissues around the optic nerve insertion. 20 Similarly, when used clinically, the concentrations of latanoprost, isopropyl unoprostone, and tafluprost at the optic nerve head are likely to be much lower than the micromolar concentrations that we used here. Thus, we need further experiments to determine if the concentration-dependent actions on the isolated ciliary arteries are clinically relevant.
In summary, esters of latanoprost, isopropyl unoprostone, and tafluprost induced concentration-dependent reductions of [Ca2+]i in isolated strips of mouse ciliary artery. The mechanism was not dependent on endothelial-derived factors, such as endogenously produced PGs or NO. Moreover, FP and EP3 receptors responsible for the IOP reduction may not be involved in the mechanism of PG analogue-induced [Ca2+]i reductions, and latanoprost and tafluprost may act, at least partially, through prostanoid receptor-independent pathways.
Footnotes
Author Disclosure Statement
No competing financial interests exist.