
Abstract
Abstract
Purpose:
To compare the inhibitory effects of 4 different types of black currant anthocyanins (BCAs) on ocular elongation in 2 different chick myopia models.
Methods:
In the first model, diffusers were used to induce form vision deprivation. In the second model, negative (−8D) spherical lenses were used to create a defocused retinal image. Either the diffusers or the −8D lenses were placed on the right eyes of 8-day-old chicks for 4 days. Ocular biometric components were measured using an A-scan ultrasound instrument on the third day after application of either the diffusers or −8D lenses. Interocular differences (globe component dimensions of the right diffuser or eyes covered with −8D lenses minus those of the open left eyes) were considered to evaluate the effect of BCAs. The BCAs used were cyanidin-3-glucoside (C3G), cyanidin-3-rutinoside (C3R), delphinidin-3-rutinoside (D3R), and delphinidin-3-glucoside (D3G). Each anthocyanin was administered intravenously at a dose of 0.027 μmol/kg once a day for 3 days.
Results:
Compared to the vehicle treatment, C3G and C3R treatments significantly reduced both differential increases (positive values of interocular differences) of the ocular axial length induced by diffusers or −8D lenses (diffusers; C3G, C3R, and control: 0.32±0.051 mm, P<0.05; 0.25±0.034 mm, P<0.01; and 0.52±0.047 mm, −8D lenses; C3G, C3R, and control: 0.25±0.049 mm, P<0.01; 0.17±0.049 mm, P<0.001; and 0.50±0.056 mm). In contrast, compared to vehicle treatment, D3R treatment significantly decreased the differential increases in the ocular axial length only in chicks with myopia induced by −8D lenses (D3R and control: 0.17±0.049 mm and 0.50±0.056 mm, P<0.001). D3G did not inhibit the differential increase in the ocular axial length induced by either diffusers or −8D lenses.
Conclusions:
This study showed that the 4 tested BCAs had different effects on the 2 different experimental models of myopia.
Introduction

Chemical structures of BCAs. BCAs, black currant anthocyanins.
C3G, C3R, D3G, and D3R have antioxidant activity such as other anthocyanins. 7 C3G and C3R possess inhibitory action on invasiveness of cancer cells. 8 C3G has antiobesity and antidiabetic activities. 3 And C3R has inhibitory action on α-glucosidase. 9
Recently, we reported that oral administration of a BC extract, which contained 25% BCAs, significantly, but not completely, inhibited the ocular growth induced by negative (−8D) lenses in chicks. 10 We also found that intravenous administration of a mixture of the purified BCAs inhibited the ocular growth induced by negative lenses.
Therefore, in the previous study, we confirmed that the BCAs were the active constituents of the BC extract. In that study, the individual effect of each BCA on the −8D lens-induced ocular growth was not determined.
BCAs have different pharmacological actions such as stimulation of the regeneration of rhodopsin 11 by C3G and C3R, inhibition of endothelin-1(ET-1)-induced ciliary muscle contraction 12 by C3R and D3R, and inhibition of monoamine oxidase (MAO)B 13 by C3G, C3R, D3G, and delphinidin. Therefore, it is interesting to evaluate the individual effect of each BCA on experimental myopia.
Both form vision deprivation-induced myopia models and negative lens-induced myopia models are widely used to investigate the mechanism of myopia progression 1 and evaluate the effect of drugs.14,15 Bartmann and Schaeffel reported that translucent occluders, which create form vision deprivation, trigger the increase of ocular growth due to a lack of spatial/temporal transients. 16 On the other hand, negative lens-induced hyperopic defocus seems to be similar to clinical conditions such as an increased accommodation lag and peripheral (off-axis) hyperopia that are linked to the juvenile-onset myopia. 17 It has been reported that disulfiram [an inhibitor of retinoic acid (RA) synthesis] 18 and 6-hydroxydopamine (selectively neurotoxic to dopaminergic cells) 19 inhibited form deprivation myopia (FDM), but not lens-induced myopia (LIM). It has also been reported that the oscillatory potentials measured during electroretinogram (ERG) recordings are reduced only in FDM eyes, even though the other ERG components are the same in both FDM and LIM. 20 These results indicate that FDM and LIM may differ with respect to mechanism.
In the present study, we compared the inhibitory action of 4 BCAs on ocular component elongation induced by either diffusers or negative lenses to determine which anthocyanins were effective at inhibiting ocular elongation in 2 different models of myopia.
Methods
Animals used in these experiments
One-day-old white leghorn chicks were obtained and maintained in temperature-controlled brooders under a 12-h light–12-h dark cycle (switching on of light, 7 AM; switching off of light, 7 PM) with free access to food and water. The temperature of the brooders was set at 32°C on the first day and incrementally decreased to 25°C (1°C decrease per day). For experiments, 7-day-old chicks were transferred to a steel wire cage (2 chicks per cage) in a temperature-controlled (25°C) room with incandescent illumination of about 380 lux at the chick eye level.
These studies were approved by the Animal Committee of the Meiji Seika Food and Health R&D Laboratories. The care and use of the animals were in accordance with the guidelines of the Association for Research in Vision and Ophthalmology (ARVO) statement for the Use of Animals in Ophthalmic and Vision Research.
Application of diffusers and negative lenses
Diffusers were used to create form vision deprivation. Diffusers or negative (−8D) spherical lenses were placed on the right eyes of 8-day-old chicks. Their left eyes (open) served as controls. Diffusers were made from translucent white polypropylene sheets. Diffusers or −8D lenses were attached to the edge of size-matched rings (16 mm outer diameter) by using the cyanoacrylate adhesive for clinical use (Aron Alpha A “Sankyo®”; Daiichi Sankyo Co, Ltd., Nihonbashi, Tokyo, Japan). The rings had a hole with a 14-mm inner diameter and a 5-mm depth. The opposite side of the ring was glued to the circumorbital feathers using cyanoacrylate. Each ring had 4 small holes on its sides for air exchange. In a preliminary study, we confirmed that globe component dimensions between eyes with the ring alone attached and open eyes were not different.
Measurement of globe component dimensions
The anterior chamber depth (ACD, with cornea thickness), lens thickness, the vitreous chamber depth (VCD), and the axial length of the globes were measured by an A-scan ultrasound instrument with a 12-MHz biometry probe (Ophthascan B; Biophysics Medical SA, Clermont-Ferront, France). The measurement was performed under sodium pentobarbital anesthesia [Somnopentyl®, Kyoritsu Seiyaku Corporation, Chiyoda, Tokyo, Japan; 60 mg/kg body weight (BW)] without cycloplegia.
Dosage and administration of BCAs
Commercially available BCAs (D3G•Cl, D3R•Cl, C3G•Cl, and C3R•Cl) were obtained at over 99% purity from the Tokiwa Phytochemical Co., Ltd. (Sakura, Chiba, Japan). D3G•Cl, D3R•Cl, C3G•Cl, and C3R•Cl were individually dissolved in 1 mL 5% glucose solution just before use at a concentration of 0.027 μmol/mL. The BCA concentration was based upon a recent study that demonstrated 0.027 μmol/mL to be an effective dose in reducing negative lens-induced eye growth. 10 BCAs were intravenously administered to chicks via the brachial vein under isoflurane anesthesia at a volume of 1 mL/kg BW. Three minutes later, diffusers or negative lenses were applied. BCAs were then administered once a day for 2 additional days (3 days in total). A 5% glucose solution was similarly intravenously administered to chicks as a control. Globe component dimensions were measured on the third after application of the diffusers or negative lenses.
Statistical analyses of the results
In this study, we considered the interocular differences (globe component dimension of diffuser- or −8D lens-covered right eyes minus those of the open left eyes) to be the difference in globe component dimensions induced by the diffusers/−8D lenses. Positive values of interocular differences indicate differential increases and negative values denote differential decreases. In addition, we used this interocular difference to compare the effect of the BCAs. All results are expressed as the arithmetical mean±standard error of the means (mean±SEM). To compare data between the diffuser- or −8D lens-covered right eye and the open left eye, a paired t-test was used to assess the statistical significance. Between-group measurements for the interocular difference (globe component dimension of diffuser- or −8D lens-covered right eyes minus those of the left open eyes) were evaluated using the Student's t-test or one-way ANOVA with post hoc analysis using the Dunnett's multiple comparison test. P values<0.05 were regarded as statistically significant.
Results
Figure 2 shows the globe component dimensions of vehicle-administered control chicks wearing diffusers or negative lenses. VCD and axial length of diffuser- or −8D lens-covered right eyes were significantly elongated when compared to those of open left eyes (VCD; diffuser-covered eyes and open eyes: 5.50±0.089 mm and 4.89±0.056 mm, P<0.001, −8D lens-covered eyes and open eyes: 5.46±0.098 mm and 4.86±0.047 mm, P<0.001, axial length; diffuser-covered eyes and open eyes: 9.21±0.089 mm and 8.69±0.074 mm, P<0.001, −8D lens-covered eyes and open eyes: 9.19±0.080 mm and 8.69±0.053 mm, P<0.001). However, there were no significant differences in ACD or lens thickness between the open left eyes and diffuser- or −8D lens-covered right eyes. These results were similar to previously reported data indicating that both form deprivation and negative lens-induced hyperopic defocus cause alteration in eye growth that are largely confined to the posterior vitreous chamber. 21
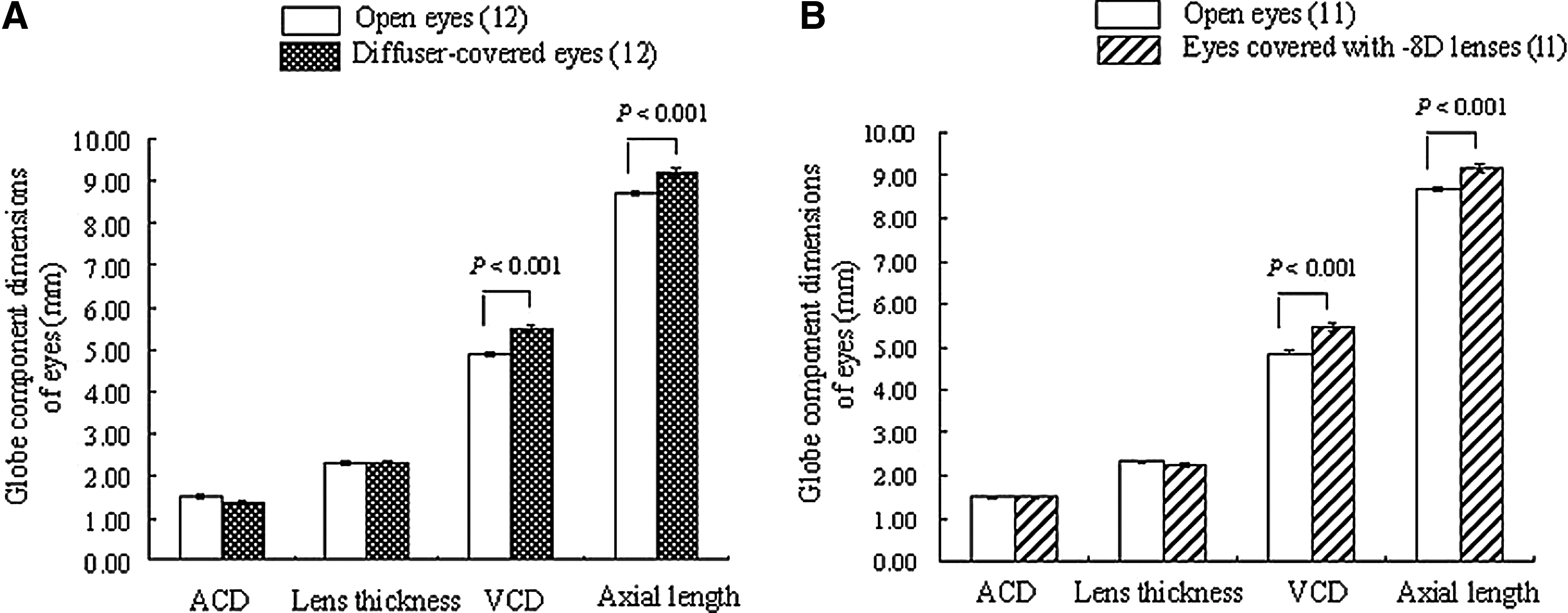
Globe component dimensions of open eyes and diffuser-
The C3G- and C3R-treated chicks displayed significantly reduced differential increases (positive values of interocular differences) in VCD induced by either diffusers or −8D lenses when compared to those of the vehicle-administered control chicks (Fig. 3, diffusers; C3G, C3R, and control: 0.33±0.076 mm, P<0.01, 0.26±0.055 mm, P<0.001 and 0.61±0.068 mm, −8D lenses; C3G, C3R, and control: 0.34±0.067 mm, P<0.05, 0.26±0.076 mm, P<0.01 and 0.60±0.079 mm). In contrast, D3R-treated chicks showed significantly decreased differential increases in VCD only in myopia induced by −8D lenses when compared to vehicle-administered control chicks (Fig. 3, D3R and control: 0.19±0.052 mm and 0.60±0.079 mm, P<0.001). Additionally, D3R-treated chicks showed significantly decreased differential increases in VCD induced by −8D lenses when compared to those induced by diffusers (−8D lenses and diffusers: 0.19±0.052 mm and 0.50±0.037 mm, P<0.001). However, D3G did not inhibit the differential increase of VCD induced by either diffusers or −8D lenses (Fig. 3).
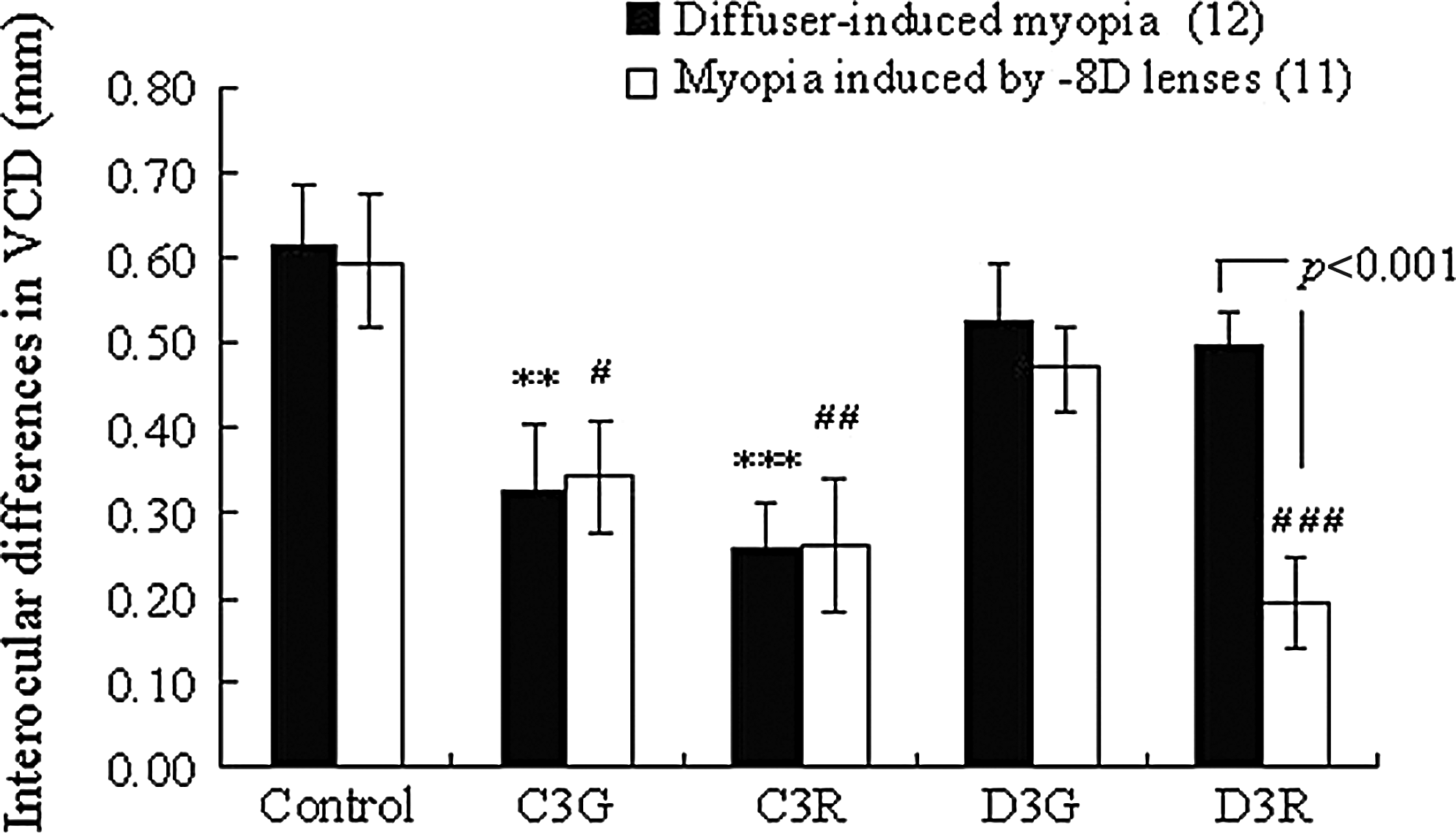
Effects of BCAs on interocular differences of VCD induced by diffusers or −8D lenses. Each column shows the mean±SEM. The numbers in parentheses indicate the number of chicks used. **P<0.01; significantly different from control (diffuser-induced myopia). #P<0.05, ##P<0.01, ###P<0.001; significantly different from control (−8D LIM). P<0.001; significantly different from −8D LIM. P values were calculated by the Student's t-test or the Dunnett's multiple comparison test. LIM, lens-induced myopia; C3G, cyanidin-3-glucoside; C3R, cyanidin-3-rutinoside; D3G, delphinidin-3-glucoside; D3R, delphinidin-3-rutinoside.
The inhibitory effects of C3G, C3R, and D3R on the differential increases in the axial length were of a similar proportion to those on the differential increases of VCD when compared to vehicle-administered control chicks (Fig. 4). Namely, C3G and C3R showed a significant inhibition of the differential increase in the axial length induced by both diffusers and −8D lenses when compared to vehicle-administered controls (Fig. 4, diffusers; C3G, C3R, and control: 0.32±0.051 mm, P<0.05, 0.25±0.034 mm, P<0.01, and 0.52±0.047 mm, −8D lenses; C3G, C3R, and control: 0.25±0.049 mm, P<0.01, 0.17±0.049 mm, P<0.001 and control: 0.50±0.056 mm). In contrast, D3R significantly inhibited only the differential increase in the axial length induced by −8D lenses when compared to those of vehicle-administered controls (Fig. 4, D3R and control: 0.17±0.049 mm and 0.50±0.056 mm, P<0.001). Additionally, D3R-treated chicks displayed significantly decreased differential increases in the axial length induced by −8D lenses when compared to those induced by diffusers (−8D lenses and diffusers: 0.17±0.036 mm and 0.43±0.0043 mm, P<0.001). D3G did not inhibit the differential increase in the axial length induced by either diffusers or −8D lenses (Fig. 4).
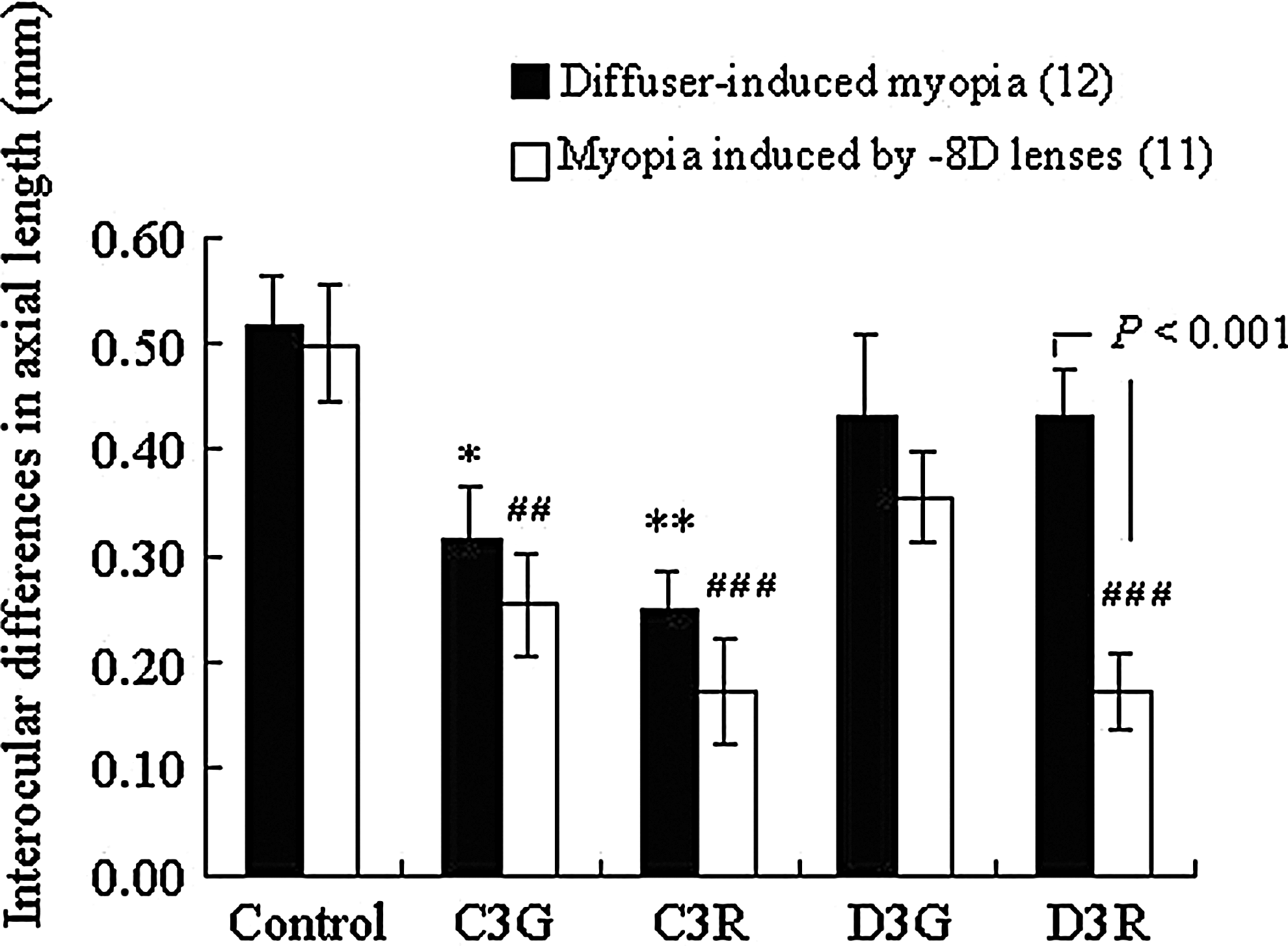
Effects of BCAs on interocular differences in the axial length induced by diffusers or −8D lenses. Each column shows the mean±SEM. The numbers in parenthesis indicate the number of used chicks. *P<0.05, **P<0.01; significantly different from control (diffuser-induced myopia). ##P<0.01, ###P<0.001; significantly different from control (−8D LIM). P<0.001; significantly different from −8D LIM. P values were calculated by the Student's t-test or the Dunnett's multiple comparison test.
C3G, C3R, D3G, and D3R did not influence the intraocular differences in ACD or lens thickness induced by either diffusers or −8D lenses (Table 1). Therefore, we speculate that the inhibitory effects of C3G, C3R, and D3R on the differential increases in the axial length were due to the inhibition of the differential increase in VCD.
Data show the mean±SEM. The numbers in parenthesis were the number of used chicks.
ACD; anterior chamber depth; C3G, cyanidin-3-glucoside; C3R, cyanidin-3-rutinoside; D3G, delphinidin-3-glucoside; D3R, delphinidin-3-rutinoside; SEM, standard error of the mean.
Discussion
In the present study, C3G and C3R inhibited ocular elongation induced by both diffusers and −8D lenses. On the other hand, D3G did not inhibit ocular elongation induced by both diffusers and −8D lenses.
Since, in an in vitro study, C3G, C3R, and D3G all inhibit MAOB, 13 which metabolize retinal dopamine, 22 these anthocyanins can increase the dopamine concentration in the retina by inhibiting MAOB. Because dopamine agonists can inhibit both FDM and negative LIM,23,24 C3G, C3R, and D3G were expected to be able to inhibit ocular elongation induced by both diffusers and −8D lenses via increased dopamine. However, the action of D3G on ocular elongation induced by both diffusers and −8D lenses was different from that of C3G and C3R. Therefore, the inhibitory effect of C3G and C3R on ocular elongation induced by both diffusers and −8D lenses was thought to be due to a process other than inhibition of MAOB.
C3G and C3R could stimulate the regeneration of rhodopsin,11,25 but D3G did not. It is known that the all-trans RA in the retina, choroid, and fundal layer plays an important role for the development of myopia induced by both form deprivation and negative lenses.18,26–28 Because the stimulation of rhodopsin regeneration by C3G and C3R might decrease the synthesis of RA via reduction of the all-trans retinaldehyde during the bleaching process in the visual cycle, 29 we suspect that C3G and C3R were able to inhibit ocular elongation in both myopia models due to the fact that these molecules stimulate the regeneration of rhodopsin (thus attenuating the production of RA)11,25 and inhibit MAOB (increasing dopamine). 13 Although D3G is also a MAOB inhibitor with similar potency to C3G and C3R, 13 it was unable to inhibit the ocular elongation induced by either diffusers or −8D lenses due to its inability to stimulate the regeneration of rhodopsin.
Namely, it was thought that C3G and C3R inhibited ocular elongation due to the stimulation of rhodopsin regeneration. However, it is impossible to exclude the dual action of these compounds on MAOB inhibition and the stimulation of rhodopsin regeneration because C3G and C3R have the effect on both MAOB and rhodopsin regeneration.
Surprisingly, in the present study, D3R inhibited only the ocular elongation induced by −8D lenses, but not the elongation induced by the diffusers. There are currently no reports in the literature about the inhibitory action of D3R on MAOB. Dreiseitel et al. 13 reported on the inhibitory action of delphinidin (an aglycone of D3R) and their data suggest the possibility of D3R inhibition of MAOB.
As D3R, similar to D3G, does not affect the regeneration of rhodopsin, D3R is predicted not to inhibit ocular elongation induced by both diffusers and −8D lenses. However, D3R inhibited only the ocular elongation induced by −8D lenses. D3R dissimilar to D3G is known to inhibit spasm-like contractions of the bovine ciliary muscle induced by ET-1. 12 The observation that D3R inhibits ocular elongation induced by −8D lenses, but not by diffusers led us to speculate that D3R affects accommodation, especially through the ciliary muscle. Nevertheless, accommodation alone may not fully explain the ocular elongation induced by negative lenses.
Wildsoet et al. 30 and Schaeffel et al. 31 reported that in the negative LIM models, the increase of ocular growth and the development of a significant degree of chick myopia are not necessarily related to accommodation, because they observed that ocular elongation and development of myopia were not prevented by the ciliary nerve section or injury of the Edinger–Westphal nucleus. Therefore, the evaluation of the effect of D3R in the negative LIM model, with the ciliary nerve section or injury of the Edinger–Westphal nucleus, would allow us to confirm whether D3R inhibits the increase of ocular growth via the action on ciliary muscle.
In the present study, we confirmed that C3G, C3R, and D3R, similar to the BC extract and a mixture of BCAs, could inhibit the ocular elongation induced by negative lenses.
Therefore, these compounds did not have additive effects from the stimulation of rhodopsin regeneration and accommodative mechanisms.
In chicks, the sclera is composed of an inner cartilaginous and an outer fibrous layer. The experimentally induced ocular elongation is a result of the growth of the cartilaginous layer (measured as increased tissue mass, increased protein synthesis, and an increase of proteoglycan synthesis).32–36
The RA content was increased in the retina and decreased in the choroid during the development of myopia.27,37 Since the sclera, which does not synthesize RA, has the receptors for RA, 38 the choroidal RA affects the growth of the cartilaginous sclera.
Therefore, we speculate that C3G and C3R could increase the RA content in the choroid by the decrease of the RA content in the retina due to the stimulation of rhodopsin regeneration and inhibit the ocular elongation.
Recently, Schaeffel and Matis 39 reported (ARVO abstract 2011) that the BC extract did not inhibit axial elongation or change of refractive development induced by negative (−7D) lenses. We considered the possibility that the discrepancy between our previous study and theirs might be due to differences in experimental protocols such as laboratory illumination, the evaluation period, the number of chicks used per group, or the use of a different chick strain. For example, regarding the observation period, Ashby and Schaeffel 40 reported that bright light inhibited −7D lens-induced refractive development from the second to the fourth treatment day, but not on the fifth day. The results of this study suggest that chicks wearing −7D lenses might have achieved full compensation before the final measurement (on the fifth day) took place. The bright light may inhibit −7D LIM due to the action of dopamine, which is thought to be one of the mechanisms of the BCA activity (inhibitory action on MAOB). 39 In our previous study, we evaluated the effect of BC on −8D lens-induced ocular elongation on the fourth day, rather than on the fifth day. 10 It may be necessary then to evaluate whether BC and each BCA compound can inhibit ocular elongation in experimental myopia over the course of a longer observation period.
In summary, C3G and C3R inhibited ocular elongation induced by both diffusers and negative lenses, but D3R inhibited ocular elongation induced by negative lenses only.
These experiments showed that BCAs had an antimyopia effect not only in a form of mixture, but also in a single form. In addition, the 4 tested BCAs had differential effects on 2 different experimental myopia models.
Footnotes
Author Disclosure Statement
No competing financial interests exist.