
Abstract
Abstract
Purpose:
PF-06653157 is a bifunctional antagonist monoclonal antibody (mAb) that targets human VEGF-A ligand and PDGF-Rβ. With the advent of PF-06653157 as an angiogenesis inhibitor and potential treatment for angiogenesis deregulation diseases, a relevant toxicology species is needed for toxicity and efficacy studies. Investigative studies were conducted to validate the mAb dual antagonist properties in a human system and determine its cross-reactive pharmacology in nonhuman cells.
Methods:
Sequence alignment was used to determine percent sequence identity of VEGF and PDGF receptors and ligands; qualitative reverse transcription polymerase chain reaction (qRT-PCR) was used to determine the presence of PDGF-Rβ on cells of interest. The functional activity of PF-06653157 antibody was assessed in human, dog, porcine, rabbit, rat, mouse, and cynomolgus monkey cells treated with VEGF and PDGF ligands through cell proliferation assays and western blot analysis of AKT and p44/p42 (ERK1/2) protein phosphorylation and enzyme-linked immunosorbent assay.
Results:
PF-06653157 attenuated phosphorylation of AKT and p44/p42 proteins in human and cynomolgus monkey cells. The antibody did not attenuate AKT nor p44/p42 phosphorylation in any other species tested. PDGFR signaling could not be activated with human PDGF ligand in the porcine cells, so PF-06653157 activity in porcine remains inconclusive.
Conclusion:
The PF-06653157 mAb cross-reacts with cynomolgus monkey cells in a similar manner to human cells. Therefore, cynomolgus monkeys are considered the appropriate species for efficacy and regulatory toxicology studies in PF-06653157 development.
Introduction
A
PF-06653157, a bifunctional antagonist, is a stable and efficiently produced monoclonal antibody (mAb), which has been proposed as an angiogenesis inhibitor. 13 PF-06653157 targets human VEGF-A ligand and PDGF receptor β (PDGF-Rβ) in humans 13 and inhibits both VEGF and PDGF signaling cascades. VEGFA is one of the predominant VEGF variants in the angiogenesis cascade and is a critical factor for vascular development 14 ; its isoforms play a role in promoting angiogenesis.14–17 PDGF-Rβ has been shown to be prominently expressed on pericytes and, upon ligand binding, promotes pericyte recruitment needed for microvascular stability and function. 18
With the advent of PF-06653157 as a form of treatment for upregulated angiogenesis in disease, a model species is needed for preclinical toxicology and pharmacology. In this study, we aim to validate the antibody's dual antagonist properties in a human system and determine its cross-reactive pharmacology in nonhuman cells. By determining percent sequence identity of VEGF and PDGF-R in human and nonhuman species, PDGF-R expression across cell lines, and assessing the functional activity of PF-06653157 in different cell models, we identified a cross-reactive species for toxicity and efficacy studies.
Methods
Sequence alignment
Protein and RNA sequences for VEGF-A and PDGF-Rβ from human, pig, mouse, rat, rabbit, dog, and cynomolgus monkey cells were obtained from the NCBI Protein (www.ncbi.nlm.nih.gov/protein) and Nucleotide databank (www.ncbi.nlm.nih.gov/nucleotide) (Table 1). Sequence alignment analysis was conducted using the Clustal Omega online alignment tool for protein and RNA (www.ebi.ac.uk/Tools/msa/clustalo).
Cell culture
Neonatal human dermal fibroblasts (HDFn), pig aortic endothelial cells (POAECs), canine aortic endothelial cells (CnAOECs), and human umbilical vein endothelial cells (HUVECs) were obtained from Cell Applications (San Diego, CA). Dog lung fibroblasts (CF52), fibroblast from Sus scrofa testis at 80–90 days of gestation (Swine testis [ST]), pig turbinate mucosa (PT-K75), rat lung fibroblast (RFL-6), mouse embryo fibroblast (3T3-Swiss Albino CCL-92), and rabbit cornea epithelial (SIRC) cells were obtained from ATCC (Manassas, VA). Cynomolgus dermal fibroblasts (CDFs) were obtained from Primate Biologicals (MD). All cells were grown according to manufacturers' directions. Recombinant human PDGF-BB (platelet-derived growth factor with two B chains) and VEGF ligands were purchased from R&D Systems (#220-BB-010) and Millipore (#GF315), respectively, and murine PDGF-BB and VEGF ligands were purchased from Sigma Aldrich (#SRP3229-10UG, #SRP4364-10UG).
Cell proliferation
Cells were seeded at 2,000 per well in 96-well flat bottom tissue culture-treated white wall plates and allowed to adhere for 6–20 h (allowing for minimal cell division). The growth media were then removed and the cells were serum starved overnight in serum-free media (SFM) consisting of DMEM-F12 media supplemented with 1 × ITS (insulin, transferrin, selenium), 0.1% BSA, 1 mM Na pyruvate, and 2 mM glutamine (Life Technologies). The media were then replaced with SFM containing either VEGF (50 ng/mL) or PDGF-BB (50 ng/mL) ligands, or both, and the cells were treated with PF-06653157 at concentrations ranging from 0 to 10 μg/mL; ligand-free SFM were used for negative control. Following 24, 48, or 72 h of incubation, cell proliferation was assessed using either the BrdU Cell Proliferation Kit (Millipore) or the Cyquant® Proliferation Assay Kit (Life Technologies). One-way analysis of variance with Bonferroni post hoc test and Trend testing were conducted using SAS software. Results were considered statistically significant if P < 0.05.
Gene expression
RNA isolation was conducted using the RNeasy Plus Mini Kit (Qiagen) with freshly thawed cells. Two micrograms of RNA per sample was converted to cDNA with the ABI High-Capacity cDNA Reverse Transcription Kit (#4368813; Life Technologies). The ABI TaqMan Gene Expression Master Mix (#4369016; Life Technologies) and ViiA™ 7 Real-Time PCR System (Thermo Fisher Scientific) were used for real-time qualitative polymerase chain reaction (qPCR) quantification. All procedures were conducted according to the manufacturer's instructions.
The data were normalized to the averaged value of two housekeeping genes, GAPDH and Beta-Actin, and the 2−(ΔCt) method of analysis was used to determine the expression level. Primer probes were obtained from Life Technologies: Human PDGF-Rβ (HS01019589_m1), Human GAPDH (Hs99999905_m1), Human Beta-Actin (HS99999903_m1), Canine PDGF-Rβ (Cf02626572_m1), Canine GAPDH (Cf04419463_g1), Canine Beta-Actin (Cf03023880_g1), Rabbit PDGF-Rβ (AIMSIMO), Rabbit GAPDH (Oc03823402_g1), Rabbit Beta-Actin (Oc03824857_g1), Rat PDGF-Rβ (Rn 00709573_m1), Rat GAPDH (Rn01775763_g1), Rat Beta-Actin (Rn00667869_m1), Mouse PDGF-Rβ (Mm00435546_m1), mouse GAPDH (Mm99999915_g1), Mouse Beta-Actin (Mm00607939_s1), Pig PDGF-Rβ (AJ89KMJ), Pig GAPDH (Ss03374854_g1), Pig Beta-Actin (Ss03376160_u1), Cynomolgus PDGF-Rβ (Mf02842790_mH), Cynomolgus GAPDH (Mf04392546_g1), and Cynomolgus Beta-Actin (Mf04354341_g1).
Immunoblot assay (western analysis)
Cells were seeded, 1 million per well, in 12-well plates. They were allowed to adhere for 6–20 h, after which the growth media were replaced with SFM. After 3–4 h of starvation, varying concentrations of PF-06653157 antibody (0–10 μg/mL) were added to individual wells and incubated at 37°C for 15–20 min. Following incubation with the PF-06653157, either VEGF (50 ng/mL) or PDGF-BB (50 ng/mL) ligands were added into the media and the plates were incubated for another 10 min. The media were aspirated and the individual plates were placed on a bed of dry ice to stop the cell signaling reaction. Protein lysates were prepared using RIPA buffer and supplemented with protease inhibitors (Thermo Scientific™ Pierce™). SDS-PAGE was performed using 10–20 μg of whole cell lysate per sample on a 4%–12% Bis-Tris gel using the Invitrogen XCell II Mini-Lock system at 200 V for 1 h. Transfer to 0.45 μm nitrocellulose membranes was conducted using iBlot® Dry Blotting System (Life Technologies) at 20 V for 7 min.
Following transfer, the membranes were blocked using Rockland Blocking Buffer (#MB-070) and rocked gently at room temperature for 1–3 h. The blocking buffer was removed and the membranes were incubated with primary antibodies from Cell Signaling Technology: phospho-PDGF-Rβ Tyr751 [#4549], PDGF-Rβ [#4564], phospho-VEGF-R2 Tyr1175 [#2478], VEGF-R2 [#2479], phospho-AKT Ser473 [#9271], AKT [#9272], phospho-p44/p42 Thr202/Tyr204 [Cat. No. 9101], and p44/p42 [#9102] at a dilution of 1:1,000 in 1 × Tris-buffered saline with 0.5% Tween 20 (1 × TBST) (Kirkegaard & Perry Laboratories, Inc.). The membranes were rocked gently overnight at 4°C, then washed at room temperature with 1 × TBST in a series of two 10-min washes and one 15-min wash. The blots were then incubated with the secondary antibody, Li-COR anti-rabbit IRDye 800CW (#926-32211), at a dilution of 1:20,000 in 1 × TBST for 1 h in the dark. After incubation, the membranes went through a second series of washes before imaging. The membranes were scanned using the Odyssey® Infrared Imaging Systems (LI-COR). The 800 nm channel was selected for scan; the resolution was set to 169 μm, quality was set to medium, and the focus offset was at 0.0 mm.
Phospho ELISA analysis
Seeding and PF-06653157 treatment of cells for enzyme-linked immunosorbent assay (ELISA) analysis followed the same procedure set in place for the immunoblot assay. The plate-based analysis was performed with the PathScan® Phospho-Akt (Ser473) Sandwich ELISA kit obtained from Cell Signaling (Cat. No. 7160). Phospho-signal was normalized using the PathScan Total Akt ELISA Sandwich ELISA kit (Cat. No. 7170). The assay and data analysis were performed according to the manufacturer's instructions.
Results
Sequence alignment
A multispecies alignment was done using VEGF-A and PDGF-Rβ protein and RNA sequences from human, pig, rabbit, rat, mouse, and dog cells. Only PDGF-Rβ alignment is presented for cynomolgus monkey since its VEGF-A protein sequence is known to be identical to human VEGF-A.19,20 Table 2 displays percent sequence identity of RNA and protein sequences from each species to human VEGF-A and PDGF-Rβ. Pig RNA returned the greatest percent sequence identity to human VEGF-A (96.56%), followed by rabbit (92.92%), dog (91.81%), rat (86.93%), and mouse (86.88%). Similarly, the pig protein sequence returned the highest percent identity with human VEGF-A protein (95.79%), followed by dog (95.33%), rat (90.65%), rabbit (90.49%), and mouse (84.25%). In terms of PDGF-Rβ RNA sequence, dog PDGF-Rβ had a greater sequence identity to human RNA (87.09%), followed by cynomolgus monkey (86.43%), then rabbit (82.23%), mouse (78.49%), rat (78.27%), and pig (75.54%) cells. However, for PDGF-Rβ protein sequence identity, cynomolgus monkey PDGF-Rβ had the highest percent identity to human (98.37%), followed by dog (90.46%), rabbit (89.60%), rat (86.42%), mouse (73.64%), and pig (73.13%) cells.
Sequence alignment of all species tested indicated that the cynomolgus monkey PDGF-Rβ proteins have the highest similarity to human sequences.
Gene expression
Since PF-06653157 binds to PDGF-Rβ, qRT-PCR was conducted to determine receptor expression in our cells of interest. PDGF-Rβ gene expression was evaluated in HDFn, HUVECs, CnAOECs, canine lung fibroblasts (CF52), ST, pig turbinate mucosa (PT-K75), pig aortic endothelial cells (POAECs), rat lung fibroblast (RFL-6), mouse embryo fibroblast (3T3-Swiss Albino), rabbit cornea epithelial (SIRC) cells, and CDFs. Figure 1 displays PDGF-Rβ gene expression for each cell line. PDGF-Rβ expression was similar across the majority of the cell lines with the exception of PAOECs, HUVECs, and ST, which were the lowest expressers. HUVEC expression of PDGF-Rβ was extremely low (<1.5% of other cell lines), and no amount of PDGF-Rβ expression could be detected in ST.
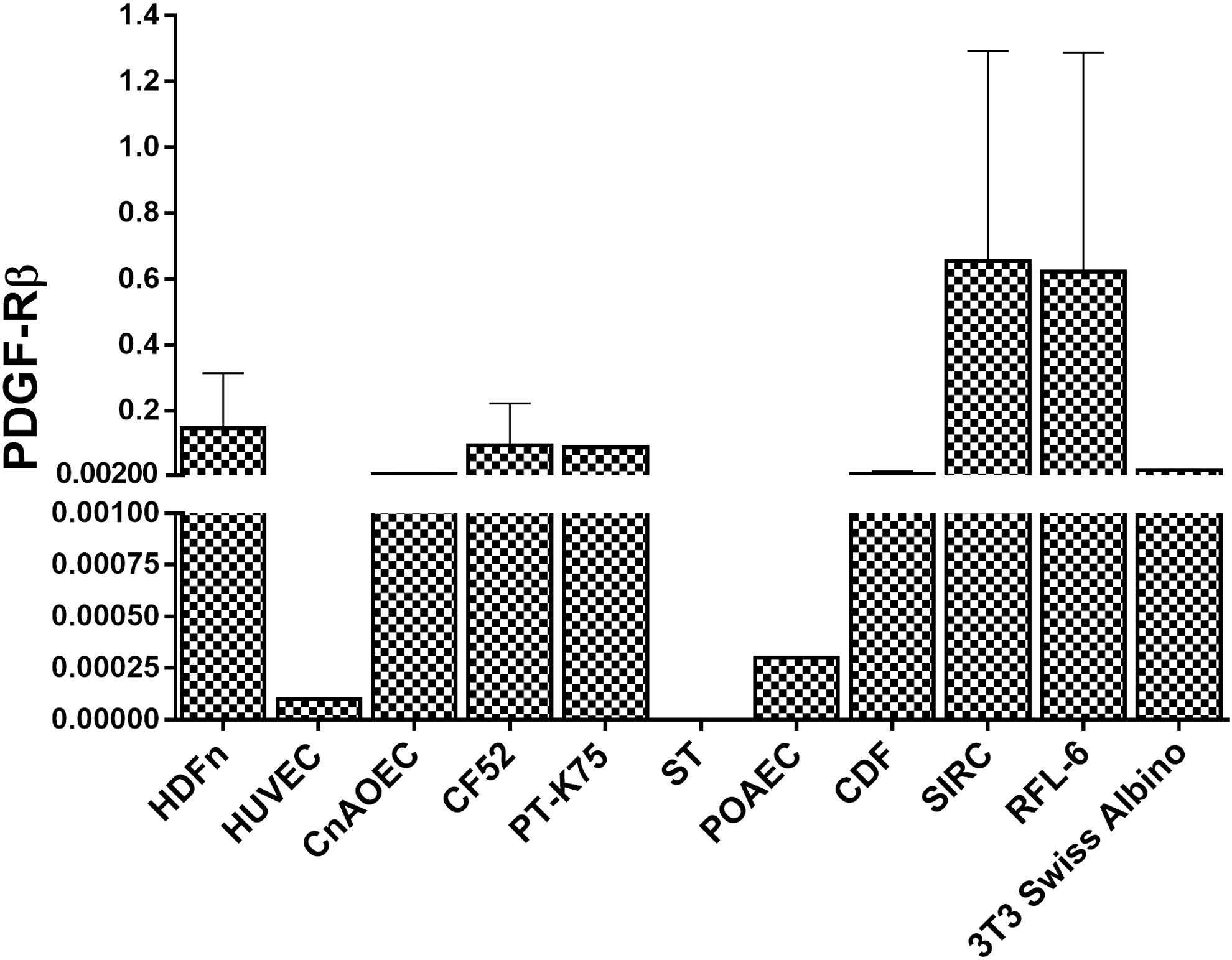
Gene expression of PDGF-Rβ across cell lines (N = 3, data normalized to GAPDH and Beta Actin averaged value). All cell lines, with the exception of pig testis (ST), expressed detectable levels of PDGF-Rβ; however, HUVECs and POAECs appeared to express negligible levels of PDGF-Rβ in comparison with the other cell lines (<4.5%). SIRC and RFL-6 appeared to be the highest expressers; however, when taking into account the standard deviation, their PDGF-Rβ gene expression levels fall in line with the expression levels of the rest of the positive cell lines.
Validation of PF-06653157 functional activity in human cells
The antagonistic property of the PF-06653157 mAb, in terms of its anti-human PDGF-Rβ and human VEGF-A functional activity, was assessed using neonatal HDFn and HUVECs. For immunoblot analysis, the HDFn cells were pretreated with 0, 1, or 5 μg/mL of PF-06653157 and stimulated with VEGF (V), PDGF-BB (P), or V + P ligands for 10 min. An untreated media-only condition was included as a negative control. Immunoblot analysis revealed that PDGF and V + P stimulation of HDFn cells increased the phosphorylation of PDGF-Rβ and the phosphorylation of its downstream kinase, Akt (Fig. 2A). Treatment with PF-06653157 antibody attenuated the PDGF and V + P-induced phosphorylation, indicating that the antibody was able to antagonize PDGF-Rβ function (Fig. 2A). VEGF was unable to induce any phosphorylation signal in HDFn cells since HDFn cells do not express VEGF-R,21,22 therefore V + P treated cells can be considered equivalent to PDGF-treated cells.

Attenuation of PDGF and VEGF-induced signaling in human cells.
The antagonist property of PF-0665317 on human VEGF was assessed using immunoblot analysis of Akt and p44/p42 (Erk1/2) phosphorylation in HUVECs (Fig. 2B); HUVECs are known to express VEGFR and little to no PDGFR.21,22 The cells were pretreated with 0, 1, or 5 μg/mL of PF-06653157 antibody, followed by stimulation with either VEGF or PDGF or no ligand. Immunoblot analysis indicated that VEGF stimulation induced VEGF-R2 phosphorylation and increased phosphorylation of two downstream kinases, Akt and p44/p42 (Fig. 2B). The VEGF-induced phosphorylation was blocked by the PF-06653157 antibody (Fig. 2B). PDGF was unable to induce a response, therefore PDGF-Rβ antagonism was not assessed in HUVECs (Fig. 2B).
To confirm and recapitulate the data observed in the immunoblot experiment, a plate-based phosphorylation assay was conducted. HDFn cells were treated in a similar manner as previously described and phosphorylation of Akt kinase was assessed using the total Akt phospho-ELISA method (Fig. 3A). The results shown are normalized to total Akt protein signal. As indicated in Fig. 3A, PDGF stimulation resulted in a > 0.50-fold increase in the phospho-Akt/Akt ratio compared with the untreated cells. This increase was attenuated with the addition of PF-06653157. The increase of phospho-Akt/Akt ratio in PDGF-stimulated cells as well as the attenuation by PF-06653157 was statistically significant at all doses (P < 0.05). Since V + P-treated HDFn are essentially equivalent to PDGF-treated HDFn, statistical analysis results were similar between the two treatments. Trend testing indicated that PDGF and V + P-treated cells displayed significant dose-related attenuation by PF-06653157 (P < 0.0001).

Attenuation of PDGF induced phosphorylation and proliferation in HDFn cells.
The BrdU incorporation assay, as a measure of cellular proliferation, was conducted to test antagonistic properties of PF-06653157 independent of protein phosphorylation. The cells were stimulated according to the cell proliferation assay protocol and were treated with 0, 0.5, 2, or 8 μg/mL PF-0665317. Figure 3B shows the experimental results of BrdU incorporation assays in HDFn cells at 24 h poststimulation with ligand. PDGF was able to increase BrdU signal > 0.5-fold over untreated cells, indicating an increase in cell proliferation (Fig. 3B). This increase was attenuated by PF-06653157 antibody, particularly at the highest concentration used. The increase in BrdU signal of PDGF-treated HDFn relative to untreated cells was statistically significant (P < 0.05). Attenuation by PF-06653157, compared with ligand only treatment, was also statistically significant at the 2 and 8 μg/mL doses (P < 0.05), but not the 0.5 μg/mL dose (P ≥ 1.0). A similar trend was noted in the V + P-treated cells, but attenuation by PF-06653157 was not statistically significant based on pairwise comparisons (as opposed to ligand only). However, when trend testing was used over all the doses, both PDGF and V + P-treated cells showed significant dose-related attenuation by PF-06653157 (P < 0.01).
Evaluation of PF-06653157 functional activity in nonhuman cells
After establishing experimental assays to examine PF-06653157 antagonism of PDGF and VEGF function in human cells, PF-06653157 effects on nonhuman cells were evaluated.
Dog
Immunoblot analysis of dog aortic endothelial cells and fibroblast (CF52) treated with 0, 0.4, 2, or 10 μg/mL PF-06653157 was conducted (Fig. 4), and phosphorylation of Akt as well as p44/42 was assessed. In dog aortic ECs, human PDGF was able to induce a robust increase in phospho-Akt and phospho-p44/p42 signal compared with control cells (Fig. 4A); this increase was not abrogated by PF-06653157 antibody (Fig. 4A). The CF52 cell line was responsive to human PDGF stimulation as evaluated by an increase in Akt phosphorylation, but the signal increase was not abrogated by the addition of PF-06653157 antibody (Fig. 4B). PF-06653157 did not antagonize PDGF-Rβ signaling in the dog species using two independent cell types, one a tumor cell line (CF52) and the second a primary cell (dog aortic endothelial cells), indicating a lack of functional activity in dog.

PF-06653157 did not attenuate phosphorylation of downstream kinases in canine cells stimulated by human PDGF. Canine CF52 and aortic endothelial cells were pretreated with 0, 2, and 10 μg/mL PF-06653157, then stimulated with human PDGF (nonstimulated cells were used for control).
Pig
Immunoblot analysis of phospho-Akt and phospho-p44/p42 in pig aortic endothelial cells (ECs) and ST was conducted following similar conditions to the dog cells. Human PDGF stimulation was unable to induce an increase in Akt kinase phosphorylation in the ST cell line, and no increase was seen in either Akt or p44/42 phosphorylation in pig ECs (data not shown). Functional activity of PF-06653157 could not be assessed in pig, therefore its porcine cross-reactivity is inconclusive.
Rabbit
PF-06653157 treatment of the rabbit corneal epithelial cell line (SIRC) was also assessed using immunoblot analysis (Fig. 5). The cells were treated with the same concentrations of PF-06653157 as the dog and pig cell lines and phospho-Akt and phospho-p44/p42 protein signal was assessed. As shown in Fig. 5, human PDGF induced a strong increase in both Akt and p44/p42 phosphorylation in SIRC, which PF-06653157 was unable to attenuate. This observation was similar to the results obtained for the dog cell lines and indicates a lack of pharmacological effect in rabbit.

PF-06653157 did not attenuate phosphorylation of AKT and p44/42 in rabbit SIRC cells. SIRC cells were pretreated with 0, 2, and 10 μg/mL PF-06653157, then stimulated with human PDGF (nonstimulated cells were used for control). Human PDGF was able to induce phosphorylation of AKT and p44/42, which were not reduced with PF-06653157 treatment.
Cynomolgus monkey
Cynomolgus monkey was examined using primary dermal fibroblasts isolated from cynomolgus monkey skin tissue. Immunoblot analysis was conducted following the same conditions and treatments as the other nonhuman cells. Phospho-Akt and phospho-p44/p42 protein signal was once again assessed. As shown in Fig. 6, human PDGF-induced Akt phosphorylation in the cynomolgus primary dermal fibroblasts and this effect was attenuated by the PF-06653157 antibody (Fig. 6), demonstrating cross-reactive functional activity in this species. The attenuation in the phosphorylation signal was less pronounced when phospho-p44/p42 was assayed (Fig. 6).
PF-06653157 attenuates phosphorylation of AKT and p44/42 in CDFs. CDFs were pretreated with 0, 2, and 10 μg/mL PF-06653157, then stimulated with human PDGF (nonstimulated cells were used for control). Human PDGF induced phosphorylation of both AKT and p44/42. The increase in AKT phosphorylation by PDGF was abrogated by PF-06653157 at both doses tested. The reduction of P-p44/42 signal (although not as robust as P-AKT) is visible in the upper band of the couplet (P-p44). CDF, cynomolgus dermal fibroblast.
Rat
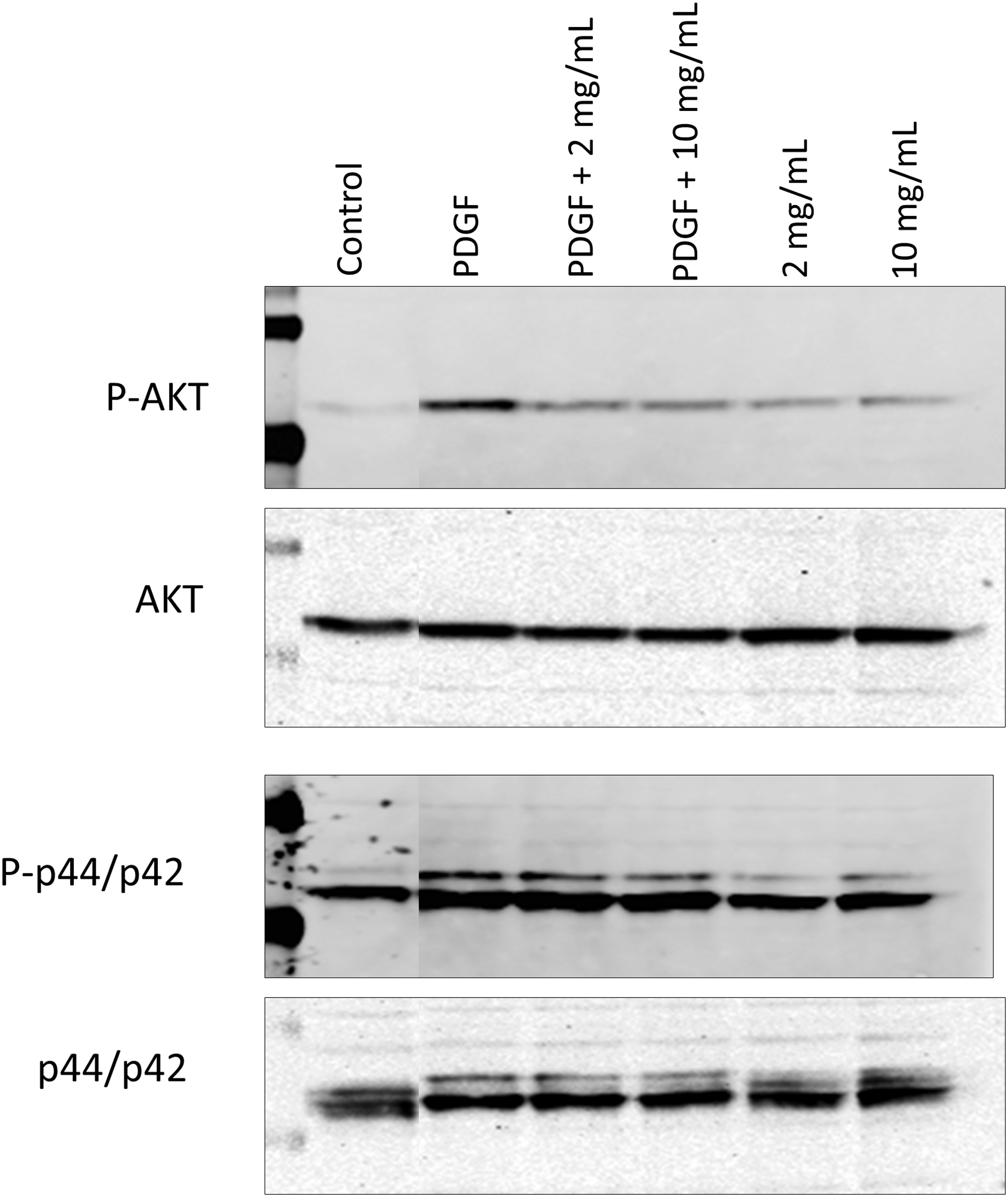
To investigate PF-06653157 activity in the rat species, a cell proliferation assay using the CyQuant DNA dye was performed on RFL6 cells, a fetal lung fibroblast line. The cells were treated with human and murine PDGF and 0, 0.4, 2, or 10 μg/mL PF-06653157. As shown in Fig. 7A, human PDGF was able to induce a small increase (≤ 0.5-fold) in DNA content relative to untreated cells during a 48-h treatment period. This change was not affected by the PF-06653157 antibody. Confirmation with immunoblotting analysis demonstrated that human and murine PDGF did increase phosphop44/p42 signal in RFL6 cells and that Pf-06653157 did not reduce that signal (Fig. 7B), indicating that PF-06653157 is not reactive with the rat fibroblast cell line.

PF-06653157 does not alter PDGF-induced proliferation or kinase phosphorylation in rat fibroblast (RFL6).
Mouse
To investigate PF-06653157 activity in mice, 3T3-Swiss Albino mouse embryo fibroblasts were treated with both human and murine PDGF and the PF-06653157 antibody. The CyQuant DNA dye was used to determine proliferation effects, followed by immunoblot analysis. After 72 h of incubation, the relative DNA content of 3T3 cells was significantly increased by both human and mouse PDGF ligands (P < 0.01) (Fig. 8A). However, there was no attenuation by the PF-06653157 antibody at 0.4, 2, or 10 μg/mL (Fig. 8A). Immunoblot analysis indicated a lack of attenuation of phospho-p44/p42 signaling, confirming that PF-06653157 does not alter phosphorylation p44/p42 in murine cells (Fig. 8B).
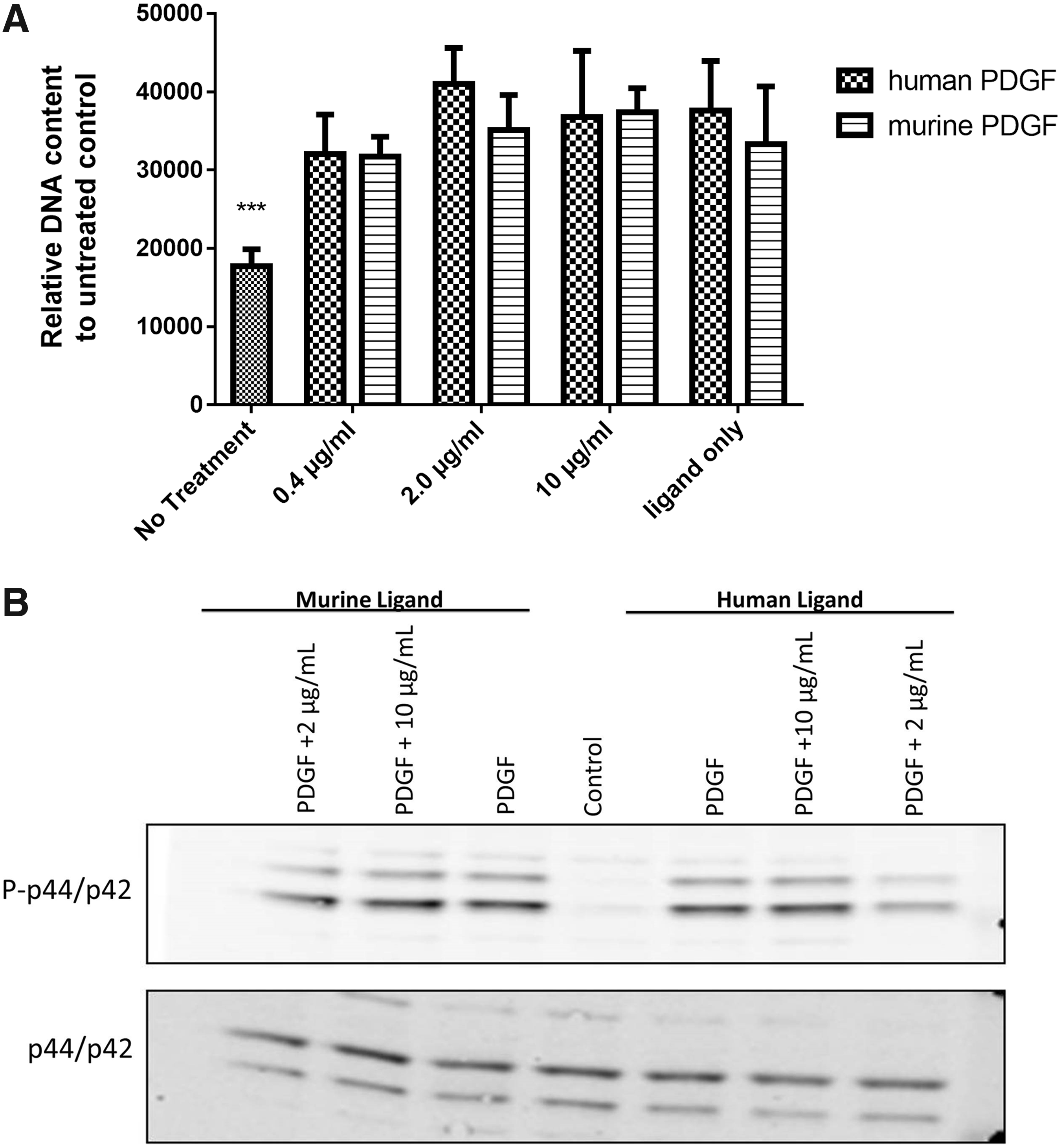
PF-06653157 does not alter PDGF-induced proliferation or kinase phosphorylation in mouse fibroblasts.
Discussion
The purpose of this study was to identify a model species for efficacy and regulatory toxicology studies for PF-06653157 mAb, a PDGFR, and VEGF dual antagonist, proposed as therapy for diseases in which angiogenesis is upregulated. According to the International Conference on Harmonization of Technical Requirements for Registration of Pharmaceuticals for Human Use (ICH) guidelines, selection of a species for toxicology studies should involve assessment of functional activity, which can be assessed in a species-specific cell-based system, and modulation of a biological response or pharmacodynamic marker. 23
The experiments in this study looked at species-specific modulation of kinase phosphorylation in response to PF-06653157 antibody treatment. HDFn and HUVEC types were specifically chosen to display PF-06653157's dual antagonistic properties through one pathway at a time. HDFn lack VEGFR expression21,22 and were used to display PF-06653157's antagonism of the PDGF pathway, while the HUVEC line was a control to confirm PF-06653157's antagonism of the VEGF pathway (since HUVECs express VEGFR, but little to no PDGFR21,22).The antagonistic properties of PF-06653157 were validated in human cells by assessing the reduction in AKT and p44/42 phosphorylation and cell proliferation in VEGF and PDGF-stimulated cells.
AKT and p44/42 are key intracellular mediators of angiogenesis, 24 which are activated through PDGF and VEGF signaling. Binding of VEGF and PDGF ligands to their receptors activates the PI3K/AKT and MAPK pathways,24–26 which leads to the phosphorylation of AKT and p44/42. By binding to VEGF-A ligand and PDGF-Rβ, PF-06653157 should inhibit ligand binding and activation of PI3K/AKT and MAPK pathways and attenuate phosphorylation induced by human VEGF and PDGF. This attenuation was made evident in PDGF-mediated phosphorylation of AKT and p44/42 in HDFn (Fig. 2A) and VEGF-mediated phosphorylation in HUVECs (Fig. 2B); a clear reduction in signal was observed in both immunoblot analysis (Fig. 2) and the total Akt phospho-ELISA (Fig. 3A). A significant dose-dependent decrease (P < 0.05) in cell proliferation was also seen in PDGF-stimulated HDFn cells treated with PF-6653157 (Fig. 3B), which further supports PF-06653157 inhibition of ligand binding, since AKT and p44/42 activation promotes cell proliferation through their downstream pathways.27,28 These results confirm the functional activity of PF-0665317 in human cells and provide a reference for assessment of PF-0665317 functional activity in nonhuman species.
Since the goal of these studies was to determine the primary pharmacological activity across species, to eliminate potential pharmacological or toxicological model species, various cell types were considered. Therefore, convenient in vitro models expressing the appropriate target receptors were employed and included fibroblast, endothelial, and epithelial cells from different organs/tissues. All species' cell types (with exception of pig cells) were confirmed to express PDGFR or VEGFR, or both, using immunoblot analysis, through increase in phosphorylation of AKT and p44/42 in the presence of PDGF and VEGF ligands (Figs. 2, 4–6, 7B, and 8B).
The functional activity studies in nonhuman species suggest that PF-06653157 mAb cross-reacts with cynomolgus monkey cells in a manner similar to human cells. Sequence alignment of PDGF-Rβ protein from cynomolgus monkey shared the highest percent identity to human PDGF-Rβ (98.37%), when compared with dog, pig, rabbit, and rodents (Table 2), and VEGF-A protein sequences of cynomolgus monkey and human cells are known to be identical.19,20 Since comparison of percent sequence identity between nonhuman and human PDGF-Rβ and VEGF-A is predictive of PF-06653157's potential cross-reactivity, it follows that the greater the percent identity between protein sequences, the greater the chance of PF-06653157 mAb binding its target epitope in nonhuman species. This finding proved true in that cynomolgus monkey was the most relevant species with respect to displaying attenuation of protein phosphorylation by PF-06653157.
Immunoblot analysis revealed that human PDGF induced AKT and p44/42 phosphorylation in the cynomolgus primary dermal fibroblast and that its effect was attenuated by the PF-06653157 antibody (Fig. 6). Inhibition of AKT phosphorylation was clearly evident at all doses of the antibody, but attenuation of the phosphorylation signal was less pronounced when phospho-p44/p42 was assayed (Fig. 6), although band intensity measurements did support a dose-dependent decrease in signal (data not shown). This suggests the possible presence of additional PDGF-R isoforms on dermal fibroblasts and/or secondary signals induced by PDGF stimulation that feed into p44/p42 activation in these particular cells.29,30 Two types of PDGF receptors have been identified in literature, PDGF-Rα and PDGF-Rβ; PDGF-Rβ binds to PDGF-BB with high affinity, whereas PDGF-Rα can bind to all three homodimeric isoforms of PDGF. 29
VEGF stimulation in cynomolgus monkey was not assessed since human and monkey protein sequences are identical.
Pharmacology studies in other nonhuman species did not indicate any cross-reactivity of PF-06653157. Although human PDGF did cross-react with canine and rodent species, PF-06653157 mAb did not elicit a pharmacological response in any of the canine or rodent cells tested. Porcine activity was inconclusive given the lack of pathway activation with nonporcine-specific reagents and the commercial unavailability of porcine-specific reagents. Therefore, based on the functional activity of PF-06653157 in these experiments, the cynomolgus monkey would be the species of choice for efficacy and regulatory toxicology studies.
Footnotes
Acknowledgments
The authors would like to thank Dingzhou Li and Brad Hirakawa for their statistical support and quality control (QC) review, respectively.
Author Disclosure Statement
No competing financial interests exist.