
Abstract
Abstract
Purpose:
The ocular half-life of intravitreally (IVT) injected drugs is of major relevance for the suitability of a drug intended for chronic intraocular treatment, as the half-life determines the dosing frequency. Thus, half-life extension principles are very attractive as they can reduce the IVT dosing frequency. In this study, we investigated the ocular pharmacokinetics (PK) of the IVT injected Nanobody® BI-X and whether the noncovalent binding of BI-X to vitreous albumin could increase its ocular half-life.
Methods:
Wistar rats were dosed IVT with 3 μg BI-X alone or coadministered with human serum albumin (HSA), and the ocular exposure was measured in a pilot experiment using whole eye homogenates. New Zealand White rabbits received IVT injections of 500 μg BI-X alone or coadministration with HSA. Concentrations of BI-X were determined in aqueous humor, vitreous body and plasma and pharmacokinetic parameters were calculated.
Results:
Ocular concentrations of BI-X in rats were about 10- or 3-fold higher at 24 and 72 h, respectively, when dosed with HSA. In rabbits, coadministration with albumin led to an about 3-fold increased vitreous half-life and an about 5-fold higher exposure in vitreous humor.
Conclusion:
As small amounts of albumin are present in the vitreous body of healthy human eyes and the albumin concentration is even increased under disease conditions like diabetic retinopathy, high affinity binding to albumin may be a promising strategy to extend the half-life of IVT injected drugs, allowing for longer dosing intervals.
Introduction
I
The molecular weight (MW) of a compound is one important factor for its ocular half-life after intravitreal injection as shown by del Amo and Urtti 4 in rabbits. It became obvious that small molecules with a MW below 1,000 Da have ocular half-lives usually far below 1 day and thus are generally not useful for a chronic dosing without special formulation efforts such as sustained release systems or ocular implants. By contrast, macromolecules with MW larger than 50 kDa like fusion proteins, antibodies, antibody fragments, or aptamers have intrinsic ocular t1/2 of greater than 1 day. Thus, all current IVT antineovascular treatments used in the clinics are macromolecules (ranibizumab, pegaptanib, aflibercept, and bevacizumab with MW of 48, 50, 100, and 149 kDa, respectively) with determined or predicted human ocular t1/2 between ca. 5 and 11 days.5–8
For subcutaneous or intravenous treatment with macromolecules such as peptides, for example, liraglutide or Nanobodies, half-life extension principles have been shown to prolong the residence time of a drug candidate in the systemic circulation. One principle is high affinity binding to albumin. For example, a bivalent Nanobody® (Nb) recognizing the IL-6 receptor and albumin showed after intravenous administration a serum half-life of 6.6 days, which is similar to the expected half-life of serum albumin in this species. 9 In addition, peptides like liraglutide showed extended half-lives after subcutaneous administration due to high-affinity binding to albumin. 10
While the total concentration of soluble proteins in the vitreous body is low, the relative concentration of albumin is high as more than 50% of the vitreal soluble protein in humans is albumin.11,12 The total vitreal protein concentration in healthy humans was reported as 820 or 770 mg/L.11,12 In addition, the albumin concentration in human vitreous was found to be 293 mg/L in healthy humans and elevated in patients with diabetic, traumatic, and idiopathic proliferative retinopathy to 536, 743, and 2,215 mg/L. 12 Shitama et al. reported that the fraction of albumin remained constant with about 60% of total vitreal protein in different diseases and total protein increased, for example, up to 15.75 mg/mL (15,750 mg/L) in proliferative vitreoretinopathy. Thus, elevated albumin concentrations can be expected under most treatment conditions. With a human vitreal volume of 4 mL, a MW of 66 kDa, and the reported concentration of 293 mg/L, a total amount of approximately 18 nmoles vitreal albumin can be estimated for healthy individuals.
BI-X is a trispecific 40 kDa Nanobody, which has one high affinity binding site to human albumin (KD = 1.4 nM) and 2 target binding sites, that is, anti-VEGF and anti-Ang2. Nanobodies® are a novel class of proprietary therapeutic proteins based on single-domain antibody fragments that contain the unique structural and functional properties of naturally occurring heavy chain only antibodies. Ablynx NV is the worldwide holder of the trademark Nanobody. Antagonism of Ang2 together with VEGF may have the potential to be superior in contrast to the currently available sole anti-VEGF treatments of neovascularization in diabetic rethinopathy. 13 A dose of 0.5–1 mg of BI-X would translate into 12–24 nmoles. Thus, equimolar albumin binding could be possible. In diseased eyes, even an excess of albumin can be expected.
The purpose of our studies was to characterize the ocular PK of BI-X. To investigate if binding of BI-X to vitreal albumin and thus increase of the apparent MW of a BI-X-albumin complex could extend its ocular half-life after IVT injection, we compared the ocular PK of BI-X alone and coadministered as a premix with an excess of human serum albumin (HSA) in a pilot study in rats. As rabbits are a more suitable species for human PK prediction of intraocular drugs in comparison to rats,4,14 the effect of albumin binding on the ocular half-life of BI-X was also assessed in a rabbit study. Importantly, BI-X shows no binding to rat or rabbit albumin. Thus, no binding to endogenous vitreal rat or rabbit albumin is considered when BI-X was dosed alone into rat and rabbit eyes.
Methods
Rat studies
All procedures were conducted according to the German Animal Welfare Act, were approved by the local authorities, and followed the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research.
In total, 18 male Wistar rats (RjHan:WI, Janvier Labs) were used in 2 studies. Animal weights were ca. 225–250 g. Animals were housed in groups of 3 according to the subgroups for sampling at different time points in standard Macrolon® cages (type IV) on LIGNOCEL® FS 14 (Altromin) bedding during the study. The temperature was held at 20–24°C and the relative humidity at 45–65%. Animals were kept in a 12-h light: 12-h dark cycle (from 6:00 a.m. to 6:00 p.m.). Food (standard rat diet no. 3438, Provimi Kliba AG) and water were available ad libitum.
The compound was dissolved in 25 mM succinate buffer, 155 mM trehalose, 5% HP-β-CD, and 0.03% (w/v) polysorbate 20 at pH 6.0 at a concentration of 10 mg/mL. One group received BI-X only. For that purpose, the stock solution was diluted 1:10 (v/v) with the buffer resulting in a final concentration of 1 mg/mL (24.6 μM) BI-X. The second group received a premixed solution of BI-X with HSA, containing a 10-fold molar excess of HSA (Sigma-Aldrich, lyophilized, fatty acid free, product no. A3782). For that purpose, 5.02 mg HSA was dissolved in 270 μL of the same buffer and then 30 μL of the stock solution (10 mg/mL BI-X) was added. This resulted in a final concentration of 253 μM HSA and 24.6 μM BI-X.
For the intravitreal administration, rats received one drop of Novesine (0.4% oxybuprocaine) per eye for topical anesthesia. Then, animals were anesthetized with isoflurane, and the injection of 3 μg BI-X in a volume of 3 μL was performed ca. 1 mm posterior to the limbus using a 10-μL syringe (Nanofil, WPI) with 35G microneedles (WPI, Nanofil NF35BV-2).
Rats received the first injection into the right eye, and immediately before terminal sampling, a second injection was administered into the left eye. Thus, the 0-h time point was obtained from each animal, whereas 3 animals per sampling time point (24, 48, and 96 h) were used. The whole eyes were sampled, transferred into preweighed FastPrep® vials (MP Biomedicals LLC), and stored frozen until quantification of BI-X.
Rabbit studies
The rabbits were treated according to the Directive 2010/63/UE European Convention for the Protection of Vertebrate Animals used for Experimental and Other Scientific Purposes and to the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research.
In total, 24 male pigmented rabbits (strain HY79b, Hypharm, Roussay, France) were included in 2 separate studies. The animals weighed 2.0–2.5 kg and were 2–3 months old. Animals were housed individually in standard cages. The temperature was held at 15–21°C and the relative humidity at 55% ± 10%. Animals were routinely exposed (in-cage) to a 10–200 lux light in a 12-h light (from 7:00 a.m. to 7:00 p.m.) and darkness controlled cycle. Animals had controlled enrichment (lucerne; SERLAB). Throughout the study, animals had free access to food and water. They were fed a standard dry pellet diet, LASQCdiet® Rab-14H (LASvendi) distributed every day. Tap water was available ad libitum from plastic bottles. The rabbit studies were conducted at IRIS Pharma, France.
The compound was dissolved at a concentration of 10 mg/mL in 25 mM succinate buffer, 155 mM trehalose, 5% HP-β-CD, and 0.03% (w/v) polysorbate 20 at pH 6.0. For coadministration with HSA, 16.6 mg/mL HSA (Sigma-Aldrich, lyophilized, fatty acid free, product no. A3782) was added and premixed. The solution was filtered (0.22 μm, Millex-GV PVDF Durapore) freshly before administration.
Animals were anesthetized by an intramuscular injection of xylazine and ketamine. A topical anesthesia was induced with 1 drop per eye of Cebesine® (0.4% oxybuprocaine). After cleaning each eye with iodinated povidone, the intravitreal injection was performed, using a contact lens (Landers Vitrectomy lens), under an operating microscope on eyes dilated with one drop of 10% phenylephrine and one drop of 0.5% tropicamide applied 15–20 min before injection. Test item (50 μL) was injected into the midvitreous of the right eye using a 30-G needle mounted on a tuberculin syringe. One drop of Gel-Larmes was topically applied on the eye to avoid ocular dryness as animals recovered.
Approximately 2.5 mL of whole blood was sampled from the central artery of the ear using the EDTA K3 tubes at the respective sampling time points until the respective individual time point of sacrifice (1, 2, 4, 8, 12, 24, 72, and 168 h p.a. or 0, 2, 4, 8, 24, 48, 168, 336, and 672 h p.a. BI-X or BI-X + HSA, respectively, sampling until the respective individual time point of sacrifice). Plasma was prepared by centrifugation for 10 min at 2,000 g at 4°C and stored at −80°C ± 7°C until shipment on dry ice to the Sponsor.
At the time points corresponding to the collection of ocular tissues for the right eyes (12, 24, 72, 168 h p.a. or 24, 168, 336, 672 h p.a. BI-X or BI-X + HSA, respectively), the test item was injected intravitreally (50 μL) into the left eye as described above (0 h). The anesthetized animals were then euthanized by an intracardiac injection of overdosed pentobarbital. Immediately after euthanasia, aqueous humor was collected and the vitreous body was sampled, weighed, and stored in dry ice before being stored at −80°C ± 15°C.
Quantification of free BI-X in ocular and plasma samples
Rat eyes were homogenized in 400 μL homogenization buffer (RIPA buffer, Sigma-Aldrich, diluted 1:4 in PBS with 0.05% Tween) using the FastPrep® homogenizer (MP Biomedicals LLC). Supernatants of homogenized rat eyes and rabbit vitreous samples were diluted in PBS with 0.05% Tween, 0.1% casein, and 0.5 mg/mL HSA (minimum dilution for rat eye 1:5, for rabbit vitreous 1:50). All other samples were further diluted in PBS with 0.05% Tween and 0.1% casein to reach the assay range. To avoid matrix effects, the minimum dilution for rabbit plasma was 1:4.
Free BI-X was quantified by a sandwich ELISA using recombinant target proteins. After coating of recombinant human VEGF (0.5 μg/mL; R&D Systems) and blocking with 1% casein, plates were incubated with the analyte followed by flag-tagged human Angiopoietin-2 (0.2 μg/mL; R&D Systems), an anti-flag antibody coupled to horse-radish peroxidase (0.2 μg/mL; Sigma), and the color reagent tetramethylbenzidine. Quality control samples with a defined concentration spiked into the same matrix and with the same dilution steps were included on each assay plate, and the plate was only considered valid when the deviation to the theoretical value was <20%. Standard curves (100–0.15 ng/mL) for rat eyes and rabbit vitreous were prepared in the presence of 0.5 mg/mL HSA.
Affinity determination
Affinity of BI-X to albumin was determined using surface plasmon resonance. Albumin from rat (Sigma, No. A4538), rabbit (Sigma, No. A0764), or human serum (Red Cross) was coated to different flow cells of the same CM5 chip (Biacore Life Sciences, GE Healthcare Europe GmbH). Sensorgrams were recorded in single cycle kinetics. As control, a goat anti-mouse albumin antibody (Bethyl Laboratories, Inc.; Nr. A90-134A) was used that bound to rat, rabbit, and human albumin (KD(rat) = 2.1 nM; KD(rabbit) = 3.5 nM; KD(human) = 209 nM). BI-X bound to human albumin with a KD = 1.4 nM, but no specific binding to rat or rabbit albumin was observed.
Pharmacokinetic evaluation
The noncompartmental pharmacokinetic evaluation was performed using ToxKin 4.1.0.1 (Entimo AG). Data evaluation used a composite design for all ocular data and also plasma. Therefore, no individual PK parameters could be calculated. Values below the limit of quantification (BLQ) were set to zero.
Results
Quantification of BI-X in ocular and plasma samples
For quantification of BI-X in ocular tissues and plasma, a ligand-bridging ELISA detecting the free Nanobody BI-X was developed using the target VEGF for capture and flag-tagged ANG2 for detection. To test matrix tolerance, serial dilutions of matrix in ELISA buffer (PBS with 0.05% Tween and 0.1% casein) were prepared. Defined concentrations of the analyte were spiked into these dilutions, and the measured values were compared to the theoretical values. Matrix tolerance was considered acceptable when the deviation was below 20%. The following minimum dilutions were determined: rat eye 1:4, rabbit vitreous 1:10, rabbit aqueous 1:1, rabbit plasma 1:4.
Presence of HSA led to a decrease in the ELISA signal. Saturation was reached at around 0.05 mg/mL HSA, so that higher HSA concentrations did not further decrease the signal. Therefore, quantification in rat eyes and rabbit vitreous was performed in the presence of 0.5 mg/mL HSA, thus ensuring an excess of HSA. Quality control samples confirmed the accuracy of this method. Accuracy was 80–120%, and intra-assay precision was <20%. Lower limits of quantification in the study were for rat eye 0.1 nM, rabbit vitreous 123 nM (with HSA) or 0.2 nM (without HSA), rabbit aqueous 0.5 nM, and rabbit plasma 0.1 nM.
Rat ocular exposure after IVT administration of BI-X with and without HSA
The concentrations of BI-X in whole eye homogenate after IVT injection into rats alone or as premix with HSA are listed in Table 1 and depicted in Fig. 1. As expected, immediately after injection, the Nb concentrations were similar between both groups. However, the Nb concentrations at 24 and 72 h were about 10- or 3-fold higher in the HSA than at the same time in the control group, respectively. At the latest time point (96 h), the concentration differences in both groups were small. No pharmacokinetic evaluation was performed as the study was intended to detect mechanistic differences, and therefore, only a minimum number of animals were included.
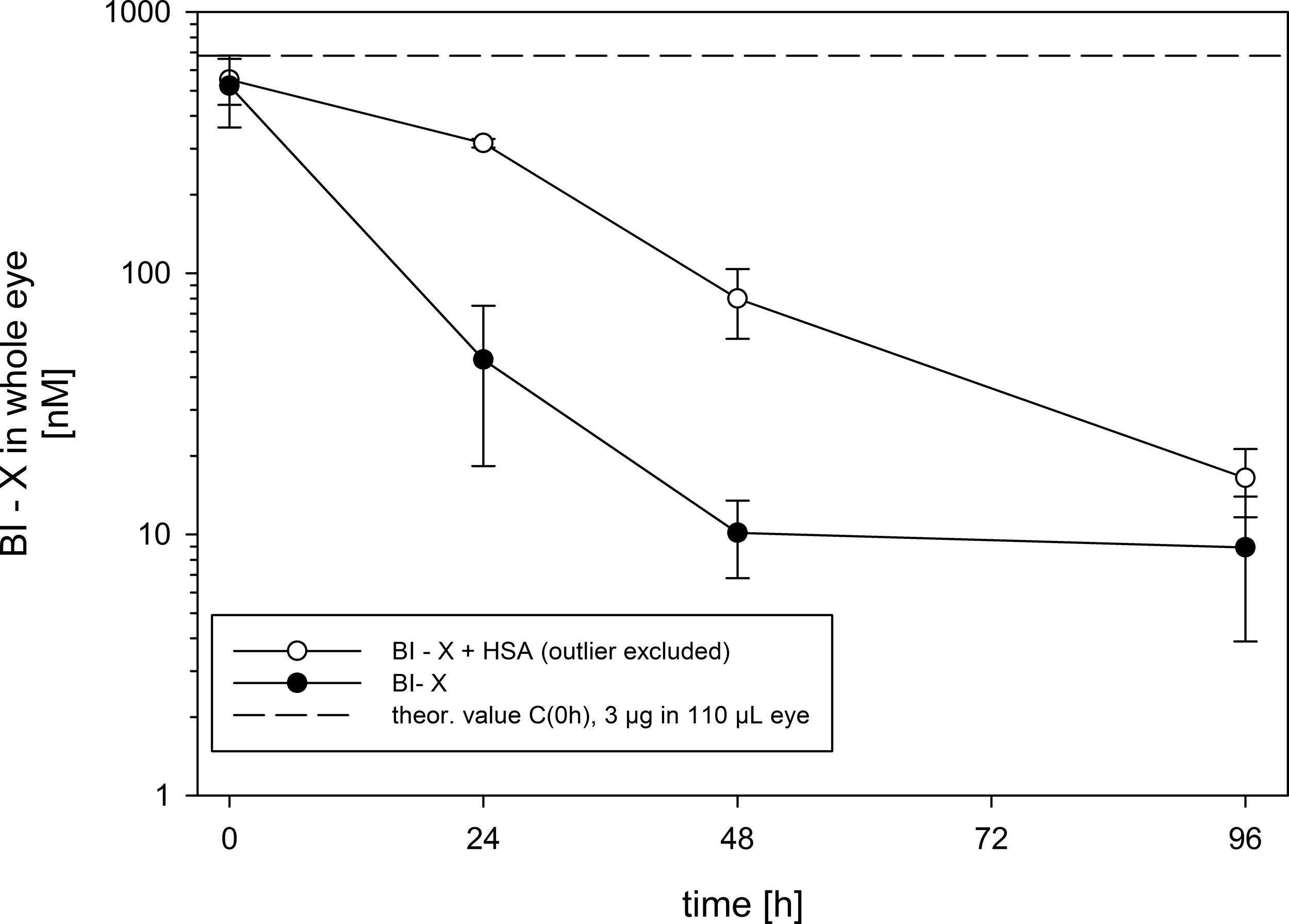
BI-X concentrations in homogenates of rat eyes after a single intravitreal injection of 3 μg (0.07 nmol) in 3 μL per eye either alone or as premix with HSA. HSA, human serum albumin.
Immediately after injection.
HSA, human serum albumin.
Rabbit ocular PK after IVT administration of BI-X with and without HSA
The concentrations of BI-X in vitreous body, aqueous humor, and plasma after IVT injection to rabbits alone or as premix with HSA are depicted in Fig. 2. The pharmacokinetic parameters are summarized in Table 2.
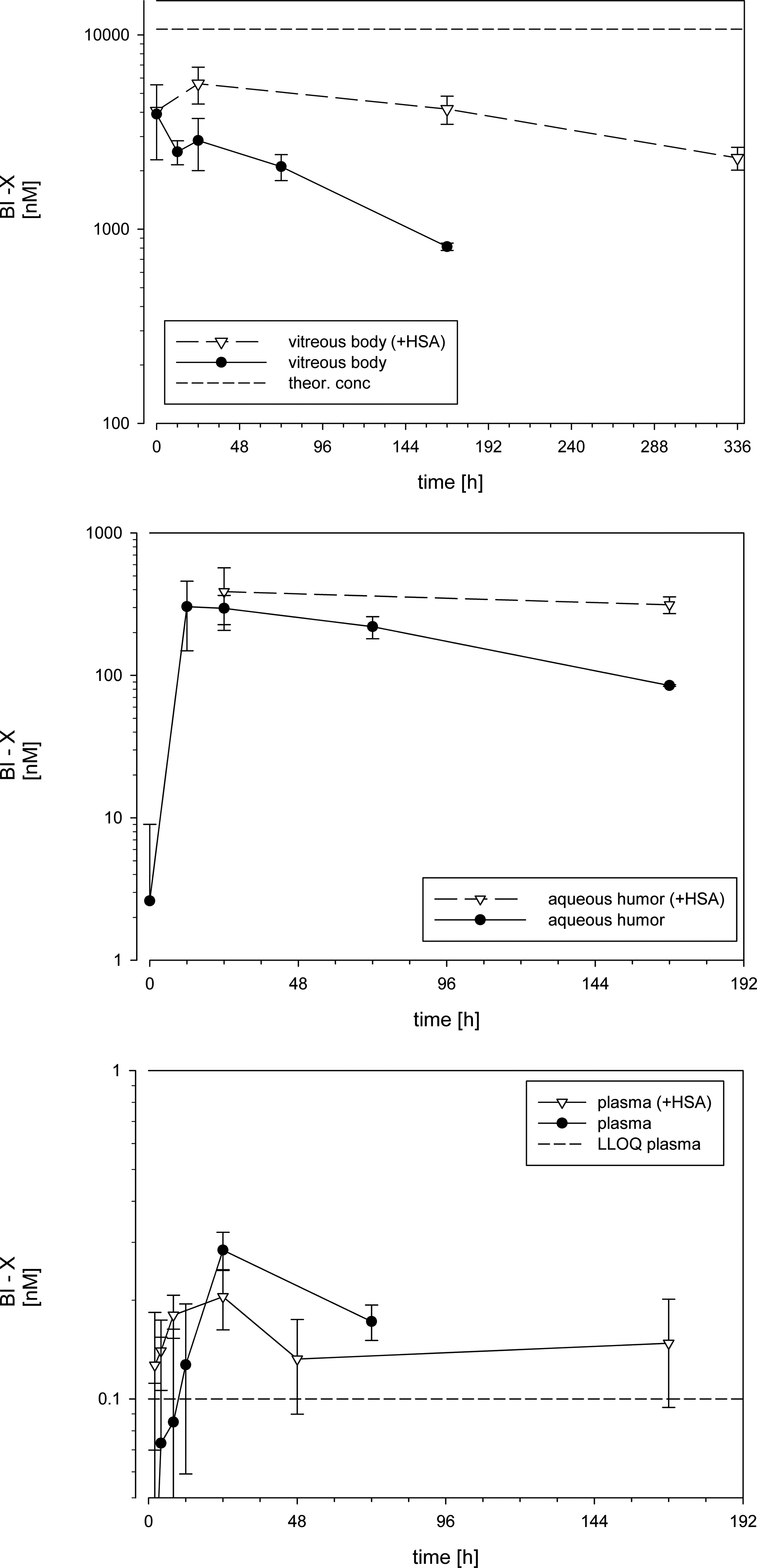
BI-X concentrations in vitreous body, aqueous humor, and plasma of rabbits after a single intravitreal injection of 500 μg (12 nmol) in 50 μL per eye either alone or as premix with HSA. Data are given based on composite data as mean values and error bars reflect SD. Plasma levels BLQ were set to zero. BLQ, below the limit of quantification.
n.a., not applicable; n.c., not calculated.
The mean ± SD of vitreal concentrations of BI-X immediately after injection was similar in both groups (4,660 ± 1,010 and 4,150 ± 2,320 nM in the control and HSA group, respectively). BI-X concentrations declined slowly with time in both dose groups but with different slope. In the BI-X + HSA group, a substantial drop in vitreous exposure was evident between 336 h (2,330 ± 313 nM) and 672 h (BLQ). The exposure to BI-X in vitreous humor was 415 μM·h when dosed alone and 2190 μM·h after coadministration with HSA. The PK parameter derived from the concentration time profiles differed between the groups: when dosed alone, BI-X showed a vitreal t1/2 of 3.2 days (77.9 h), whereas when BI-X was dosed with HSA, the t1/2 increased 3-fold to 10 days (243.5 h). The calculated vitreal CL for BI-X was 0.5 μL/min in the BI-X group but 5.6-fold lower (0.09 μL/min) in the BI-X + HSA group. The volume of distribution varied only slightly between both groups with 3.4 mL in the BI-X group and 2 mL in the BI-X + HSA group, respectively.
The total exposure to BI-X in aqueous humor was about 10-fold lower than in vitreous humor and about 6-fold higher when dosed together with HSA (AUC0-inf = 41.2 μM·h and 268 μM·h, BI-X alone and with HSA, respectively). The respective t1/2 values in aqueous humor were 3.3 and 19.6 days (28.4 and 470 h).
Discussion
The purpose of our study was to investigate the effect of intraocular binding of the Nanobody BI-X to albumin and to characterize the ocular PK of BI-X, which is intended for intravitreal injection. Binding to ocular albumin may have the potential for half-life extension, as already described for systemically administered compounds like peptides for example, liraglutide, 10 or Nanobodies, for example, IL-6-receptor. 9 In our studies, we measured free nanobody through a sandwich ELISA using both target binding sites (VEGF and Ang-2) for capturing and detection. The assay quantified free Nb, regardless if bound to albumin or not.
The vitreous body contains only relatively low concentrations of soluble proteins. However, albumin is the protein with the highest concentration as more than 50% of the vitreal soluble protein consists of albumin in humans11,12 and in diseased eyes even an excess of albumin can be expected. Thus, equimolar binding of 0.5 mg of BI-X to endogenous ocular albumin at physiological concentrations is theoretically possible.
The principles of ocular pharmacokinetics after intravitreal injection have been described by Urtti, del Amo, and Maurice.4,14,15 In general, disposition PK is characterized by the CL (clearance, i.e. elimination) and V (volume of distribution). Ocular elimination mainly occurs through 2 pathways, that is, posterior elimination requiring permeability across the inner and outer blood retina barrier (BRB) and anterior elimination through the aqueous humor. For large molecules, the permeability across the BRB is negligible, and therefore, the anterior clearance becomes the determining elimination pathway out of the eye. Distribution within the vitreous body is mainly driven by diffusion, 15 but convective processes are also possible.16,17 At steady state, and in the absence of pronounced distribution into other ocular tissues like the retina, the ocular volume of distribution of an IVT injected compound is equal to the physiological volume of the vitreous body. In fact, most of the PK data summarized by del Amo and Urtti show a volume of distribution in the range of the physiological vitreal volumes in rabbits and also in humans. As mentioned, the MW of a compound is one important determining factor for its ocular half-life. As a consequence, an increase of the apparent MW by high affinity binding of BI-X to albumin may have the potential for an increase in ocular half-life.
To test the hypothesis, a mechanistic pilot study in rats was conducted. For this purpose, BI-X was premixed with a 10-fold molar excess of human albumin and ocular exposure was compared to the exposure after the same intravitreal dose of BI-X alone in rats. The albumin binding moiety of the Nb is species specific, and high affinity binding was shown for human albumin, but no specific binding was observed to rat or rabbit albumin. Thus, although rat albumin is present also in the rat vitreous body even at higher concentrations than in rabbits and humans, 18 BI-X does not bind to it. The results clearly supported the hypothesis of half-life extension by albumin binding as marked time-dependent exposure differences were observed between the control group (BI-X alone) and the HSA group (Table 1 and Fig. 1) in rats. This clearly demonstrates that the residence time in the rat eye is prolonged when the compound is coinjected with human albumin. At the latest time point (96 h), the concentration differences in both groups were small, but still about 2-fold higher concentrations were found in the HSA group. This may be explained by some high affinity low capacity binding to ocular tissues, as the compound was measured in whole eye homogenates.
To characterize the effect of albumin binding on ocular PK of BI-X quantitatively, we used the rabbit as an animal model. While using a 10-fold excess of albumin in the mechanistic rat study, equimolar amounts of HSA and BI-X were used in the rabbit study to mimic physiological conditions. However, it has to be noted that injection of a premixed Nb and HSA may be different to injection of the Nb into the eye with endogenous vitreal albumin.
As shown in Fig. 2 and Table 2, the vitreal concentrations of BI-X immediately after injection were similar in both groups. These concentrations were, however, lower than expected based on the administered dose and the typical rabbit vitreal volume reported with 1.154 or 1.5–1.8 mL. 19 Ideally, assuming instant homogenous distribution into the vitreous and/or quantitative sampling of all vitreal material, the concentration would be about 11 μM. During sampling of the vitreous body, care was taken not to aspirate other ocular tissues. This may have resulted in an incomplete recovery of vitreal material. Thus, the mean sample weight of vitreous body in this study was slightly lower than the reported anatomical volumes with 0.982 ± 0.182 g and 0.946 ± 0.257 g in the control and HSA group, respectively. For example, Dickmann et al.20,21 noticed that it takes up to 48 h until homogenous distribution of a 50 μL IVT bolus of fluorescent labeled ranibizumab in rabbit vitreous body occurs. Ranibizumab has a MW of 48 kDa, which is similar to the MW of our Nb (41 kDa). Similar results were obtained by computer simulation of FITC-dextran concentration within the vitreous body after IVT bolus injection. 17 Thus, inhomogeneous distribution of BI-X at early time points together with incomplete sampling of the vitreous body may explain the deviation of the Cmax values from the theoretical values. This may also explain the slight increase in the mean vitreal concentrations of BI-X in the HSA group between 0 and 48 h. After 48 h, homogenous distribution can be expected, and under homogenous distribution, even aliquots of the vitreous are considered representative for vitreal concentrations of a compound. Thus, the vitreal half-lives are determined when homogenous distribution can be assumed and are therefore regarded as reliable.
When dosed alone, BI-X shows a t1/2 of 3.2 days (77.9 h). This is in line with ranibizumab, which has a similar MW and vitreal t1/2 in rabbits (2.89 days at a dose of 625 μg 22 ). The calculated vitreal CL for BI-X was 0.5 μL/min, which is in line with the reported data of del Amo and Urtti from similar macromeolecules. 14 When BI-X was dosed with HSA, the t1/2 increased 3-fold to 10 days (243.5 h) and the CL decreased 5.6-fold to 0.09 μL/min. In contrast, the volume of distribution changed only slightly between both groups and decreased from 3.4 mL to 2 mL. Both values are close to the anatomical vitreal volumes and in very good agreement to the reported values of other macromolecules. 14 The only slight differences in volume compared to the substantial differences in CL indicate that the differences in the t1/2 are mainly driven by a decreased clearance rather than an altered distribution into intraocular tissue compartments outside the vitreous body.
Unexpectedly, BI-X concentrations were BLQ at 672 h, whereas at 336 h, a mean BI-X concentration of 2,330 nM was measured. Even assuming the shorter half-life of BI-X without HSA from 336 h onward, the concentrations at 672 h would be still far above the lower quantification limit of 0.2 nM. Thus, the drastic decline of BI-X concentrations after 336 h may rather be explained by formation of anti-drug antibodies (ADAs). It has been described that even after intravitreal injection, biological molecules can elicit an ADA formation, for example, ADAs have been found in the eye after intravitreal injection of Lucentis. 23 ADAs usually lead to highly accelerated clearance of the drug from systemic circulation; however, this effect is not investigated in ocular PK. In addition, binding of ADAs to the drug can interfere with the quantification by ELISA due to competition at the binding sites or steric hindrance. However, we did not further pursue this question as it was out of scope of this study and extrapolation of the influence of ocular ADA formation from rabbits to humans is not feasible.
As already mentioned, compounds diffuse from the vitreal body into the anterior chamber and are eliminated through bulk flow of aqueous humor (AH) out of the eye. In larger animals like rabbits, AH can be easily sampled without any contamination from other ocular compartments by paracentesis, that is, puncture of the anterior chamber, and ocular PK data can be derived from AH. The AH flow rate determines the posterior CL, and AH concentrations at steady state can be expected to be lower than in vitreous humor. This is in line with our data, where the exposure to BI-X in aqueous humor concentrations is about 10-fold lower than vitreal concentrations and the vitreal and AH concentrations decline in parallel within one dose group. Similar data have been reported for ranibizumab 24 and bevacizumab 21 and also for radiolabeled albumin in rabbit eyes. 25
Like in the vitreous body, dosing of BI-X with HSA increased also the exposure to BI-X in the aqueous humor. Determination of the half-lives, and therefore, calculation of AUC0-inf was hampered by the number of data points where BI-X was quantifiable. As the decrease of BI-X concentrations in the different ocular compartments declined with the same rate within one treatment group, it is obvious that the prolongation in vitreal half-life is also reflected in aqueous humor t1/2.
Finally, slight differences were also observed in the plasma exposure of BI-X. However, concentrations in plasma were exceptionally low and most of the early and late time points were BLQ. The most pronounced difference in BI-X plasma levels between both groups were observed at 168 h. At this time point, BI-X was BLQ in all samples of the control group but quantifiable in all samples of the HSA group resulting in a mean value of 0.148 nM. This clearly reflects the prolonged ocular half-life. However, plasma exposure is only about 0.007% of the vitreal exposure.
Taken together, coadministration of BI-X and HSA led to an about 3-fold prolongation of the vitreal half-life of BI-X and an about 5-fold increase of ocular exposure to BI-X in the rabbit. Figure 3 contains the vitreal t1/2 of BI-X alone and with HSA plotted together with vitreal half-lives of macromolecules after IVT injection to rabbits collected by del Amo and Urtti 14 and Gadkar et al. 26 In addition, the t1/2 of rabbit albumin as determined by Maurice is included. This demonstrates that an apparent MW increase increases the ocular half-life and the increase is more pronounced for BI-X (with and without albumin) compared to other molecules between 40 and 150 kDa. In particular, Gadkar et al. 26 reported that doubling the MW from a single Fab (50 kDa) to a Fab2 (100 kDa) only slightly increased the MW. However, the hydrodynamic radius of a compound is the main compound-specific parameter for its diffusion properties. Thus, a doubling of the MW may have very different consequences on the hydrodynamic radius depending on its “shape.” In addition, the net charge also was shown to affect the diffusion rate in vitreous. 27 Finally, it was reported that purified human and bovine serum albumin tends to form oligomers after reconstitution from lyophilized powders. 28 Thus, dimers, tetramers, and hexamers are found, and only 50% of the HSA was present as monomers immediately after dissolution. Binding of BI-X to albumin is highly specific and 1:1 binding is assumed. Thus, it is not clear if the binding site is still accessible in those oligomers, but a BI-X oligomer complex would have a markedly higher MW than a BI-X HSA monomer complex. This was not investigated in more details.
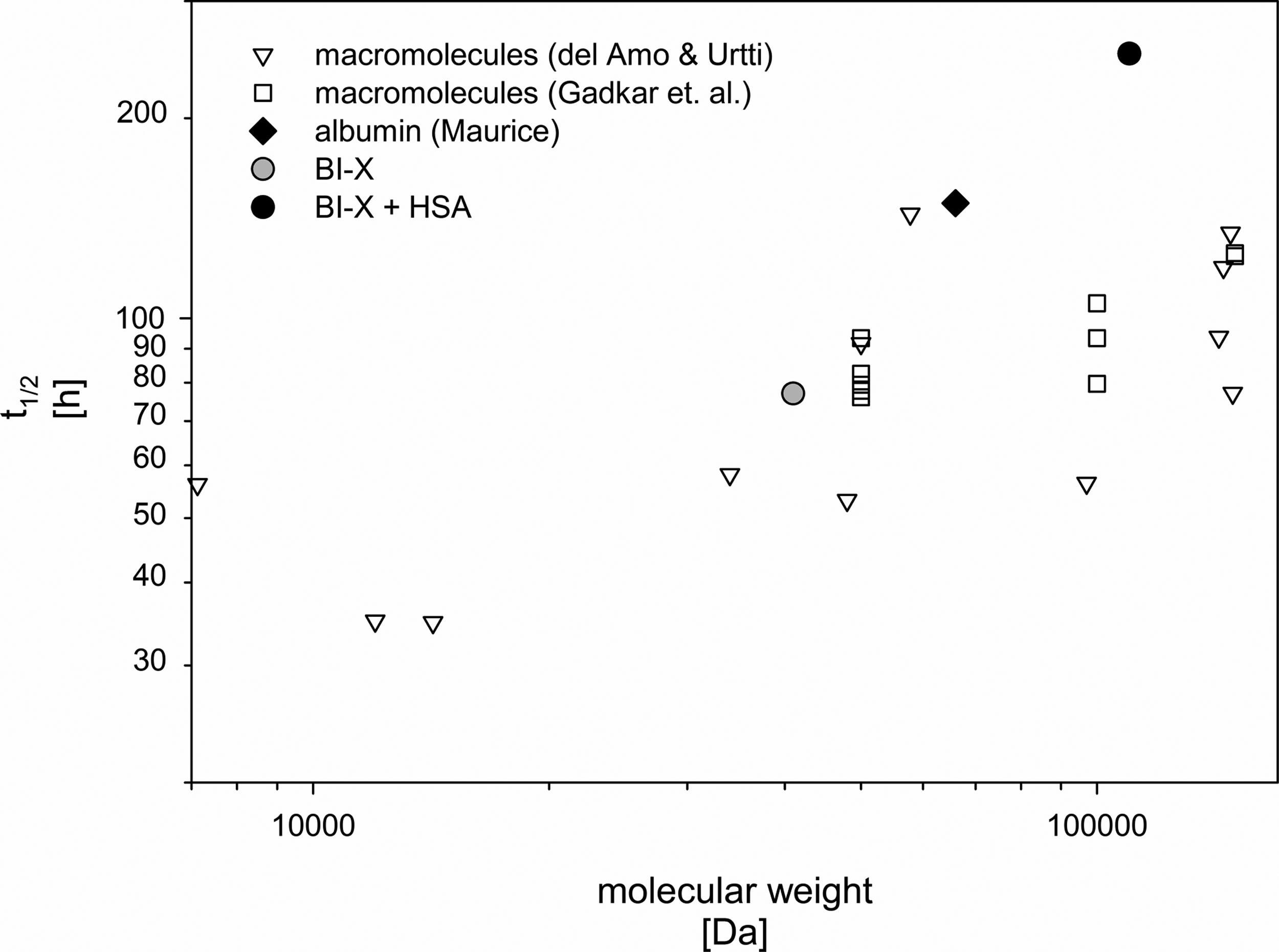
As mentioned before, a dose of 0.5 mg/eye of the nanobody would allow equimolar binding to vitreous albumin in normal human eyes. The vitreal albumin concentrations are markedly higher in diseased eyes; therefore, even for higher doses, sufficient albumin would be available. Only in case of doses exceeding the ocular albumin amounts, the individual albumin concentration may be a source for interpatient variability of the pharmacokinetics of the compound.
In conclusion, the present data clearly demonstrate that binding of an intravitreally injected compound to albumin has the potential to prolong its ocular half-life and may therefore be an interesting option to optimize the ocular pharmacokinetic properties to minimize the injection frequency. In addition, rats may be considered as suitable species for mechanistic studies. For quantitative assessment of vitreal PK, the rabbit is preferred as vitreal sampling is easy and the 2 main physiological properties that drive intraocular PK, that is, AH flow rate and vitreal volume, are much closer to humans than in small rodents.
Footnotes
Acknowledgments
The authors would like to thank Heidi Aßfalg, Janine Werner, and Daniel Hösch for expert technical assistance (rat experiments, ELISA measurements, and affinity determinations, respectively). The rabbit study was conducted at IRIS Pharma, Nice, France, by Dr. Solveig Lefranc-Jullien as study director. In addition, the authors thank Dr. Achim Sauer and Michel Garneau for critically reading the article.
Author Disclosure Statement
Both authors are employees of Boehringer Ingelheim.