
Abstract
Abstract
Diadenosine tetraphosphate abbreviated Ap4A is a naturally occurring dinucleotide, which is present in most of the ocular fluids. Due to its intrinsic resistance to enzyme degradation compared to mononucleotides, this molecule can exhibit profound actions on ocular tissues, including the ocular surface, ciliary body, trabecular meshwork, and probably the retina. The actions of Ap4A are mostly carried out by P2Y2 receptors, but the participation of P2X2 and P2Y6 in processes such as the regulation of intraocular pressure (IOP), together with the P2Y2, is pivotal. Beyond the physiological role, this dinucleotide can present on the ocular surface keeping a right production of tear secretion or regulating IOP. It is important to note that exogenous application of Ap4A to cells or animal models can significantly modify pathophysiological conditions and thus is an attractive therapeutic molecule. The ocular location where Ap4A actions have not been fully elucidated is in the retina. Although some analogues show interesting actions on pathological situations such as retinal detachment, little is known about the real effect of this dinucleotide, this being one of the challenges that require pursuing in the near future.
Introduction
D
The discovery of these molecules occurred in the 60s of the previous century, when scientists discovered some interesting changes in circular dichroism spectra 2 when studying the reaction of assembly between amino-acids and their corresponding transfer RNA (tRNA).3–5 Originally, these molecules were investigated as intracellular messengers involved in aspects such as the initiation of DNA replication process, including their role in the duplication of DNA when this nucleic acid is damaged, which were controversial.6,7 Also, the relationship of dinucleoside polyphosphates with FHIT (fragile histidine triad) protein, a tumor suppression gene, has been an interesting point of research relating this group of nucleotides with carcinogenic processes. 8
The perspective of dinucleoside polyphosphates as intracellular messengers was expanded when they were found as constituents of platelet-dense granules, 9 chromaffin granules,10,11 and synaptic vesicles, 12 therefore becoming potential extracellular messengers. 13 Dinucleoside polyphosphates and particularly diadenosine polyphosphates (ApnA) have become, together with adenosine triphosphate (ATP) and adenosine, the third element of the purinergic system. 14
Most of the actions of the dinucleotides, including Ap4A, are carried out by acting on membrane P2 receptors. Of the two families of P2 receptors, P2X or ionotropic, and P2Y or metabotropic receptors, diadenosine polyphosphates can bind to both but not to all of them and not with the same affinity. 15
In this study, we wish to present the actions of the dinucleotide Ap4A, in the ocular structures, indicating their physiological roles as well as the possible therapeutic applications.
Presence of Ap4A in Tears
The presence of Ap4A in the tear film was interesting due to its properties to enhance the secretion of some tear components and to increase the wound healing rate. 16 Ap4A was discovered simultaneously with its congener Ap5A, in New Zealand rabbit tears. 17 Diadenosine tetraphosphate concentration was 2.92 ± 0.28 μM and diadenosine pentaphosphate concentration was 0.587 ± 11 μM. Human tears also contain measurable amounts of both dinucleotides. 18 Apart from the presence of Ap4A and Ap5A, it was also possible to measure some levels of Ap3A. In human tears, the most representative dinucleotide is Ap4A whose concentration is several folds higher than the others. In this sense, the levels of diadenosine polyphosphates are 2.07 ± 2.2 nM, 108.7 ± 18.3 nM, and 37.07 ± 6.2 nM for Ap3A, Ap4A, and Ap5A. These dinucleotides can activate P2 purinergic receptors (inotropic P2X receptors and metabotropic P2Y receptors)19,20 localized on different ocular surface structures such as the cornea and conjunctiva. 21 Their presence in tears is a consequence of shear stress induced on the corneal epithelial cells. Srinivas et al. found also, in cultured rabbit corneal epithelial cells, that the secretion of ATP was due to a shear stress. 22 Carracedo et al. demonstrated that shear stress stimulus induced Ap4A and Ap5A release from human corneal epithelial cells both in in vitro and in vivo experiments. 23 The motion of the upper eyelid during the blinking process is the responsible to exert shear stress in the epithelial cells and therefore to raise the levels of both diadenosine polyphosphates, concomitantly with the increase in the blinking frequency.
The concentration of diadenosine polyphosphates in tears and, in particular Ap4A and Ap5A, has been studied in different conditions and situations. The most important is the dry eye (Table 1).24,25 The association between the increase of Ap4A in tears and dry eye pathology has been proposed as a potential molecular biomarker for this disease. In a symptomatic dry eye patient without Sjögren's syndrome but with normal tear production, Ap4A and Ap5A concentrations were 5- and 1.5-fold higher than healthy subjects, respectively. In the same kind of patients but with low tear production, diadenosine polyphosphate concentrations were increased by 100- and 345-fold than healthy subjects. 25 Moreover, Ap4A concentration increased 42-fold and Ap5A concentration 595-fold in Sjögren's syndrome patients, compared with normal individuals. 24 The relationship between diadenosine polyphosphates and dry eye could be explained by the high blinking rate in a dry eye patient, for compensating the tear instability. 26 Gender differences have been also described. 25 Symptomatic women apparently had higher levels of diadenosine polyphosphates than men. These differences may be due to a hormonal factor that may deserve further investigation.
The values are mean ± SD taken from the indicated references.
SD, standard deviation; SEM, standard error of the mean.
Ap4A and other dinucleotide concentrations have been evaluated in other conditions such as a congenital aniridia and keratoconus.27,28 In the case of congenital aniridia patients, the diadenosine polyphosphate concentrations increase compared with healthy subjects and are related with age and aniridia pathophysiology. 27 It has been hypothesized that the increased levels of diadenosine polyphosphates act as a compensatory mechanism to stimulate the proliferation and differentiation of limbal stem cells, which are deficient in aniridia disorder. On the contrary, keratoconus patients showed high levels of Ap4A in tears, 20-fold higher than healthy subjects. 28 This concentration may be due to mechanical stimulation of epithelial cells resulting from eye rubbing or due to greater degrees of corneal staining.
Refractive surgeries or contact lens wear are situations where ocular surface integrity could be affected.29,30 Both refractive therapies are related with dry eye and corneal injury.31–34 Patients who were underwent laser in situ keratomileusis or photorefractive keratectomy showed high Ap4A concentrations only during the first day after surgery, probably to help accelerating the healing process. 29 In the case of contact lens wear, a pilot study reported more Ap4A concentrations in soft contact lens wearers. 16 In addition, patients who wore gas-permeable contact lenses on daily basis wear, showed an increase of Ap4A concentration by lens friction during blinking. However, in patients who wore reverse-geometry rigid gas-permeable lenses overnight for orthokeratology treatment, Ap4A increased during the first day, returning to baseline afterward due to the lack of blinking while sleeping. 30 Moreover, Ap4A concentrations decreased statistically significantly after 8 h of scleral lens wear. 35 It was expected that diadenosine tetraphosphate concentration remained stable due to the lack of shear stress performed during blinking, but surprisingly the concentration was lower. This fact could be due to Ap4A degradation to its metabolites since the tear is stagnant behind the contact lens during 8 h.
Effect of Ap4A on Tear Secretion
Tear secretion is a physiological process because the tear forms the interface between the air and ocular tissues. To lubricate and to provide nutrients to the ocular surface tissues and to protect against infections are the more critical features of the tear film. For a correct tear volume and stability, it is very important to maintain a compensated tear composition. A failure in the tear secretion or composition may provoke several negative effects in the ocular surface such as discomfort, dryness, irritation, or corneal and conjunctival staining. 36 Diadenosine tetraphosphate participates in the secretion process of some compounds of tear film as aqueous component, mucins, and proteins.
Diadenosine tetraphosphates have demonstrated their capability to increase the tear volume in the ocular surface. 17 In the case of Ap4A, the increment is up to 60% compared to baseline values in experiments performed with New Zealand white rabbits. Also, Ap5A and Ap6A significantly increased tear production by about 20%, but Ap2A and Ap3A are completely ineffective. However, it has not been fully clarified if the effect of these dinucleotides is to induce tear production at the lacrimal gland or in other secretory structures. The studies performed with animal models suggest that diadenosine polyphosphates act as agonist of P2Y2 receptors for increasing the tear production (Table 2).17,37
IOP, intraocular pressure.
Some authors have reported that mononucleotide instillation increases the mucin secretion via the activation of metabotropic P2Y2 receptors located in goblet cells and ductal cells in the nerve terminals.37,38 In a model of dry eye developed in rats, the topical application of the dinucleotide Up4U (INS 365) increased the release of glycoproteins from goblet cells. 39 In a recent study with the commercial Up4U, performed in dogs, the MUC5AC concentration significantly increased after administration of 3% diquafosol ophthalmic solution. 40 Regarding diadenosine tetraphosphate, the effect of Ap4A on mucin secretion, evaluated in New Zealand rabbits by conjunctival impression cytology and confocal laser scanning microscopy visualization, has been demonstrated. Ap4A increases the mucin production by goblet cells but in lesser amounts than Up4U. 41
Ap4A is also involved in protein secretion in tears, mainly lysozyme and lactoferrin. Lysozyme constitutes around 20%–40% of the total tear protein 42 and has the capacity to dissolve gram-negative bacterial walls by the enzymatic digestion of mucopolysaccharides. 43 Lactoferrin is an iron-binding protein, thus reducing the availability of iron necessary for microbial growth and survival. In addition, anti-inflammatory actions of this protein have also been proposed. 44 Both proteins have been considered as one of the essential elements of the protective tear film barrier against ocular infection. 45
A single-dose application of Ap4A in a concentration of 100 μM in the New Zealand rabbit eye induced an increase in the lysozyme concentrations, up to 93% compared with basal levels, which was measurable for several hours after the topical application of this diadenosine polyphosphate. 46 The same experiment, but measuring lactoferrin concentrations, showed a transient but significant increase of lactoferrin concentration in rabbit tears, observed after topical application of Ap4A, from 68 ± 4 μg/mL to 83.17 ± 9.89 μg/mL. 47 The results obtained with nonselective P2 receptor antagonists indicated that the increase of these proteins is mediated by P2 receptors. Both proteins are produced by the acinar cells of the lacrimal gland44,48 and it has been reported that these acinar cells of the lacrimal gland express metabotropic P2Y1, 2, 4, 12, 14 receptors. 49 Since lysozyme and lactoferrin are critical against microbial infection, Ap4A could provide additional protection against pathogen invasion into tears by increasing the levels of this natural antimicrobial agent in healthy human subjects.
Effects of Ap4A on Corneal Wound Healing
After cornea injury, the wound healing process takes place to regenerate normal epithelium. 50 Corneal wound healing involves three consecutive phases: lag phase, cell migration, and cell proliferation. Diadenosine tetraphosphate can influence in the healing processes participating in the last two steps. Regarding its effect on cell migration, it has been described that the diadenosine polyphosphates Ap3A and Ap5A delay the migration rate of primary corneal rabbit epithelial cells. 51 This effect was mediated by P2Y6 stimulation as revealed by assays with P2 agonists and antagonists (Table 1).
In contrast, the diadenosine polyphosphate Ap4A was able to increase the rate of healing by 130% in in vivo experiments with New Zealand white rabbits. 52 Likewise, the dinucleotide also accelerated the rate of corneal migration in primary cultures of rabbit corneal epithelial cells and immortalized statens seruminstitut rabit cornea cells. 53
This opposite behavior of the dinucleotides on cell migration may be explained by the activation of different P2 receptor subtypes. In fact, pharmacological studies using different P2 agents as well as experiments performed with a specific P2Y2 small interfering RNA (siRNA) demonstrated the involvement of the P2Y2 receptor in the effect evoked by Ap4A on wound healing process.51,54
The precise signaling pathways that link Ap4A-induced P2Y2 activation to the increase of reepithelialization rate were also examined. Analysis of P2Y2 downstream effectors showed that the action evoked by Ap4A on corneal epithelial wound repair was mediated by the activation of phospholipase C/protein kinase C (PLC/PKC) pathway leading to extracellular signal-regulated kinases 1/2 (ERK1/2) and rho-associated coiled-coil-containing protein kinase 1 (ROCK-I) activation.53,55 Moreover, the acceleration in corneal epithelial cell migration induced by Ap4A was reverted by tyrphostin AG1478, a selective inhibitor of endothelial growth factor receptor tyrosine kinase. 53 This result suggests that receptor tyrosine kinase-dependent mechanisms could also contribute to the effects induced by Ap4A in these cells during the reepithelialization process.
After cell migration to cover the damaged area with a cell monolayer, the next step in corneal wound healing is to initiate the mitotic process to restore normal corneal epithelial thickness. Experiments performed in vitro showed that corneal epithelial cell proliferation was increased by Ap4A or Ap3A in a similar manner at 24 and 36 h. 56
Ap4A produced its effect on proliferation by activation of a P2Y2 receptor coupled to canonical PLC/PKC/RhoA/ROCK-I and ERK1/2 pathways. In the case of Ap3A, this diadenosine polyphosphate stimulated a P2Y6 coupled to p38 pathway activation to complete the whole proliferation process. 56
Altogether, these findings invite us to consider the diadenosine polyphosphates, particularly the dinucleotide Ap4A, as an attractive therapeutic agent to treat wound healing-related diseases in the cornea.
Regulation of Corneal Barrier Permeability by Ap4A
The corneal epithelium provides a defensive barrier preventing the entrance of potentially harmful substances but also limiting helpful ocular drug delivery. This barrier function of the corneal epithelium depends on tight junctions that seal the intercellular space. Tight junction assembly and consequently barrier formation can be influenced by different agents and factors. Among them, it has been recently described the role of the Ap4A as a modulator of tight junction assembly and paracellular barrier function (Tables 2 and 3). 57
IOP, intraocular pressure; SD, standard deviation.
In vitro experiments with human corneal epithelial cells showed that the treatment with Ap4A induced a clear disruption of ZO-1 (Zonula occludens 1) distribution as well as a significant decrease in the ZO-1, occludin, and claudin-7 protein levels compared to control cells (Fig. 1). Concomitantly, transepithelial electrical resistance values were dramatically reduced. These effects were maximal 2 h after the application of Ap4A, with protein levels and transepithelial electrical resistance values returning progressively to control values after 6 h of the Ap4A treatment. 57
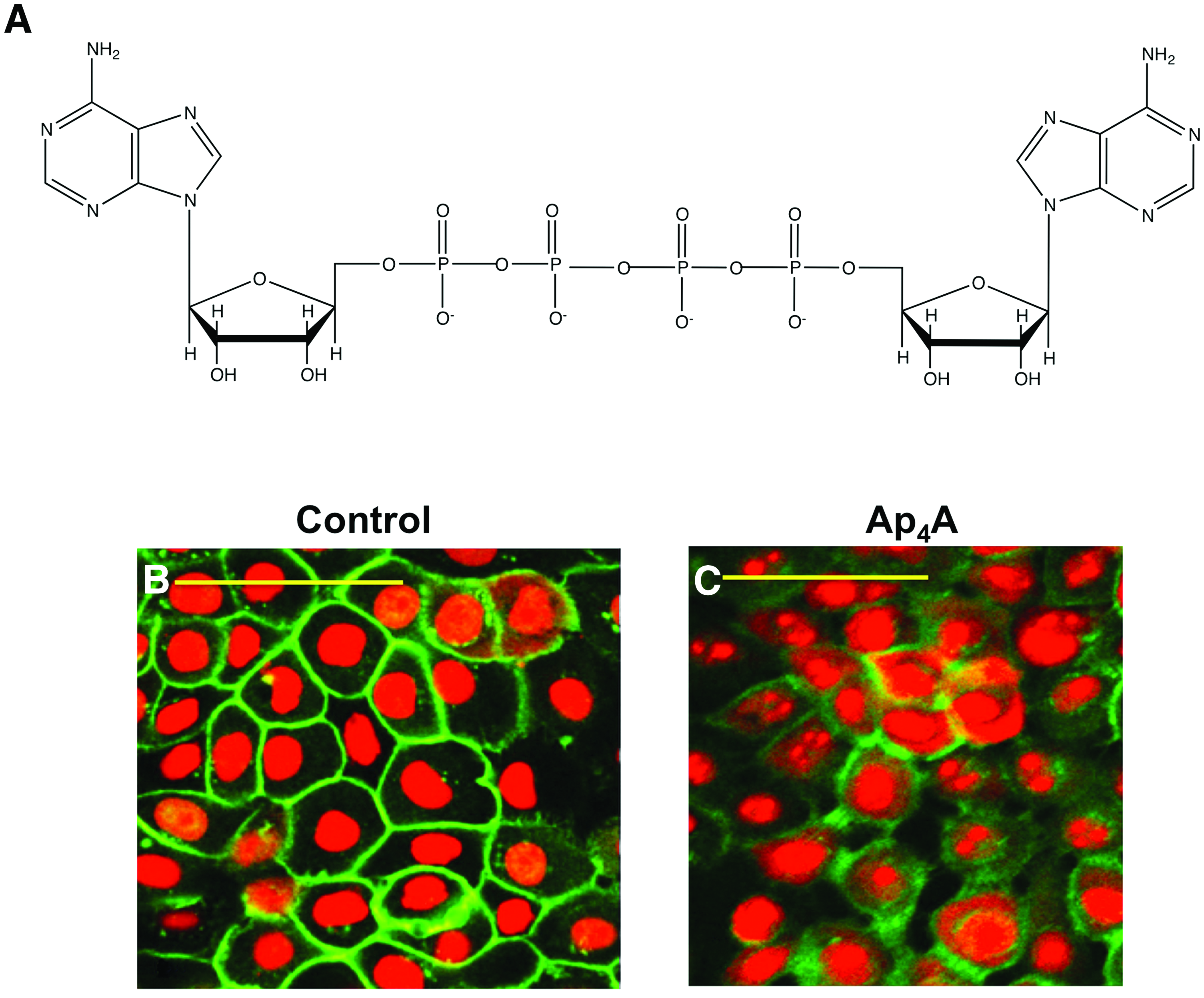
Structure of Ap4A and its effect on ZO-1 in human corneal epithelial cells.
The Ap4A-induced downregulation of tight junction protein levels was accompanied by phosphorylation of extracellular signal-regulated kinase and both effects were blocked by the ERK inhibitor U0126 and P2Y2 siRNAs, indicating that Ap4A reduces tight junction protein levels through P2Y2 stimulation and subsequent ERK activation. 57
To determine whether the increase in barrier permeability induced by Ap4A could enhance drug delivery through the cornea, additional in vivo experiments with New Zealand white rabbits demonstrated the access of hypotensive compounds such as the melatonin analogue 5-MCA-NAT (5-methoxycarbonylamino-N-acetyltryptamine) and the adrenergic drugs brimonidine and timolol (Table 3).57,58 Thus, when Ap4A was topically applied 2 h before drug instillation, increased levels of these compounds were found in the rabbit aqueous humor compared to the application of the drug alone and the hypotensive effect was also improved. Therefore, Ap4A application can improve ocular drug delivery and consequently therapeutic efficiency.
Considering the disadvantages associated with topical ocular administration of drugs by eye drops (i.e., low bioavailability, frequent instillation requirement, and potential systemic side effects), there is an unmet need for more effective delivery of ocular therapeutics in the ocular pharmacology market. In this context, Ap4A could have a remarkable impact to overcome low ocular bioavailability, reducing the concentration of the drug required as well as its potential side effects.
Presence of Diadenosine Tetraphosphates in the Aqueous Humor and Their Effects on Intraocular Pressure
Diadenosine polyphosphates are present, at low micromolar concentration, in the aqueous humor (AH) of rabbit and human eyes.59,60 This secretion fills the anterior/posterior chamber of the eye and thereby provides nutrients to avascular structures such as the cornea as well as lens. In addition, intraocular pressure (IOP) is determined by the balance between AH production (by the ciliary body) and its drainage (by the trabecular meshwork and the uveoscleral pathway). 61 Consequently, excessive production of AH or impairment of its drainage results in elevation of the IOP, which is one of the most important risk factors of glaucoma. 62 Since ciliary body and trabecular meshwork possess several P2 purinergic receptors, these endogenous dinucleotides could regulate IOP (P2 receptors in anterior/posterior chamber and dinucleotide receptors have been previously reviewed63,16). In fact, the topical application of diadenosine polyphosphates in normotensive rabbits can modify IOP. 60 For instance, Ap4A reduces IOP, while other dinucleotides, including Ap2A, Ap3A, and Ap5A, increase IOP, all in a concentration-dependent manner. 60 Hypotensive action of Ap4A seems to be mediated by a P2X2 receptor localized on cholinergic terminals that innervate the ciliary body.60,64,65 When Ap4A activates this receptor, acetylcholine is released and it contracts the ciliary muscle that finally reduces hydrodynamic resistance to the AH drainage. 64 In addition, Soto et al. have demonstrated that this dinucleotide also facilitates trabecular AH outflow via activation of a P2Y1 receptor localized on bovine trabecular meshwork cells (Table 1). 66 Interestingly, comparing the hypotensive effect of Ap4A to commercial compounds such as Xalatan, Trusopt, Timoftol, Azopt, and pilocarpine, the reduction was equivalent to most of them, and only surpassed by Timoftol or by the synthetic analogue of ATP, βγ-Me ATP (Fig. 2).

Comparative effect of Ap4A versus commercial antihypertensive compounds and other nucleotides. Column plot showing the reduction in IOP for the five compounds commonly used for glaucoma treatment as well as Ap4A and βγ-Me ATP and UTP. The values are mean ± SEM, obtained after the application of 40 μl of the commercial compounds or the nucleotides. ATP, adenosine triphosphate; IOP, intraocular pressure; SEM, standard error of the mean; UTP, uridine-5′-triphosphate.
More recently, Guzman-Aranguez et al. have reported the ability of diinosine polyphosphates to modify IOP in normotensive rabbits. 67 Among all the tested dinucleotides, Ip5I increases IOP, while others such as Ip3I and Ip4I reduce IOP. 67 Ip4I is the most potent hypotensive compound and its effect seems to be mediated, at least in part, by a P2Y receptor. 67
The Potential Therapeutic of Diadenosine Tetraphosphate in Glaucoma
Like it was mentioned before, diadenosine polyphosphates and their receptors are present in the anterior and posterior chambers of the eye. Also, exogenous dinucleotides evoke changes in IOP. All these data indicate that dinucleotides may be involved in the development of glaucoma. In this sense, Castany et al. have reported higher Ap4A and Ap5A concentrations (15- and 5.4-fold, respectively) in the AH of glaucomatous patients compared to normal individuals. 59 For this reason, the authors suggest their use as potential glaucoma biomarkers. 59
Glaucoma is an optic neuropathy that causes progressive visual field loss and is due to a continual retinal ganglion cell (RGC) death and to the loss of their axons. 62 One of the most important risk factor for glaucoma is high and persistent IOP. 68 High IOP produces an excessive release of the neurotransmitter glutamate that evokes an increase in free radicals and finally RGC death.69,70 For this reason, an effective therapeutic approach to treat glaucoma would be to lower IOP and promote the survival of RGCs. 68 In this context, Hoyle and Pintor have demonstrated the neuroprotective role of dinucleotides. 71 Thus, for example, exogenous Ap4A can preserve the sympathetic terminals that innervate the ciliary body of rabbit eyes from 6-hydroxydopamine-induced degeneration. 71
Ap4A in the Retina Nothing at All
The retina is one of the ocular tissues where a more abundant presence of purinergic receptors has been described.20,21,72,73 The purinergic system is fully represented in this part of the eye since apart from the presence of these receptors, the existence of enzymes responsible for the degradation of nucleotides, a nucleotide vesicular transporter, has also been detected. 74 Altogether nucleotides and dinucleotides may play important roles acting in the retina as autocrine and paracrine mediators.
Although the most representative nucleotide in the retina is ATP,74,75 little is known about the presence of Ap4A and other diadenosine polyphosphates in the retina. Classically, diadenosine polyphosphates, including Ap4A, have been described as cotransmitters with others such as acetylcholine and monoamines, 13 and it would not be surprising to see that, as it happens in other parts of the nervous system, these molecules are released and exert actions via some P2 receptors. In this regard, and considering the plethora of P2 receptors present in the retina, it is important to note that not all of them would be targets for Ap4A and other dinucleotides. Considering the possible effect of dinucleotides on P2 receptors, P2Y1, P2Y2, and P2X2 or P2X3 are the best candidates, since P2X7 receptors do not bind diadenosine polyphosphates unless the concentrations are above 1 mM.16,63 With this panorama, diadenosine tetraphosphate action can only be on metabotropic P2Y1, P2Y2, P2Y4, and P2Y12 receptors present in the retina21,76,77 and P2X1, P2X2, P2X3 ionotropic receptors78–81 in neural retina and in retinal pigmented epithelium.82,83
Thinking about which receptors and cells could be stimulated by Ap4A, some ideas can be suggested: for instance, the presence of P2X2 receptors exists on different neuronal populations, including photoreceptors, ganglion cells, and amacrine cells. The P2X2 signaling is implicated in retinal modulation in the OFF-pathway84,85 Similar labeling pattern was described for P2X3 receptors, finding positive immunoreactivity in bipolar, amacrine, ganglion cell, and plexiform layers. The implication of P2X3 receptor could be related with neurotransmission and modulation of outer and inner retinal processing. Considering this, in several studies it has been suggested the possible role of P2X in the modulation of inhibitory response of GABAergic amacrine cells.86–88 Furthermore, it is also to be considered that these P2X physiological properties are known to be determined by the presence of various P2X subunits.
The expression of P2Y receptor subtypes P2Y1 and P2Y2, targets for Ap4A, was detected in RGCs, bipolar cells, photoreceptors, and Müller cells.76,77 There are subsets of rat retinal neurons expressing P2Y1 receptors, especially subclasses of cholinergic amacrine cells and ganglion cells 89 that may be stimulated by diadenosine tetraphosphate.
Concerning the potential actions of Ap4A in the retina, we can infer an interesting one according to the results obtained by other synthetic dinucleoside polyphosphates. In this way, the intravitreal or subretinal treatment with INS37217, P2Y2 receptor agonist, stimulates subretinal fluid reabsorption in experimental retinal detachment. 90 Furthermore, other studies showed the enhancement of electroretinographic responses after retinal detachment due to P2Y2 receptor agonist injections.91,92 Considering all these findings and possibilities, Ap4A may be implicated in the processing of visual function via P2X and P2Y receptors and could be suggested as a potential pharmacological approach in the retinal detachment treatment and other retinal diseases.
Conclusions
Diadenosine tetraphosphate is an amazing molecule that presents a variety of actions in most of the ocular structures. Its physiological actions vary from the ocular surface to the regulation of IOP and presumably modulating retinal activity. This dinucleotide is an attractive molecule, more resistant to hydrolysis than mononucleotides, but we need the development and testing of its synthetic analogues to really understand its real therapeutic potential. Ap4A analogues with methylene or amino bridges between the phosphates may introduce a new group of active nucleotides that may be useful in the treatment of ocular surface diseases, ocular hypertension, and retinal detachment among others.
Footnotes
Acknowledgments
This work has been supported by the research grants SAF2013-44416-R, SAF2016-77084-R, RETICS RD12/0034/0003, and Universidad Complutense PR1/07-14890.
Author Disclosure Statement
No competing financial interests exist.