
Abstract
Abstract
Purpose:
To investigate the therapeutic potential of a prostacyclin (IP) receptor agonist for ocular inflammation and the effect on immune cells.
Methods:
The anti-inflammatory activities of cicaprost were determined in primary human monocyte-derived macrophages and human monocyte-derived dendritic cells (MoDC), as well as a lipopolysaccharides (LPS)-induced rat uveitis model. Multiple cytokine release was measured by utilizing Luminex Technology. Prostacyclin (IP) Receptor expression was detected by reverse transcription–polymerase chain receptor. Leukocyte infiltration and protein exudation in the rat uveitis model were measured using a hemocytometer and protein concentration by a NanoDrop instrument.
Results:
Cicapost, an IP receptor agonist, potently inhibits proinflammatory chemokines/cytokine production not only from LPS- or TNFα (tumor necrosis factor-alpha)-induced primary human monocyte-derived macrophages, but also from LPS-stimulated MoDC. While constitutively expressed in macrophages, the IP receptor was inducible by LPS stimulation in MoDCs. In a LPS-induced rat uveitis model, cicaprost efficaciously prevents ocular inflammatory cell and protein leakage, as well as inflammatory cytokine release.
Conclusion:
The IP receptor agonist cicaprost is a potent anti-inflammatory agent, implicating that the tightly controlled PGI2/IP signaling pathway is important in regulating inflammation. This response could be harnessed in ocular inflammatory disease where steroids are currently the standard of care.
Introduction
P
Low or absent expression of mRNA of IP receptors have been observed in human spleen and thymus 7 and while there is a marked expression in the thymus of rodents, this seems to have minimal impact on murine T cell maturation. 3 In human monocytes, the PGI2 analog iloprost activates IP receptors to inhibit interferon-gamma and interleukin (IL)-stimulated production of cytokines tumor necrosis factor-alpha (TNFα), IL-8, and regulated on activation, normal T cell expressed and secreted (RANTES). 8 PGI2 analogs are also able to suppress TNFα synthesis in lipopolysaccharides (LPS)-induced human leukocytes.9,10 Recent studies in mice have shown that PGI2-IP signaling can play an important role in determining T cell functional differentiation. 11 In addition to inhibiting vascular thrombosis and homeostasis of circulation, PGI2-IP signaling seems to play important roles in regulating adhesion, chemotaxis, and activation of blood cells. 6
Although PGI2 analog beraprost was found protecting retinal pericytes from advance glycation end products-induced cytotoxic injury, 12 pharmacological studies of PGI2-IP in ocular tissues were vastly focused on exploring their potential effects on intraocular pressure.13,14 PGI2 breaks down rapidly and nonenzymatically to biologically inactive hydrolysis products 6-keto-PGF1α in cells 15 indicating that it participates in a tightly controlled signaling pathway. In the current study, we evaluated the effects of PGI2-IP signaling in human primary immune cells and in an animal model of ocular inflammation as a potential therapeutic. We present that the IP receptor agonist cicaprost is a potent inhibitor of a variety of cytokines released from LPS- or TNFα-induced human monocyte-derived macrophages and dendritic cells, and highly efficacious in treating LPS-induced uveitis in a rat model.
Methods
Cells
Frozen human peripheral blood mononuclear cells (PBMCs) were purchased from Biomed Supply (Carlsbad, CA) obtained from healthy donors. CD14+ monocytes were isolated from PBMCs using the Monocyte Isolation Kit II according to the manufacturer's instruction (Miltenyi Biotech, Auburn, CA).
Reagents
DC Media includes RPMI 1640, Penn/Strep, 10 mM HEPES, 1 × nonessential amino acid (Invitrogen, Carlsbad, CA), 10% heat-inactivated defined fetal bovine serum (FBS) (Hyclone, Logan, UT), and 5.5 × 10−6 2-ME (Sigma, St. Louis, MO). In some experiments as outlined in the following sections, 2.5% of heat-inactivated charcoal and dextran-stripped FBS was supplemented instead. Cicaprost (IP agonist) was synthesized at Target Molecules (Southampton, England) and RO-1138452 (IP antagonist) was obtained from Cayman Chemicals (Ann Arbor, MI). LPS was obtained from (Sigma, St. Louis, MO).
Macrophage stimulation assays
Human CD14+ monocytes were seeded on day 0 at 1–2 × 105 cells/well of 96-well plates. To differentiate monocytes to macrophages, cells were maintained in RPMI-1640 (Invitrogen, Carlsbad, CA) with 10% fetal calf serum (Gemini Bio-Products, West Sacramento, CA), 33 ng/mL recombinant human monocyte colony

Cicaprost potently inhibits chemokine/cytokine secretion from human macrophages induced by
MoDC stimulation assays
MoDCs were derived as previously described. 16 Stimulation assays were carried out in DC media containing 2.5% heat inactivated, charcoal and dextran-stripped FBS. MoDCs were plated at 5 × 104/well in 96-well U-bottom plates in the presence of LPS (100 ng/mL) and various concentrations of cicparost, DMSO, or RO-1138452 (as indicated in the figure legends). Cells were preincubated for 10 min with the antagonist, treated with cicparost for 10 additional minutes and then stimulated under the above conditions. All cultures were incubated at 37°C in 5% CO2 incubators for 18–20 h. At the end of the assay, plates were centrifuged and the supernatants were harvested and frozen for later cytokine determination. MoDCs were monitored for viability by the alamarBlue assay (Invitrogen, Carlsbad, CA). MoDCs were resuspended in 90 μL of fresh medium and 10 μL of alamarBlue was added; the cells were incubated for 3 h at 37°C and read on a variable wavelength plate reader (SpectraMax Gemini EM; Molecular Devices, Sunnyvale, CA). Each condition was performed in triplicate from at least 3 donors.
Luminex assays
Cell media of macrophage and MoDC were tested by Luminex bead-based assay according to the manufacturer's instructions (Millipore, San Jose, CA). Human cytokine panels were used to analyze cytokine and chemokines on a Luminex IS-200 (BioRad, Hercules, CA).
AlamarBlue cell viability assay
After macrophage stimulation assay, 10 μL of alamarBlue solution (Invitrogen) was added to each well of cells with 90 μL warm fresh Opti-MEM I medium in the 96-well culture plates. After incubation at 37°C for ≥4 h, cell plates were scanned for alamarBlue intensity using a SpectraMax GeminiEM fluorescent plate reader (Molecular Devices, Sunnyvale, CA) at excitation 530 nm and emission 590 nm. The results are used for data normalization in Fig. 1.
RT-PCR and qPCR assay
Differentiated MoDCs were isolated, washed twice in phosphate-buffered saline, and mRNA isolated using the Qiagen RNeasy Kit (Qiagen, Germantown, MD). Reverse transcription–polymerase chain receptor (RT-PCR) was performed using Invitrogen SSIII one-step RT-PCR system with Platinum Taq DNA polymerase (Invitrogen, Carlsbad, CA) and the following primers for prostaglandin receptors:
Prostacyclin receptor (PTGIR) for 5′TGTGCTCCCTGCCTCTCACGATC-3′, rev 5′AGAGCTTGAGTCGCTGGAAGACAG-3′; GAPDH for 5′GAGTCAACGGATTTGGTCGTATTG-3′, rev 5′CTCGCTCCTGGAAGATGGTGATG-3′.
PCR products were run on a 3% agarose TBE gel and visualized on FlourChem HD system (Alpha Innotech, Santa Clara, CA). To perform quantitative PCR, RNA samples were reverse transcribed using the Roche's First Strand Synthesis Kit, followed by TaqMan assay (Qiagen, Germantown, MD). Primers of rat IP receptor and beta-actin were purchased from OriGene systems (Houston, TX).
LPS-induced uveitis rat model
All animal experimentation was performed in compliance and approval of the Allergan Institutional Animal Care and Use Committee (IACUC) and adheres to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. We employed the rat endotoxin-induced uveitis model. 17 Male Lewis rats (6 weeks of age, ∼200 g) were obtained from Charles River (San Diego, CA). All compounds and vehicle were applied to each eye in 10 μL of volume (concentrations as indicated in the figure legends). Cicaprost or vehicle (saline) was applied 1 h before model induction (injection of 100 μg of LPS [dissolved in saline] in the hind rear footpad) and applied twice during uveitis development (2 and 5 h). Rats were euthanized at 18 h and aqueous humor was collected and pooled bilaterally. In some experiments, Pred Forte (Prednisolone) was used as a positive control. Inflammatory markers (leukocyte infiltration and protein exudation) were measured using a hemocytometer and protein concentration by NanoDrop (ThermoFisher, Wilmington, DE). Additionally, cytokines present in the aqueous humor were measured through Luminex using the Rat Cytokine Kits from Millipore (San Jose, CA) according to the manufacturer's instructions.
Results
IP receptor agonist cicaprost inhibits cytokine release from LPS or TNFα-induced primary human macrophages
As shown in Fig. 1, cicaprost potently inhibited the release of proinflammatory cytokines IL-8, MCP-1, MIP-1α, MIP-1β, and RANTES induced by 10 ng/mL LPS (Fig. 1A) or 20 ng/mL TNFα (Fig. 1B), as well as TNFα induced by 10 ng/mL LPS (Fig. 1A), in primary human macrophages. Cicaprost was dosed from 1000–0.1 nM in 10-fold serial dilution compared with DMSO vehicle control. The range of EC50 of cicaprost is 0.1–0.8 nM for LPS-induced and 0.5–10 nM for TNFα-induced cytokines. Cell viability of each treatment condition was evaluated by alamarBlue assay, and data presented are normalized to cell viability.
IP receptor agonist potently inhibits LPS-activated DC cytokine release and is reversed by blockade using an IP antagonist
As part of an ongoing inflammatory response, monocytes are induced to differentiate into inflammatory dendritic cells in vivo. 18 These cells can be modeled in vitro using a protocol consisting of granulocyte and macrophage stimulating factor (GM-CSF)/IL-4 treatment of purified monocytes to induce dendritic cell differentiation. These cells are typically referred to as monocyte-derived dendritic cells (MoDCs). Using MoDCs we examined the effects of the IP agonist, cicaprost on LPS-activated MoDC cytokine release. MoDCs were activated with 100 ng/mL of LPS in the presence of cicaprost dosed from 1000 to 0.1 nM in a 10-fold serial dilution. Cicaprost potently inhibited IL-6, IL-8, MCP-1, TNF-a, MIP1β, and IL-23 (Fig. 2). Importantly, an IP receptor antagonist RO-1138452 reversed the inhibition by cicaprost (Fig. 2). The activity of the RO-113845 inhibitor alone on MIP-1β release suggest that there may be some endogenous PGI2 release, although none could be detected in the media (data not shown). 16
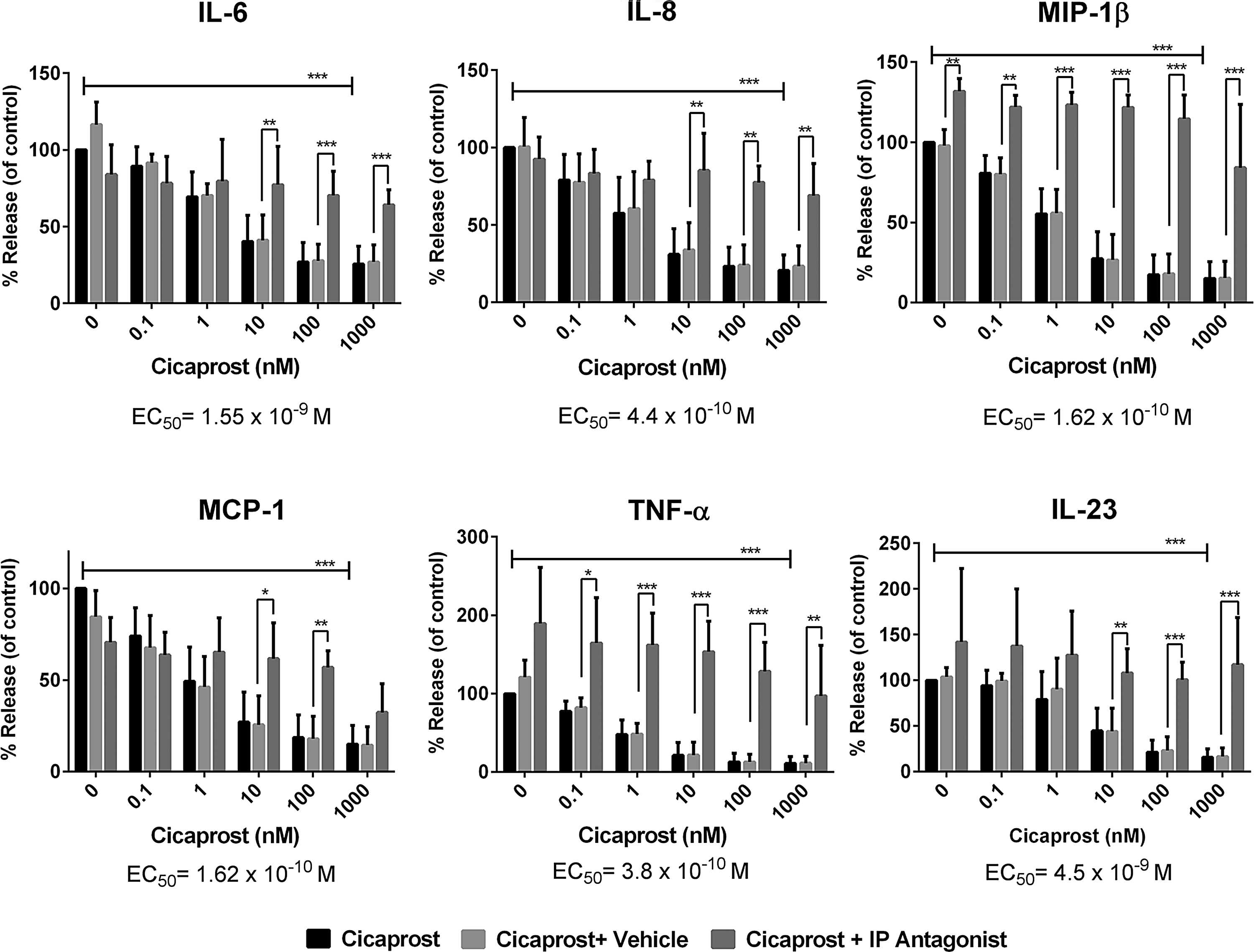
Cicaprost downregulates inflammatory cytokines through the IP receptor. MoDCs were plated at 5 × 104 cells/well and activated with 100 ng/mL of LPS in the presence of different concentrations of cicaprost, vehicle (DMSO), or cicaprost+an IP antagonist (RO-1138452). All cytokine results were normalized to LPS-stimulated untreated control (100%). The above data are the mean of 3 donors (n = 3). Statistical significance was determined by 2-way ANOVA (*P < 0.05, **P<0.01, ***P<0.001) using GraphPad PRISM software. One of 3 similar experiments is shown.
IP receptor is induced on dendritic cells upon LPS activation
After finding profound effects of the IP agonist, cicaprost, on LPS-induced cytokines, we examined the expression of the IP receptor by RT-PCR. To our surprise it was absent in resting MoDCs, although the parental monocytes had detectable expression (Fig. 3, data not shown 19 ). Given that MoDCs responded to agonist during LPS stimulation, we examined the expression of the IP receptor after LPS stimulation and found that it is upregulated following stimulation (Fig. 3).
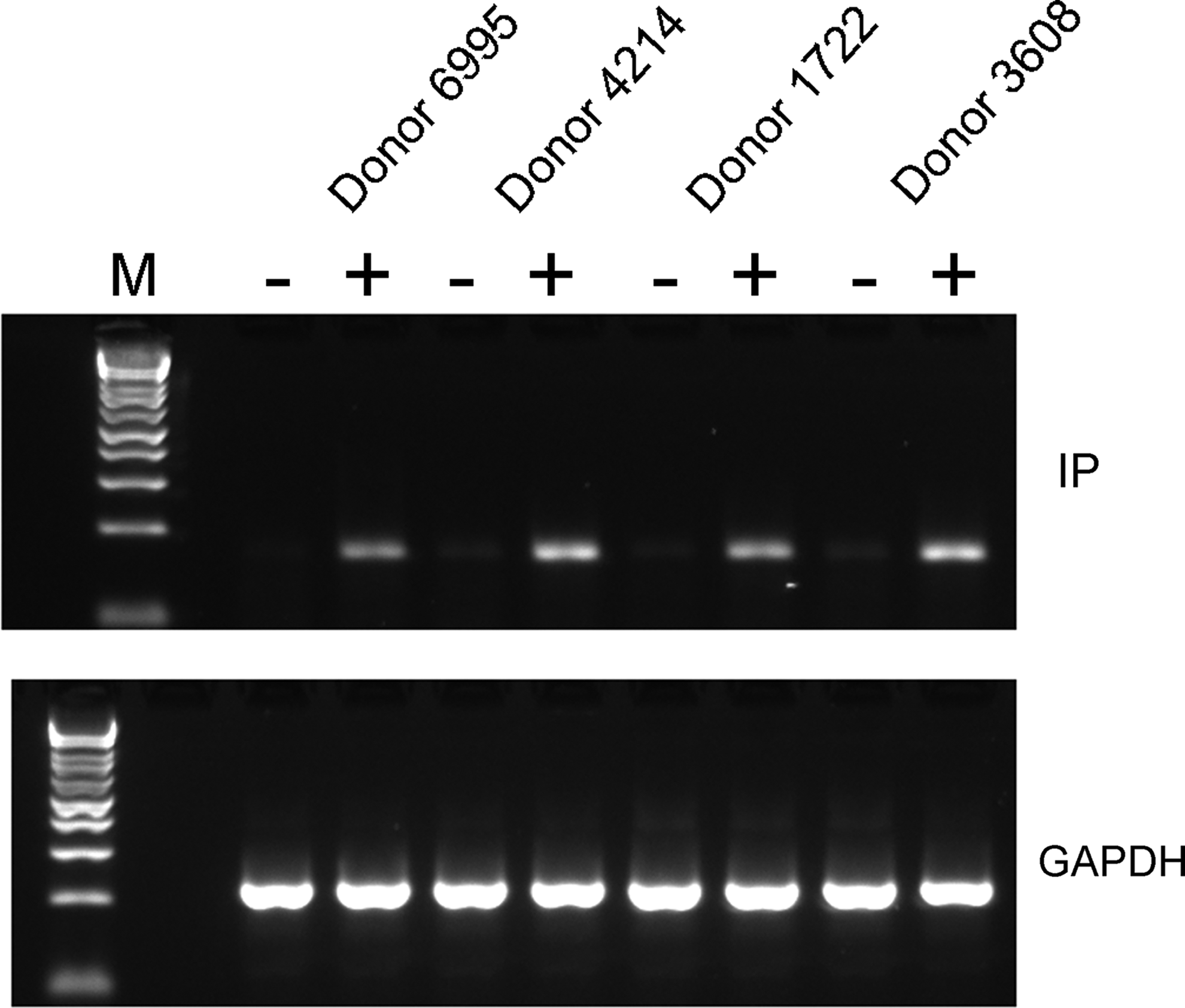
Expression of PTGIR is induced on LPS-stimulated MoDC cells. MoDCs were differentiated from isolated monocytes as described in the Methods section. Cells were cultured in 6-well plates at 1 × 106 cells per well with or without 100 ng/mL LPS for 24 h and harvested for RNA isolation. PTGIR expression was examined by RT-PCR. GAPDH expression was run as a positive loading control. Three donors (out of 4) are shown. Two replicate experiments were performed. PTGIR, prostacyclin receptor; RT-PCR, reverse transcription–polymerase chain receptor.
Cicaprost inhibits development of LPS-induced uveitis in rats
Due to the activities of cicaprost on innate immune cells (macrophages and DCs) and its potent ability to inhibit both inflammatory cytokines and chemokines, we decided to study it in an experimental model of uveitis. The LPS-induced rat uveitis model was chosen due to it having a high degree of infiltrating innate immune cells, being LPS dependent, and the presence of cytokines and chemokines that in our in vitro assays with macrophages and MoDCs, cicaprost inhibited (Figs. 1 and 2). To verify that the IP receptor was expressed in this model, rats were immunized with or without LPS and eyes were harvested after 18 h. IP receptor was detected in the retina and cornea, and less so in the conjunctiva, although appeared to be upregulated by LPS in all tissues (Fig. 4A). These data suggest that the receptor is expressed in the locations where immune cells (macrophages and dendritic cells) have previously been described.20,21 Cicaprost was formulated in saline and given as drops just before model induction and 2 times after as described in the Methods section. No nociceptive or inflammatory events were observed following cicaprost administration. Cicaprost was dosed at 0.001%–0.1% in each eye. This dosing resulted in significant inhibition of cellular infiltrate and protein leakage in a dose-dependent manner (Fig. 4B). Cytokine determinations from the collected aqueous humor also showed a dose-dependent decrease in IL-6, MCP-1, and RANTES (Fig. 4C). This inhibition was comparable to equivalent dosing of a topically administered steroid (prednisolone) (Fig. 5).
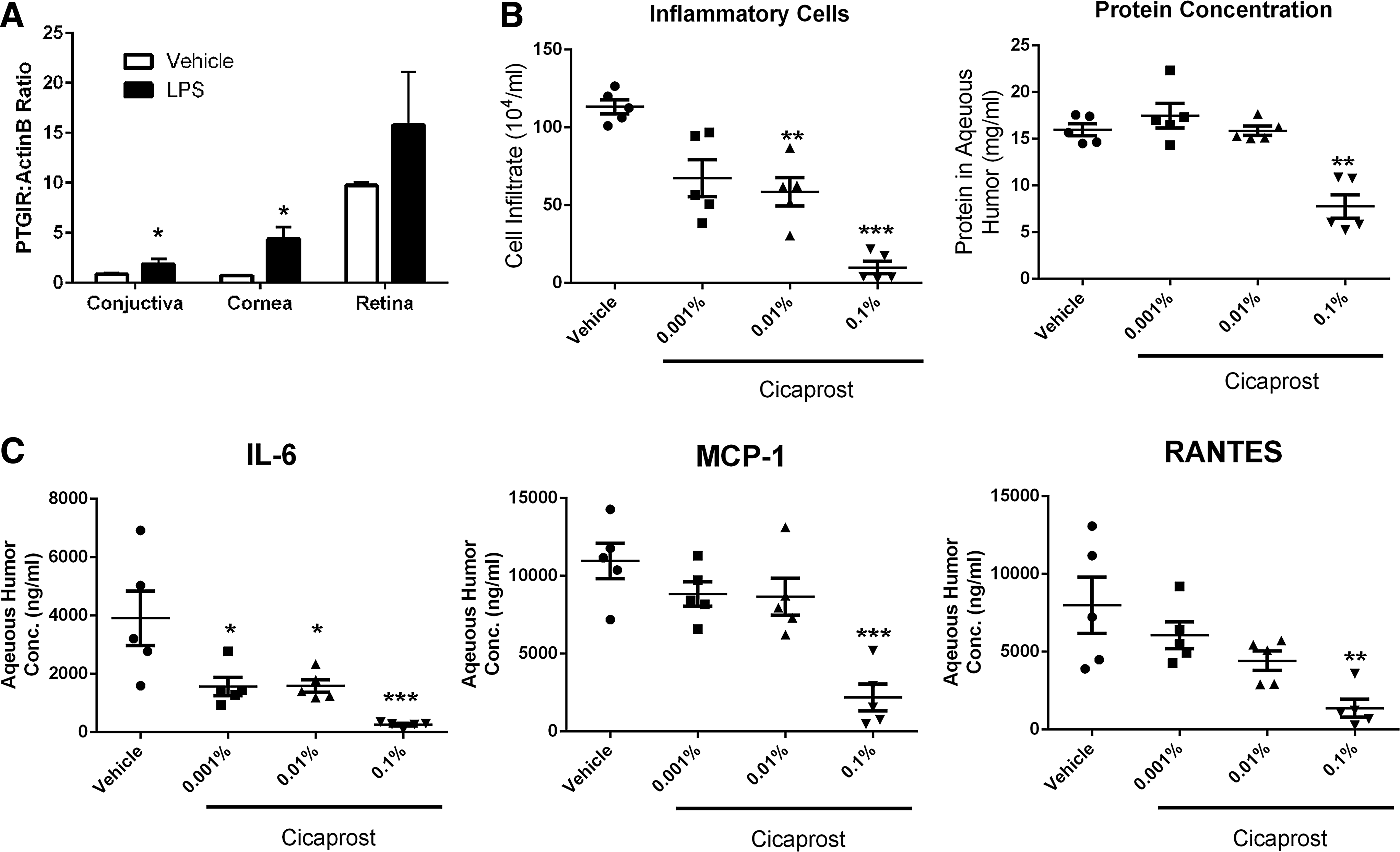
Cicaprost administered as an eye drop, inhibits uveitis development in a rat model. Approximately 200 g Lewis rats (5 per group) were given 3 treatments of topical cicaprost (or saline) bilaterally to each eye at −1, 2, and 5 h. At time 0, 100 μg of LPS was injected into the left hind footpad of the rat. Eighteen hours later the rats were euthanized.
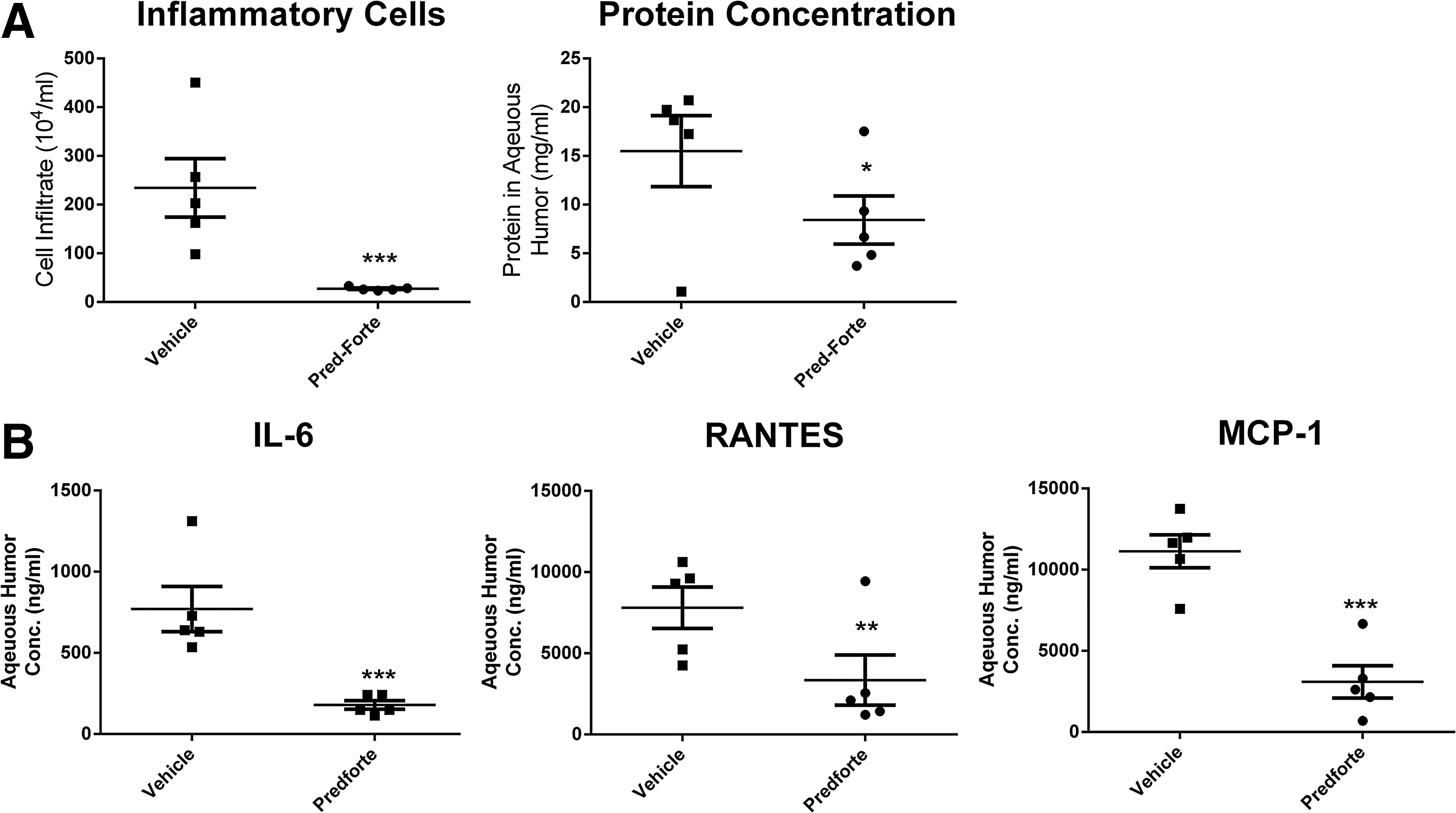
Pred Forte (prednisolone) inhibits uveitis development in a rat model. Approximately 200 g Lewis rats (5 per group) were given topical Pred Forte (1% prednisolone) or saline bilaterally to each eye at −1, 2, and 5 h. At time 0, 100 μg of LPS was injected into the left hind footpad of the rat. Eighteen hours later the rats were euthanized and aqueous humor was collected and
Discussion
Prostaglandins are at the threshold of the inflammatory/anti-inflammatory axis. Depending on the cell type, tissue location, and concentration, this diverse set of lipid mediators can mediate vastly different effects. 22 In this study we have shown that the PGI2 mimetic, cicaprost, profoundly inhibits cytokines produced by primary human macrophages and dendritic cells. This work builds on earlier findings that PGI2 mimetics can potently modulate APCs with respect to their ability to take up antigen and produce inflammatory cytokines such as TNFα.23,24 This has been shown to be cAMP dependent, which fits with the Gs-coupled nature of the IP receptor. 25 In several in vivo mouse models, PGI2 analogs have been shown to blunt inflammatory responses reviewed in Ref. 26 However, these studies have been mostly limited to cell lines and mice whose findings may not translate into humans. Indeed, although it has been shown that PGI2 mimetics influence mouse T cell differentiation and function, 26 we have found that the IP receptor is noticeably undetectable in primary human T cells, which are also refractory toward IP agonist stimulation (Poloso, unpublished observations). Data demonstrate that inhibition of select cytokine and chemokines, which act to recruit other inflammatory cells to a site of injury or infection, are correlated with cytokines upregulated in the LPS-induced uveitis model we employed.
To date, prostaglandins delivered to the eye have been used as intraocular pressure (IOP)-lowering agents. 22 PGI2 mimetics have been used for pulmonary hypertension, but as of yet have not been translated into other diseases. We, thus, undertook to study the IP analog, cicaprost, in a model of acute LPS-induced uveitis. It is now accepted that tissue-resident macrophages are present in the iris, ciliary body, and some populations can be found in the uveal tract. 20 Additionally, DCs which were once thought absent have been shown to be important in ongoing immune responses in the eye and can be located not only to the cornea, but also in contact with retinal pigmented epithelial (RPE) cells in vivo. 21 Our data show an inhibition of both protein leakage and cell infiltration. The cytokine IL-6, as well as chemokines MCP-1 and RANTES, were lower after cicaprost treatment, consistent with the inhibition of inflammatory cells. This result was in line with what we have observed with prednisolone. The potential advantage of an IP agonist over steroids is multifold. Steroids administered to the eye can lead to complications (glaucoma [or elevation of IOP] and cataract formation). 27 Thus, they can only be administered for short periods of time. Data presented here suggest that IP agonists may prove to be an effective anti-inflammatory replacement for steroid treatment.
Finally, from the perspective of animal models, the highly efficacious effects of an IP agonist in uveitis may be regarded as surprising. IP receptor antagonists are widely described as effective in rodents of acute and chronic models of inflammation. Potent IP receptor antagonists were effective analgesics and anti-inflammatory drugs in carrageenan-induced paw edema and monoiodoacetate-induced osteoarthritis model.28,29 In a chronic model of inflammatory arthritis, an IP antagonist reduced the arthritis severity by 91%. 29 This sets ocular inflammation apart from other forms of inflammation in various tissues. This is supported by the observation that cicaprost was without nociceptive or other irritant side effects when applied to the eye in our studies. This further suggests the ophthalmic drug potential of IP agonist for treating inflammatory diseases of the eye.
Footnotes
Acknowledgments
This work was entirely supported by the intramural research program at Allergan, Inc. The authors thank D.F. Woodward for critical reading of this article.
Author Disclosure Statement
No competing financial interests exist.