
Abstract
Purpose:
Recent studies have shown the presence of SARS-CoV-2 entry factors on the ocular surface, identifying the eye as an additional entry route for the virus. Moreover, the coexpression of angiotensin-converting enzyme 2 (ACE2) with other SARS-CoV-2 entry factors [transmembrane protease serine 2 (TMPRSS2), transmembrane protease serine 4 (TMPRSS4), and dipeptidyl peptidase-4 (DPP4)] facilitates the virus infection.
Methods:
Here, we performed a study over 10 adult corneal and limbal tissues from human donors, both male and female between 58 and 85 years of age. Some of the main virus entry factors were analyzed and their expression was quantified and correlated with the age and sex of the donors through western blot. The receptors' localization was investigated through immunofluorescence.
Results:
Immunofluorescence confirmed the localization of ACE2 and TMPRSS2 on the ocular surface and showed, for the first time, the localization of TMPRSS4 and DPP4 in limbal and corneal epithelial superficial cells. The quantitative analysis showed that the expression of SARS-CoV-2 entry factors on corneal and limbal cells is likely to be modulated in an age-dependent manner, in agreement with the increased susceptibility to COVID-19 in the elderly. Moreover, we found a relationship between the expression of TMPRSS proteases with the activation state of limbal cells in 80-year-old donors.
Conclusion:
This study provides information on the expression of SARS-CoV-2 entry factors on the ocular surface of 10 adult human donors and is a first observation of a possible age-dependent modulation on corneal and limbal tissues. Our data pave the way to further investigate the susceptibility to the infection through the ocular surface in the elderly.
Introduction
The recent pandemic COVID-19, as named by the World Health Organization (WHO), a severe acute respiratory syndrome, is an increasing global problem caused by the new SARS-CoV-2 virus. 1 Since the beginning of the epidemic, many efforts have been made to expand knowledge on this virus pathogenesis and transmission routes. The identification of the SARS-CoV-2 entry factors represents a fundamental step forward for the study of SARS-CoV-2 infection. 2 Numerous recent reports have documented the ocular surface and eye involvement in patients with COVID-19. 3 In fact, the virus has been detected in tears and conjunctival secretions, suggesting that ocular exposure is a potential alternative route of infection through the nasolacrimal duct to the respiratory tract. 4
Notably, nasal epithelial cells, ocular surface cells, and bronchial secretory cells are known to express the key factors for cellular susceptibility to SARS-CoV-2.
Several studies indicated that the main susceptibility factor is the angiotensin-converting enzyme 2 (ACE2), a cell-surface receptor that binds the SARS-CoV-2 spike protein. Then, the transmembrane protease serine 2 (TMPRSS2), another known and important cell surface-associated protein, allows viral entry following binding of the viral spike protein to ACE2, thus enabling entry of SARS-CoV-2 into the host cells. Hence, the coexpression of ACE2 and TMPRSS2 allows a viral infection 100 times more efficiently than in the absence of TMPRSS2. 5
Furthermore, it has been demonstrated that the human ocular surface epithelium provides an additional entry route for SARS-CoV-2, which may exploit the inflammatory driven upregulation of ACE2 and TMPRSS2 entry factors to enhance the infection through a potential extrarespiratory transmission route.6,7
In addition, other coentry factors have been identified. For instance, the transmembrane protease serine 4 (TMPRSS4), another serine protease belonging to the TMPRSS family, is coexpressed with ACE2 in the eye. 7 Zang et al. reported that, besides TMPRSS2, TMPRSS4 also promotes infection of small intestinal enterocytes by SARS-CoV-2. 8 Dipeptidyl peptidase-4 (DPP4), also known as CD26, is a widely expressed membrane-anchored serine ectopeptidase, that can act as a virus-binding protein. 9 In fact, DPP4 serves as a viral receptor for Middle East Respiratory Syndrome Coronavirus of 2012 (MERS-CoV) 10 and docking molecular investigation highlighted the potential interaction of the S1 domain of SARS-CoV-2 spike glycoprotein with DPP4. 11
On this basis, the inhibition of DPP4 has been proposed as a potential therapeutic approach to combat the COVID-19 infection. 12 Furthermore, coexpression of ACE2 with TMPRSS2, TMPRSS4, and DPP4 on the ocular surface has been recently demonstrated. 7
Thanks to the collaboration with the Eye Bank of the “San Salvatore” Hospital of L'Aquila, which provided us the human ocular surface tissues from donors, we were able to carry out molecular studies that we believe will add important information to the already existing literature on the expression of entry factors for SARS-CoV-2 on the human ocular surface. The high value of this analysis comes from the relevant number and different ages of human samples that we were able to collect. This allowed us to show for the first time, in a study, age and sex related, the different expression levels of key SARS-CoV-2 entry factors (ACE-2, TMPRSS2, TMPRSS4, and DPP4) on the ocular surface.
Methods
Sample collection from human donors
Corneal/limbal/conjunctival samples (n = 10) from human donors were collected from the Eye Bank of San Salvatore Hospital (L'Aquila, Italy) and used with written consent. Demographic information about the human donors is reported in Table 1. All the tissues were preserved in tissue culture medium Tissue-c (Alchimia, Italy) at 31°C before use. One eye sample was used for protein quantification, and the other one for histological analysis.
Age and Sex of Human Donors
Western blotting
Adult cornea, limbus, and conjunctiva were carefully dissected under a stereomicroscope and frozen at −80°C until use. For protein extraction, the samples were finely dissected in a lysis buffer (50 mM Tris.Cl pH 7.8, 1% Triton-100, 0.1% SDS, 250 mM NaCl, 5 mM EDTA, 100 mM NaF, 2 mM NaPPi, 2 mM Na3VO4, 1 mM PMSF, 1 mM Aprotinin, 1 mM Pepstatin, and 1 mM Leupeptin) by using scalpel blades on Petri dishes kept on ice and then transferred to 1.5-mL microcentrifuge tubes. The samples were centrifuged, and the soluble phase was recovered and used for the subsequent analysis.
Protein concentration was quantified by using the Bradford assay (Bio-Rad Laboratories, Milan, Italy), and 30 μg of total protein extracts were run on a Bolt 4%–12% Bis-Tris Plus (Thermo Fisher Scientific, Waltham, MA) at 200 V for 20 min. PageRuler™ Plus Prestained Protein Ladder (#26619; Thermo Fisher Scientific, Waltham, MA) was used to identify the molecular weight of the bands. Due to the small amount of conjunctival tissue obtained from the donors during the ocular surface explants, the amount of protein extract was not sufficient to perform western blot analysis on this specific tissue.
For this reason, the western blotting (WB) analysis was performed only on corneal and limbal tissues (corneas from donors 1, 3, 4, 5, 6, 7, 8, and 10 and limbus from donors 1, 2, 4, 5, 6, 7, 8, and 9). The proteins were transferred to a polyvinylidene fluoride membrane (Millipore, Milan, Italy) through the iBlot 2 Dry Blotting System (Thermo Fisher Scientific, Waltham, MA). Membranes were blocked with 5% of blotting grade milk in Tris-buffered saline containing 0.1% Tween20 (TBST). The SARS-CoV-2 entry factors investigated in this study were selected on previous published data that already demonstrated their expression on the ocular surface.6,7 SARS-CoV-2 entry factors were detected using the primary antibodies reported in Table 2.
Antibodies Used for Quantification of SARS-CoV-2 Entry Factors
ACE2, angiotensin-converting enzyme 2; DPP4, dipeptidyl peptidase-4; TMPRSS2, transmembrane protease serine 2; TMPRSS4, transmembrane protease serine 4; MW, molecular weight.
Limbal activity was determined through the quantification of p27kip1 (anti-p27kip1 OriGene, #TA325320). Primary antibodies were diluted in 5% nonfat dry milk in TBST followed by secondary antibodies, anti-rabbit or anti-mouse (depending on the primary antibody) horseradish peroxidase-conjugated (Bio-Rad Laboratories) were diluted 1:2,000 in TBST containing 5% nonfat milk. The protein bands were detected using a Bio-Rad ChemiDoc XRS-plus imaging system (Bio-Rad Laboratories). Densitometric analysis was conducted by using the ImageJ software (U.S. National Institutes of Health, Bethesda, MD) and the amount of proteins was normalized versus α-actin or GAPDH.
For comparisons based on sex, the groups considered included males and females excluding the age factor. For comparisons based on age, the samples were divided in 3 groups: 60 years old (from 58 to 64 years), 70 years old (from 65 to 74 years), and 80 years old (over 75 years).
Cryosections
Adult ocular surface samples were incubated in 4% PFA overnight, followed by 3 washes in phosphate-buffered saline (PBS), incubated overnight in 30% sucrose in PBS and then embedded in Optical Cutting medium (OCT, Tissue Tek; Qiagen, Hilden, Germany) and stored at −20°C. Twenty micrometer cryosections were collected using a Leica cryostat on Superfrost Plus slides and stored at −20°C before use.
Immunofluorescence staining
Cryosections were used for immunostaining to visualize the SARS-CoV-2 entry factors and detect their localization. Five percent bovine serum albumin (BSA) was used to block nonspecific bindings. Sections were then incubated overnight at 4°C with the same primary antibodies used for WB (Table 2) diluted 1:250 in 1% BSA. Secondary antibodies were: anti-mouse IgG conjugated to green fluorescent dye (Alexa Fluor 488; Molecular Probes, Invitrogen, Carlsbad, CA) for anti-ACE2, anti-TMPRSS2, anti-TMPRSS4, and anti-rabbit IgG conjugated to green fluorescent dye (Alexa Fluor 488; Molecular Probes, Invitrogen) for anti-DPP4. Secondary antibodies were diluted 1:1,000 in PBS (1 × ) and incubated at 37°C for 2 h. The cryosections were counterstained with bisbenzimide (Hoechst) nuclear dye.
Confocal images were acquired using a Nikon 80i confocal microscope. The specificity of the immunofluorescence staining was assessed by omitting the primary antibodies (negative control) (Supplementary Fig. S1).
Statistical analysis
Student's t-test and linear regression analysis were used to perform the statistical analysis. First type error was set at 5%. The statistical analysis was performed by using the SigmaPlot 12.0 software.
Results
Localization of SARS-CoV-2 entry factors on cornea and limbus from adult human donors
Previous studies suggested that the ocular surface may serve as an entry route for SARS-CoV-2 due to the expression of its entry factors by several cell types of the eye.7,13 Corneal/limbal cryosections from adult human donors were used to detect and localize the SARS-CoV2 entry factors on the ocular surface (Fig. 1). Cornea and limbus tissues were identified by bisbenzimide nuclear staining (Fig. 1a). First, we confirmed the presence of ACE2 and TMPRSS2 in limbal and corneal cells (Fig. 1b–e). In addition, to the best of our knowledge, we found, the presence of additional SARS-CoV-2 entry factors, TMPRSS4, and DPP4, in limbal and corneal epithelial superficial cells through immunolocalization for the first time (Fig. 1f–i).

Representative confocal images of adult corneal/limbal cryosections immunostained for SARS-CoV-2 entry factors (green). The images show the presence of ACE2, TMPRSS2, TMPRSS4, and DPP4 in the superficial epithelium of cornea and limbus.
Quantitative analysis of ACE2 expression in cornea and limbus of adult human donors
ACE2 is the main SARS-CoV-2 receptor and its presence with other SARS-CoV-2 viral entry factors on the human ocular surface has already been identified.6,7 For the first time, in this study, we investigated their expression levels in correlation with the sex and age of the donors. The proteins were quantified in corneal and limbal tissues obtained from adult human donors.
Figure 2a and b shows the expression of ACE2 protein in both cornea and limbus region, respectively. In both tissues a greater expression of ACE2 was found in the female group, although the differences were not statistically significant.
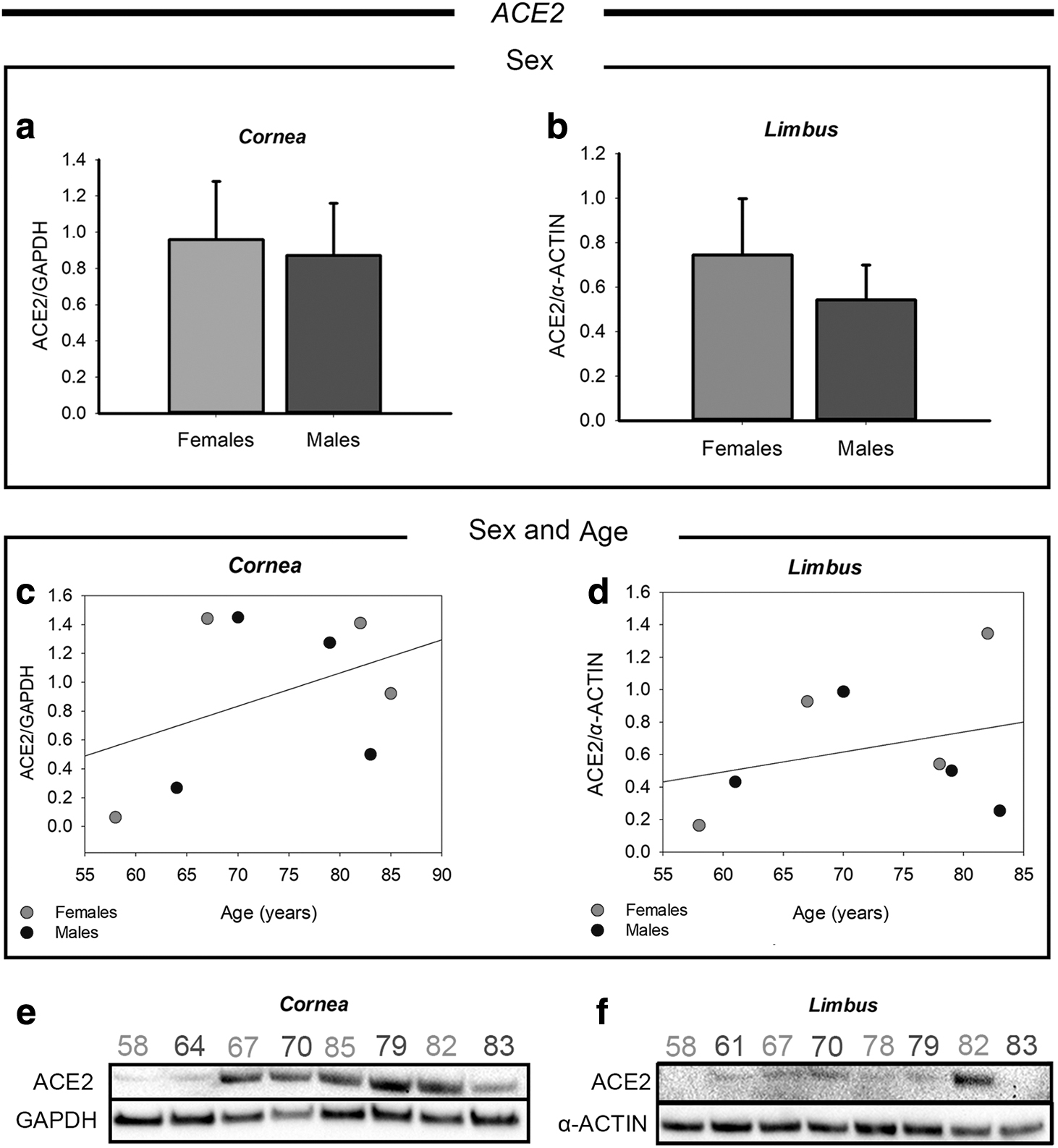
Western blot analysis of ACE2 expression in corneal and limbal tissues from adult human donors.
In Fig. 2c and d we reported scatter plot graphs showing the correlation between ACE2 protein levels and the age of the donors. Each dot represents one donor, and it is marked in dark grey or light grey for male or female, respectively. ACE2 expression in the cornea was found to be higher over 60 years of age both in males and females, as also showed by the slope of the regression line. This is also clearly visible from the western blot bands reported in Fig. 2e. Conversely, in the limbus, ACE2 protein levels did not correlate with age (Fig. 2d), since the donors showed similar expression levels, except of the 82-year-old female donor (donor 8), who presented a markedly increased ACE2 expression. The other age-matched donors showed consistent expression of ACE2 in the limbal region. Figure 2f shows the original western blot bands of ACE2 in the limbus.
Quantitative analysis of TMPRSS2 expression in cornea and limbus of human donors
As a second step, we performed a quantitative analysis of TMPRSS2 in the corneal and limbal regions. TMPRSS2 is the main SARS-CoV-2 entry factor, enabling increased susceptibility to the virus entry when it is coexpressed with ACE2 on the cell surface. TMPRSS2 protein quantification in correlation with sex demonstrated no difference between males and females both in the cornea and limbus (Fig. 3a, b). Conversely, an increase of expression was observed in relationship with age in the cornea in donors ≥67 years of age, as shown in the scatter plots of Fig. 3c and clearly appreciable from the western blot bands reported in Fig. 3e.
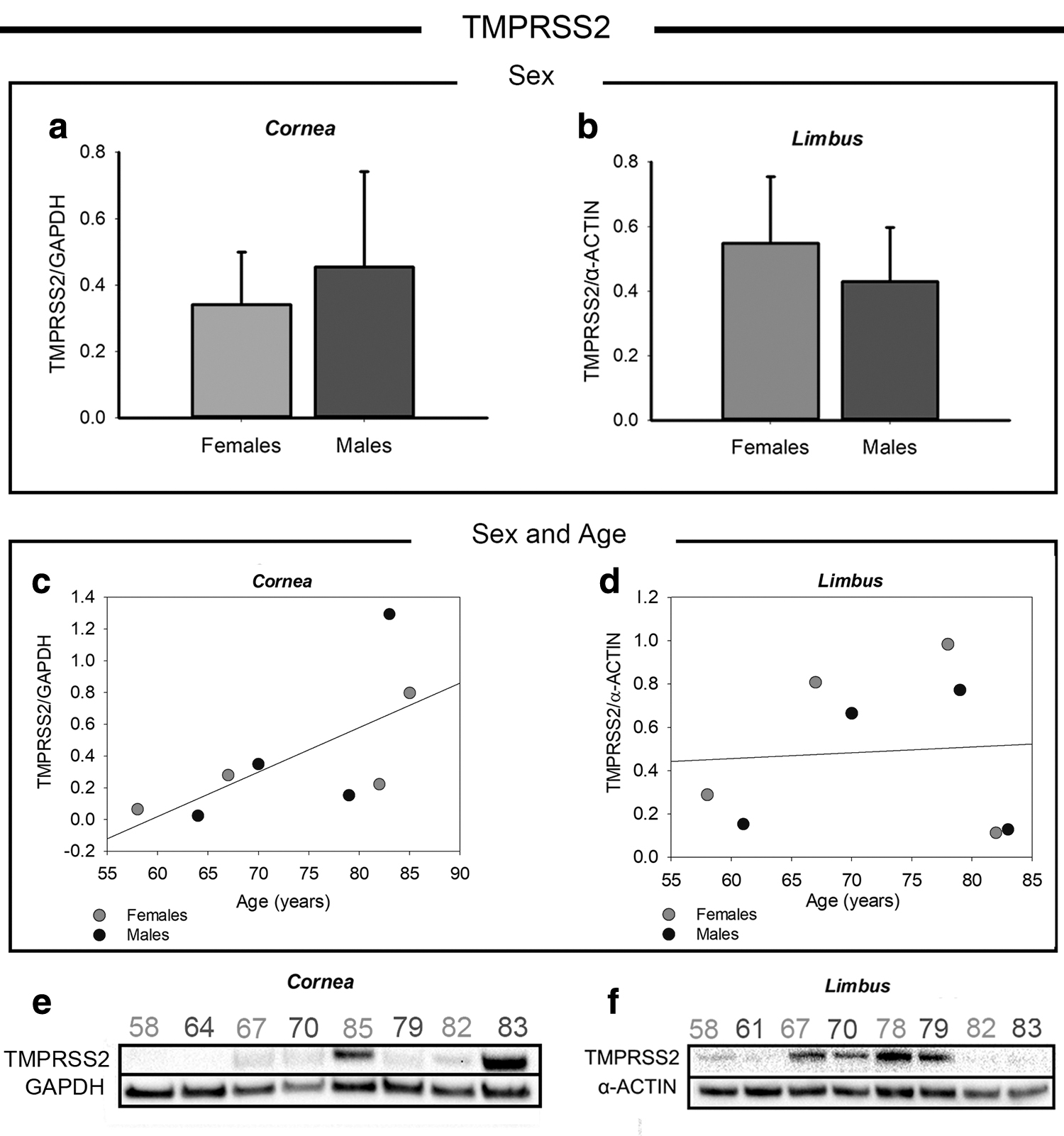
Western blot analysis of TMPRSS2 in corneal and limbal tissues from adult human donors.
The analysis of the samples from the oldest donors showed the highest TMPRSS2 expression in the cornea, whereas the other age-matched donors showed similar TMPRSS2 protein levels to those of the 70-year-old donors. In the limbus, a high variability was found between human donors without a clear correlation with sex or age (Fig. 3d), as clearly visible from the western blot bands of Fig. 3f. Intriguingly, the high variability of TMPRSS2 expression in the limbus may be associated with the activation state of limbal cells (reported in a dedicated paragraph below).
Quantitative analysis of TMPRSS4 expression in cornea and limbus of human donors
TMPRSS4, another TMPRSS family member of Transmembrane Serine Proteases, was also quantified and correlated with age and sex among the adult human cornea and limbus samples (Fig. 4). The antibody used for this experiment allowed us to identify both the basal and the glycosylated TMPRSS4 (Gly-TMPRSS4) forms. Hence, we performed a first analysis considering the total amount of the TMPRSS4 protease, and a second analysis considering the ratio between the Gly-TMPRSS4 and its basal form (Gly-TMPRSS4/TMPRSS4). Both the analysis did not show any difference between males and females either in the cornea (Fig. 4a, b) and in the limbus (Fig. 4c, d).
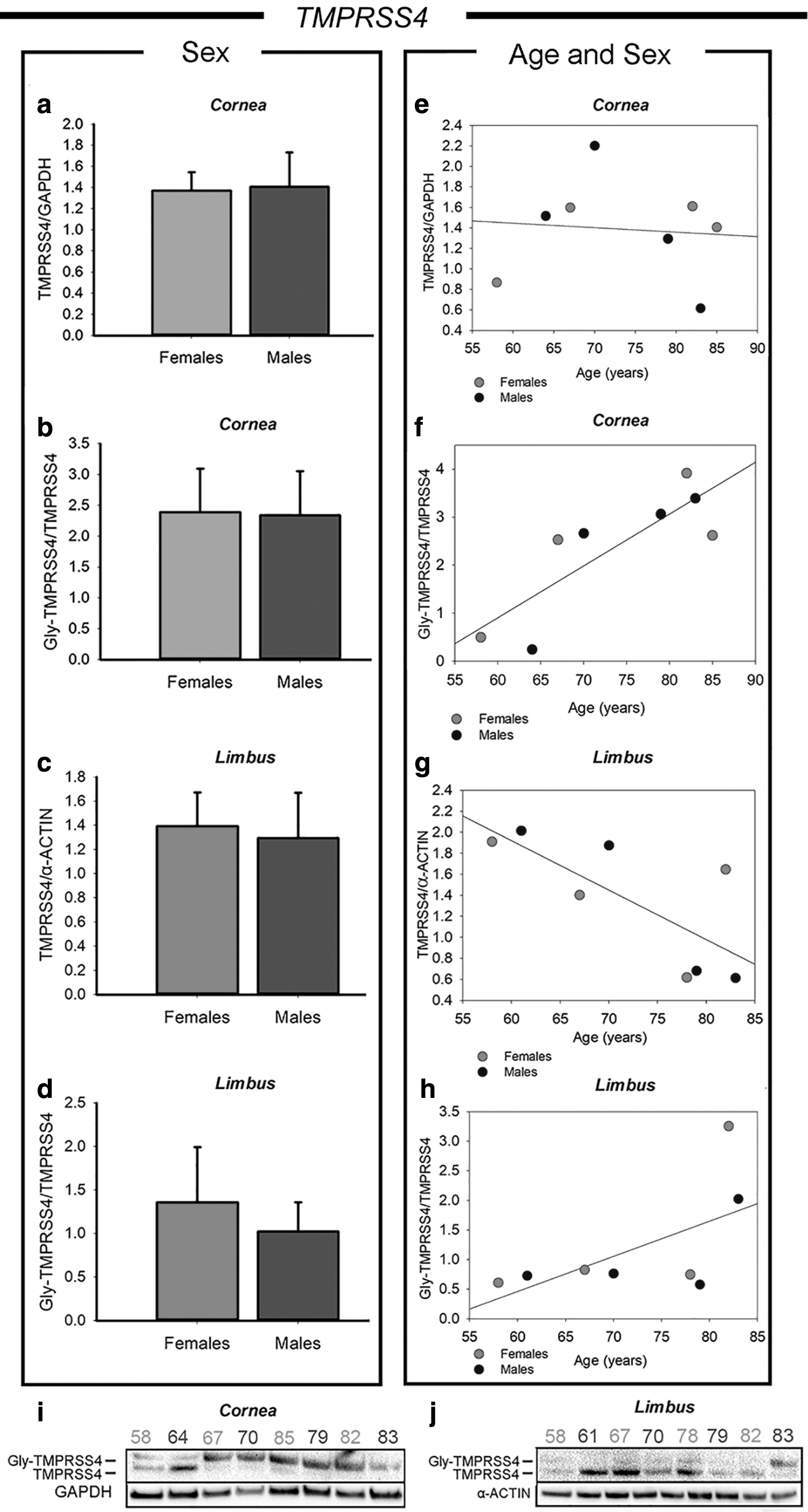
Western blot analysis of TMPRSS4 expression in corneal and limbal tissues from adult human donors. The protein amount was reported as total TMPRSS4 normalized versus the housekeeping (TMPRSS4/α-actin or GAPDH) and the ratio between glycosylated TMPRSS4 (Gly-TMPRSS4) and basal TMPRSS4 (Gly-TMPRSS4/TMPRSS4) after normalization versus the housekeeping.
The densitometric analysis based on age differences showed a more interesting result (Fig. 4e, f). In the cornea, the total amount of TMPRSS4 was not correlated with age differences among the human donors (Fig. 4e). However, the Gly-TMPRSS4/TMPRSS4 ratio showed a strong correlation with age (Fig. 4f), being the protein levels increased in the oldest donors compared with the younger ones. Moreover, regression analysis indicated that the correlation between Gly-TMPRSS4/TMPRSS4 with the donors' age is statistically significant (P = 0.011; R2 = 0.684). These data suggest that in the elderly there could be an increase of TMPRSS4 activity in the cornea, because of an increased glycosylation (Gly-TMPRSS4/TMPRSS4), rather than an increased expression of the TMPRSS4 itself. Although it is not specifically known whether this post-translational modification brings to a gain or loss of function of TMPRSS4.
In the limbus, the TMPRSS4 analysis (Fig. 4g, h) showed a different trend compared with the corneal region. In fact, the protein expression showed a decreasing trend from age 58 to 83 (Fig. 4g), which resulted to be statistically significant by linear regression analysis (P = 0.036; R2 = 0.545). Instead, the Gly-TMPRSS4/TMPRSS4 ratio (Fig. 4h) was very similar between all the donors, except of the oldest ones (ages 83 and 85), who showed increased levels of Gly-TMPRSS4/TMPRSS4. Figure 4i and l shows the original whole western blot bands of TMPRSS4 in the cornea and limbus regions, respectively.
Quantitative analysis of DPP4 expression in cornea and limbus of human donors
The last SARS-CoV-2 coentry factor analyzed in this study is the DPP factor 4 (DPP4), whose expression and its coexpression with ACE2 was already demonstrated in the ocular surface. 7 However, age- and sex-dependent differences of DPP4 expression have never been studied until now. As for the other analyzed markers, no significant differences were found between males and females both in the cornea and in the limbus (Fig. 5a, b).
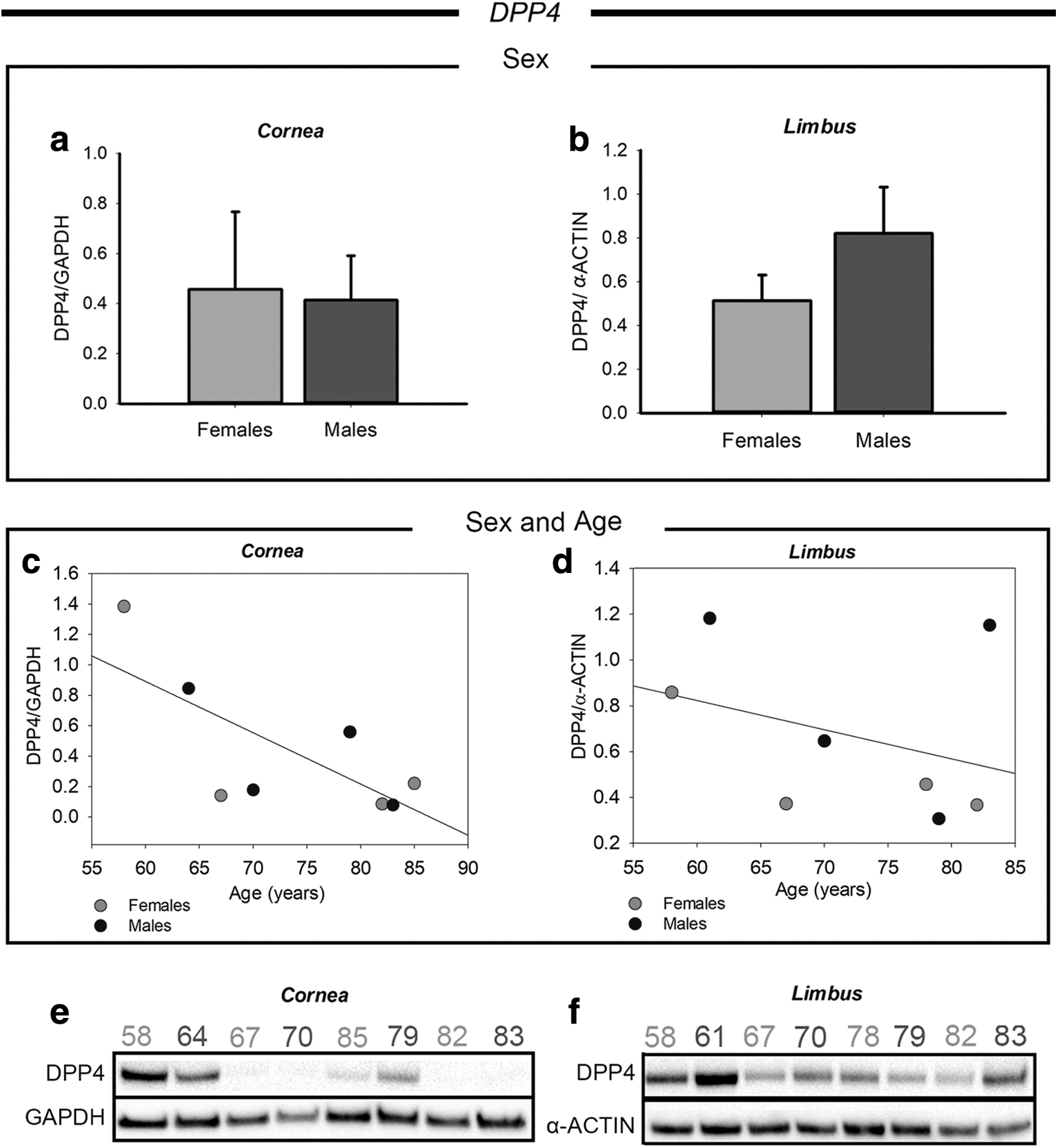
Western blot analysis of DPP4 in corneal and limbal tissues from adult human donors.
Differently, a correlation was found in relationship with the age of the donors in both the regions of the ocular surface, as reported in Fig. 5c and d. Particularly, the donors from 58 to 64 years of age showed a higher DPP4 expression compared with the older donors both in the cornea and in the limbus. From 67 to 85 years, the donors displayed a more consistent expression level of DPP4. Figure 5e and f shows the original whole western blot bands of DPP4 in cornea and limbus region, respectively.
Correlation between limbal activated/resting state and expression of SARS-Cov2 entry factors
Human corneal stem cells, lying in the basal layer of the limbus, are responsible for the renewal of the ocular surface epithelium. For this purpose, human limbal stem cells produce transit amplifying progenitors (activated state). When the limbal stem cells are quiescent (resting state) they express specific cell markers, such as the cyclin/Cdk inhibitor p27kip1, which enable cell cycle arrest. 14 In this study, we investigated the expression of p27kip1 in the limbus of adult human donors (Supplementary Fig. S2) to correlate the activation state of limbal stem cells 14 with the expression of SARS-CoV-2 entry factors (Table 3). The 58- and the 61-year-old donors showed a p27kip1 (−) phenotype, indicating an activated state, whereas the 67- and the 70-year-old donors showed a p27kip1 (+) expression, indicating a resting state. The 80-year-old limbal samples showed a different scenario.
Correlation Between p27kip1 and SARS-CoV-2 Entry Factors' Expression in Limbal Samples
The table shows the expression of p27kip1 (+ or −) correlated with ACE2, TMPRSS2, TMPRSS4, and DPP4 expression in the limbus of each donor. The original whole western blot bands of p27kip1 are shown in Supplementary Fig. S2. Further details about the donors are reported in Table 1.
In fact, donors 6 and 7 (78 and 79 years old, respectively) displayed a resting state (p27kip1 +), whereas donors 8 and 9 (82 and 83, respectively) showed an activated state (p27kip1 −). This difference was strongly associated with the expression of the TMPRSS proteases. p27kip1 (+) donors showed higher levels of TMPRSS2 expression and reduced Gly-TMPRSS4/TMPRSS4 ratio compared with the age-matched p27kip1 (−) donors. This important evidence could suggest that the activation state of limbal stem cells might influence the susceptibility to SARS-CoV-2 infection through the modulation of the TMPRSS protease expression.
No correlation between limbal activated/resting state and ACE2 or DPP4 expression was found.
Discussion
The ocular surface has been proposed to be an additional route of entry by SARS-CoV-2 due the presence of ocular manifestations in infected patients 3 as well as to the identification of the virus in the eye tears.4,15 Moreover, the expression of SARS-CoV-2 receptors has been identified in several ocular cell types.7,13 For instance, a previous immunohistochemical study on samples from 5 adult human donors demonstrated the expression of ACE2 and TMPRSS2 on the whole ocular surface (cornea, limbus, and conjunctiva). 16
Afterward, the coexpression of ACE2 and TMPRSS2 in the superficial conjunctival epithelium was also confirmed by Mencucci and coworkers through immunohistochemistry on 7 conjunctival samples from adult human donors. 17 Finally, the single cell analysis on corneal, limbal, and conjunctival cells derived from 4 human donors provided a wider overview over the expression of the main SARS-CoV-2 entry factors in adults. 7
The present study provides a descriptive analysis on the expression of the entry factors for SARS-CoV-2 on the human ocular surface from 10 adult human donors, implementing the information of the existing literature, which is restricted to small sample size, probably due to the difficulty in the retrieval of eyes from human donors. Moreover, for the first time in this study, the expression of the SARS-CoV-2 entry factors on corneal and limbal tissues was investigated in relationship with sex and age of the donors.
The main finding of our study suggests that the expression of SARS-CoV-2 entry factors on the ocular surface is likely to be modulated in an age-dependent manner, in agreement with the increased susceptibility to COVID-19 in the elderly observed during the pandemic. 18 In particular, we analyzed the protein levels of ACE2, TMPRSS2, TMPRSS4, and DPP4 in corneal and limbal tissues from adult human donors from 58 to 85 years of age. Although the sampling did not comprise younger donors, we found notable and interesting differences from 60 to 80-year-old donors, and we can speculate that greater differences could be found in comparison with younger people.
Particularly notable is the general increase of ACE2 expression observed after 60 years of age, which is more evident in the cornea region. ACE2 is the main SARS-CoV-2 entry factor through the binding to the S protein on the virus surface. 2 Hence, ACE2 increase in the elderly, as found in this study, suggests a higher susceptibility to SARS-CoV-2 infection through the ocular surface with aging. Notably, an age-dependent modulation of SARS-CoV-2 entry factor's expression was observed in other tissues that are more directly involved in SARS-CoV-2 infection, such as the nasal, 19 oral, 20 and lung epithelia. 21 Moreover, it was shown that the presence of comorbidity, a common condition observed in the old population, increases the expression of ACE2 and TMPRSS2. 22
In addition, we also investigated the expression of other SARS-CoV-2 coreceptors involved in the infection pathway. In fact, TMPRSS2 and TMPRSS4, belonging to the TMPRSS family of proteases, have been shown to facilitate the virus infection process.5,8 Both the proteases showed a general increasing expression trend with aging either in corneal and limbal tissues, except for the total amount of TMPRSS4, which resulted to be reduced with aging. Intriguingly, we found a strong relationship between the expression of those factors with the activation state of limbal cells in 80-year-old donors.
Although we can speculate that this correlation could be related with the limbal activated/resting state, future studies that will include other limbal niche marker, such as isoforms β and γ of the ΔNp63, 23 ABCG2, ABCB5, 24 C/EBPσ, Bmi1, and Notch-1, 25 are needed to corroborate our evidence. Nevertheless, the results presented herein find additional support from the recent published literature as TMPRSS family members, among which TMPRSS4, were found in the ocular surface, including the limbus region and the quiescent limbal stem cells as well. 7
Taken together, these evidences suggest that the limbus activation state could influence the expression of such receptors to enhance infection in the ocular surface. Furthermore, we found a significant increase of TMPRSS4 glycosylated form compared with the basal form after age 60 in the cornea. Although, so far, no studies have investigated the functional activity of the glycosylated TMPRSS4 form, it is known that post-translational modifications like glycosylation may influence the catalytic activity of proteases. 26 Moreover, it is known that glycosylated proteins are mainly present on the cell surface, and therefore the increase of Gly-TMPRSS4 in the eldest donors could in turn induce an increased susceptibility to the infection.
Hence, TMPRSS4 could be more active and effective in prompting the body entrance of the SARS-CoV-2 virus through the ocular surface.
The last coentry factor analyzed in this study is the DPP4, whose relevance as SARS-CoV-2 entry factor has been previously suggested in relationship to docking molecular studies, 11 to the coexpression with ACE2 in several cell types, including ocular surface cells, 6,7,13 and to the demonstrated role as receptor for other coronaviruses (i.e., MERS–CoV). 12 In this study, we show that DPP4 is differently expressed in cornea and limbus region from age 60 to 80. Interestingly, DPP4 tended to decrease with aging, differently from the other entry factors investigated in this study.
In addition to the age of the donors, their sex was also taken into account to evaluate a possible sex-related expression of the SARS-CoV-2 entry factors in the eye. Indeed, epidemiological data have already shown that males are more susceptible to SARS-CoV-2 infection and showed a prolonged viral RNA shedding compared with the females. 18 Also, sexual differences have been highlighted for the main SARS-CoV-2 receptors expression.22,27,28 Other studies have proposed that COVID-19 could also be related to the androgen sensitivity.29,30 Accordingly, patients with prostate cancer showed an increased risk of COVID-19, whereas patients undergoing androgen-deprivation therapies, due to the prostate cancer, showed a reduced risk. 31
Moreover, these evidences suggest that the activity of the androgen receptor could represent another major feature in driving the SARS-CoV-2 infection. However, we did not find any correlations between the sex and the expression of the SARS-CoV-2 entry factors in the cornea and limbus. The high variability observed was clearly associated with differences in the expression of the factors depending on their age, suggesting that on the ocular surface, the expression of the SARS-CoV-2 entry factors is more susceptible to age than sex. Nevertheless, further studies on a larger sample size are needed to better investigate whether sex differences existed in age-matched donors.
Conclusion
Taken together, our results confirm the presence of the SARS-CoV-2 entry factors on the ocular surface and further enhance the evidence that the eye may serve as an additional route of entry for the virus. Our study was conducted on a sample size of 10 adult human donors from 58 to 85 years of age. Further studies will allow one to expand the knowledge also to a wider aging range.
Nevertheless, this is the first evidence of an age-dependent modulation of SARS-CoV-2 entry factors on the ocular surface and it represents an important feature to be considered about the increased susceptibility to COVID-19 infection in the elderly. This is particularly important especially on the basis of new SARS-CoV-2 variants' occurrence. For instance, the P.1, B.1.351, and B.1.1.7 variants (from Brazil, South Africa, and United Kingdom, respectively), display mutations that modify the virus S protein and impact on its binding affinity with the ACE2.32–35 On this basis, the presence of a wide range of SARS-CoV-2 entry factors on the ocular surface is even a more important feature to be considered and calls for the need of ocular protective devices as a way to limit the virus spread.
Footnotes
Authorship Confirmation Statement
All persons who meet authorship criteria are listed as authors.
Acknowledgment
The authors would like to thank Dr. Giulia Carozza for technical support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research received no specific funding.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.