
Abstract
Purpose:
This study aims to compare the cytotoxic, apoptotic, and oxidative effects of preserved and preservative-free forms of brimonidine 0.15% on the human corneal epithelial cell (HCEC) line.
Methods:
Time-dependent cytotoxicity studies were performed with the Alamar Blue method. For apoptotic studies, PE Annexin V and 7-amino-actinomycin (7-AAD) staining and flow cytometry were performed. Messenger RNA (mRNA) expressions of Bax, Bcl-2, and caspase-3, -9, -12, and protein expressions of Bax and Bcl-2 were evaluated by quantitative real-time polymerase chain reaction and Western blot method, respectively.
Results:
Cell viability was 76.4% with the preserved solution and 36.05% with the preservative-free solution at the fifth minute. No significant difference was observed with either solution at the 15-min mark, whereas cell viability did not change significantly after 1 h. In the apoptosis evaluation, it was observed that the preservative-free solution increased the early apoptotic activity to a greater degree (P < 0.05). Preservative-free solution also induced gene expression of proapoptotic Bax, caspase-9 and -12, and protein expression of Bax while reducing the protein expression of anti-apoptotic Bcl-2 (P < 0.0001). Preserved solution induced only the gene expression of caspase-12, and reduced the protein expression of Bcl-2 (P < 0.0001). No significant difference was observed in the reactive oxygen species (ROS) levels of either solution compared with the control group (P > 0.05).
Conclusion:
It was demonstrated that the preserved solution is less cytotoxic to the HCEC line in the early period, has less early apoptotic activity, and does not significantly increase ROS levels.
Introduction
Glaucoma is a chronic and multifactorial optic neuropathy characterized by progressive optic nerve damage and the second most common cause of vision loss worldwide after cataracts. 1 Although factors such as age, family history, and race contribute to the development of glaucoma, high intraocular pressure (IOP) is recognized as the greatest risk. Today, all drugs used for the medical treatment of glaucoma aim to prevent optic nerve damage by lowering IOP. Brimonidine [5-bromo-6-(2-imidazolidinylideneamino) quinoxaline] is a highly selective alpha 2-adrenergic receptor agonist that lowers IOP by reducing aqueous humor inflow and increasing uveoscleral outflow. It is frequently preferred for both short-term and long-term treatments after laser procedures because of its advantages such as effective aqueous suppression, quick onset of action, and rapid elimination. 2
Although generic brimonidine tartrate was produced in 0.2% and 0.15% concentrations and contains benzalkonium chloride (BAK), a newer, better tolerated form containing 0.005% Purite® has been introduced (Alphagan P; Allergan). 3 This was followed by preservative-free brimonidine 0.15% preparations using PureFlow® technology in the following years (Brimogut; Bilim Ilac). An important goal in formulating topical ophthalmic glaucoma medications was to maintain effective IOP reduction through effective drug delivery while enhancing safety and tolerability. 4
Preservatives constitute an important component of ophthalmic solutions in terms of deterring microbial growth in the bottle and preventing the decomposition of the active drug. 5 BAK has become most commonly used preservative in ophthalmic solutions. 6 Owing to the known toxic effects of BAK on the ocular surface, the pursuit for novel preservative molecules that minimize toxicity continues. Molecules such as stabilized oxychloro complex (SOC; Purite), sofZia®, polyquad, sodium perborate, and sorbic acid are among the preservatives commercially used for this purpose. 7 Purite is an SOC and has long been used in water purification systems (sodium chloride). 8 SOC assumes its preservative effects from its oxidative properties. 9 In recent years, the manufacture of multidose preservative-free bottles with designs that prevent the entry of microorganisms into the bottle has become an increasing trend.
However, the literature has shown that preservative-free brimonidine yields more symptoms of irritation in the early period of application compared with its preserved form. 7 As we have observed a similar situation in our clinical practice, in this study, we aimed to examine and compare the cytotoxic, apoptotic, and oxidative effects of 2 different forms of brimonidine on human corneal epithelial cells (HCECs).
Methods
Eye drops investigated in experiments
To compare cytotoxic, apoptotic, and oxidative effects of preserved and preservative-free forms of brimonidine 0.15% on the HCEC line, Alphagan P, Allergan (preserved form), and Brimogut, Bilim Ilac (preservative-free form) were used as their commercial presentation. Although both preparations have the same molecule (brimonidine tartrate 0.15%) as the active ingredient; Alphagan P contains 0.005% Purite as preservative agent, whereas Brimogut uses PureFlow bottle technology to avoid contamination. Each milliliter of Alphagan P contains the inactive ingredients sodium carboxymethylcellulose, sodium borate, boric acid, sodium chloride, potassium chloride, calcium chloride, magnesium chloride, hydrochloric acid, and/or sodium hydroxide to adjust pH and purified water. The inactive ingredients in Brimogut are as follows: polyvinyl alcohol, sodium citrate dihydrate, citric acid monohydrate, sodium chloride, disodium edetate, hydrochloric acid, and/or sodium hydroxide to adjust pH and purified water.
In vitro human cell culture studies
Growth of HCECs
Human corneal epithelial cells (HCECs; PCS-700-010) were obtained from The American Type Culture Collection (ATCC, Rockville, MD). Cells were incubated in Dulbecco's modified Eagle's medium (DMEM) high glucose medium containing 10% fetal bovine serum, 2% penicillin–streptomycin, and 1 mM
Cytotoxicity studies
Time-dependent cytotoxicity studies
To determine the time-dependent cytotoxic effect of commercially available preserved and preservative-free brimonidine 0.15% preparations on HCEC, cells were seeded in 96-well plates at 104 cells per well. Viability of cells incubated for 5 min, 15 min, 1 h, 6 h, and 24 h with brimonidine and brimonidine–purite was determined by spectrophotometer (MultiScanGO) using Alamar Blue (Invitrogen, Thermo Fisher Scientific, Waltham, MA). For this, a single clinical dose of both chemicals was delivered (50 μL) via pipette tips. This concentration was equivalent to the use of drops on the clinical ocular surface and a total volume of 200 μm was completed with DMEM high glucose. Three replications were made for this experiment.
Evaluating the effects of preserved and preservative-free brimonidine 0.15% preparations on cell apoptosis and necrosis with flow cytometric methods
Flow cytometry assay was performed to determine the apoptosis and necrosis rates of preserved and preservative-free brimonidine 0.15% solutions on HCECs. For this, HCECs were first seeded in 6-well plates and preserved and preservative-free brimonidine 0.15% solutions added and incubated for 15 min at 5% CO2, 37°C.
At the end of the incubation, the cells were taken into clean Eppendorf tubes and centrifuged at 1,500g for 5 min. Commercial PE Annexin V Apoptosis Detection Kit I (ABD, Cat. No. 559763; BD Pharmingen, San Diego, CA) was used for flow cytometry analysis. Cell death caused by apoptotic or necrotic processes was investigated by staining with PE Annexin V and 7-amino-actinomycin (7-AAD) dyes included in the kit. The pellet was dissolved with 100 μL of 1 × binding buffer and stained with PE Annexin V and 7-AAD dyes. About 400 μL of 1 × binding buffer was added to the cells incubated for 20 min in the dark and analyzed with the NovoCyte Flow Cytometer System (Acea). Thus, early/late apoptosis and necrosis rates were determined.
Detection of reactive oxygen
Reactive oxygen species (ROS) are natural byproducts of normal oxygen metabolism and play important roles in cell signaling. ROS levels can increase significantly in conditions related to oxidative stress. The Cellular ROS Assay Kit (ab186029) used in the study uses an ROS sensor to measure ROS in living cells. The dye included in the kit is cell permeable and produces red fluorescence when it reacts with ROS. This fluorescent signal can be measured by fluorescence microscopy, flow cytometry, and microplate fluorometry. For this, cells were first cultured in 96-well plates with 104 cells/90 μL per well. Cells reaching sufficient density were treated with preserved and preservative-free 0.15% brimonidine and incubated at 5% CO2, 37°C. After 15 min of incubation, 10 × test buffer prepared in phosphate-buffered saline (PBS) was added to the cells and ROS production of the cells was stimulated for 15 min. ROS was determined using a fluorescent spectrophotometer at Ex/Em = 650/675 nm.
Gene expression studies
Quantitative real-time polymerase chain reaction (qRT-PCR) studies were performed to determine the effects of preserved and preservative-free 0.15% brimonidine preparations on messenger RNA (mRNA) expressions on HCECs. HCECs were treated with preserved and preservative-free 0.15% brimonidine preparations. Total RNA was isolated by the Trizol method. 10 The amount and purity of the RNA obtained were measured with the Bioanalyzer, and the amount of RNA to be used in complementary DNA (cDNA) synthesis was calculated. cDNAs were synthesized with the iScript™ cDNA synthesis kit. qRT-PCR was performed using 2 μL cDNA, 7.2 μL RNAase-free water, 0.4 μL forward primer, 0.4 μL reverse primer, 10 μL SYBR green, and the CFX-Connect Real-Time PCR system (Bio-Rad). Then, for the reaction mixture of the PCR process, the first activation was performed at 95°C for 30 s, denaturation was carried out at 95°C for 5 s and at 56°C for 30 s, and the process was terminated in a total of 40 cycles.
Changes in Bax, Bcl-2, Caspase-3, Caspase-9, and Caspase-12 gene expressions were determined by calculating the ratio of specific gene mRNA expressions to housekeeping genes (GAPDH) mRNA expression. The primers used were designed using Primer 3 software and their specificity was checked with NCBI (Table 1).
Primer Sequencing for Selected Genes
Protein expression studies
Western blot studies were performed to determine the effects of preserved and preservative-free 0.15% brimonidine preparations on GADPH (60004-1-Ig), Bax (50599-2-Ig), and Bcl-2 (ab182858) expressions. For this, cells were first grown in 100 mm Petri dishes, treated with preserved and preservative-free 0.15% brimonidine, and incubated at 37°C for 15 min. After incubation, cells were removed mechanically with cold PBS using a cell scraper. Cells were taken into clean Eppendorf tubes and were centrifuged at 4,000g at +4°C for 5 min. The pellet was resuspended by adding Ripa buffer (10 × ) + ddH2O + PMSF mixture. The cells were centrifuged again at 14,000g at +4°C for 10 min and the protein quantification was determined by making measurements at 562 nm wavelength according to the BCA method using the bovine serum albumin Standard. 11 Subsequently, their effects on the expression of GAPDH, Bax, and Bcl-2 proteins involved in the apoptosis pathway were determined by the Western blot technique described by Laemmli. 12
According to this method, proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) by applying vertical electrophoresis and specific antibodies were used to determine the expression of each protein.
Statistical analysis
Statistical analyses of the findings of the studies were conducted using the GraphPad Prism version 8.00 (GraphPad Software, San Diego, CA) program. The mean ± standard deviation values of all results were analyzed. The mean values were calculated using the Student's t-test (*P < 0.05, **P < 0.001, ***P < 0.0001).
Results
Cytotoxicity studies
Time-dependent cytotoxicity studies
Five minutes into the test, the ratio of the viable cells was 76.4% with the preserved preparation and 36.05% with the preservative-free preparation. When all the results were examined, it was observed that >50% of the cells died by the 15th minute, and cell viability did not change significantly after 60 min (Fig. 1).
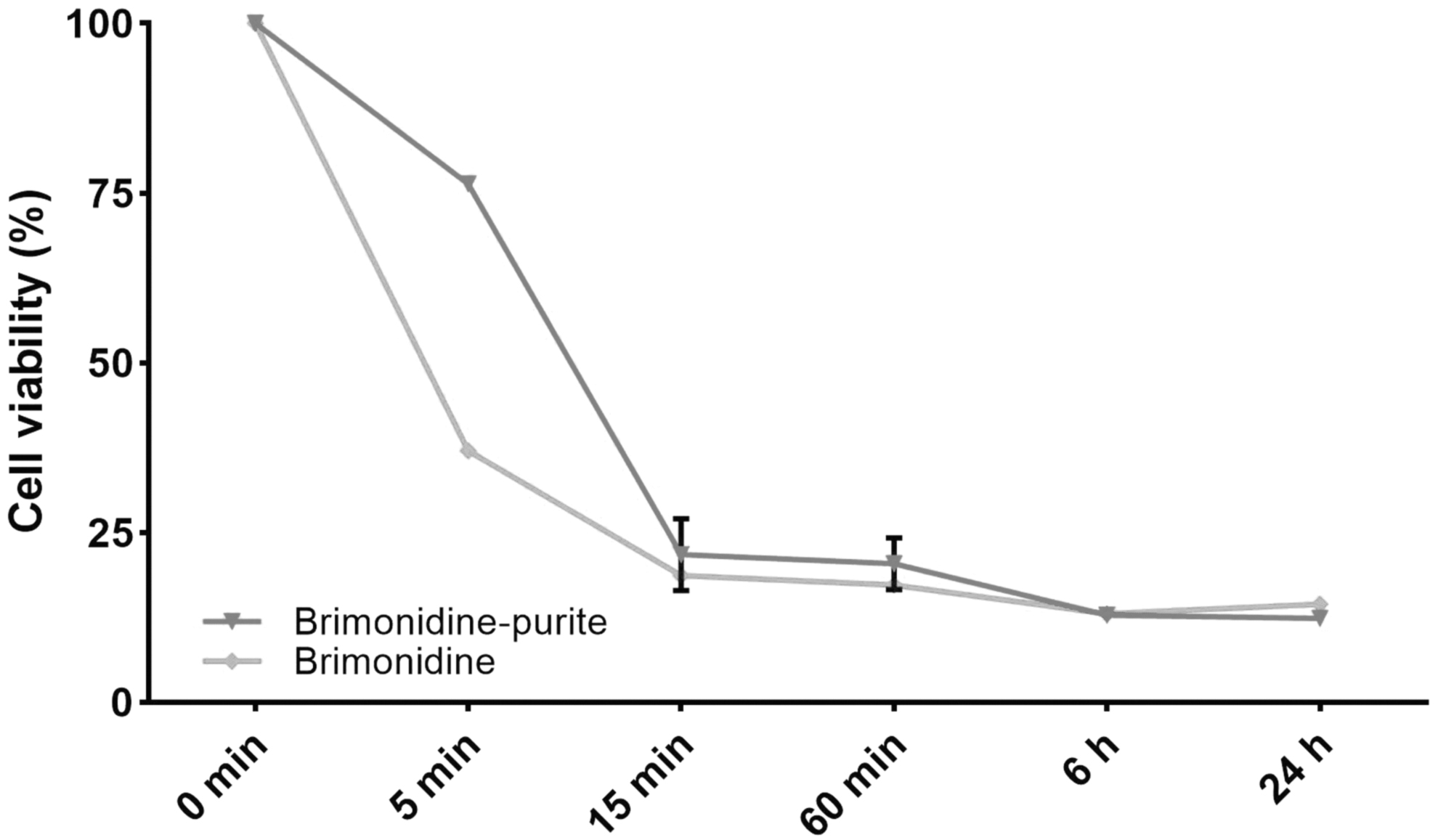
Cytotoxic effects of the preserved and preservative-free brimonidine 0.15% preparation on immortalized HCEC line; HCEC. The time-dependent toxicity of the preparations was followed for 24 h. All experiments were repeated 3 times. The results were presented as mean ± SD, n = 4. HCEC, human corneal epithelial cell; SD, standard deviation.
Investigation of the effects of preserved and preservative-free brimonidine 0.15% preparations on cell apoptosis and necrosis with flow cytometric methods
When apoptotic and necrotic effects were evaluated with flow cytometry, it was found that 9.4% of HCECs treated with preserved preparation underwent early apoptosis, 7.2% underwent late apoptosis, and 1.4% underwent necrosis. Thus, preserved brimonidine 0.15% induced 1.32-fold more early apoptosis in HCECs than control cells (P < 0.05) (Figs. 2 and 3). It was found that 20.7% of HCECs treated with preservative-free brimonidine underwent early apoptosis, 7.6% late apoptosis, and 1.2% necrosis. Thus, preservative-free brimonidine drove HCECs to early apoptosis 2.91-fold more than control cells (P < 0.05) (Figs. 2 and 3).
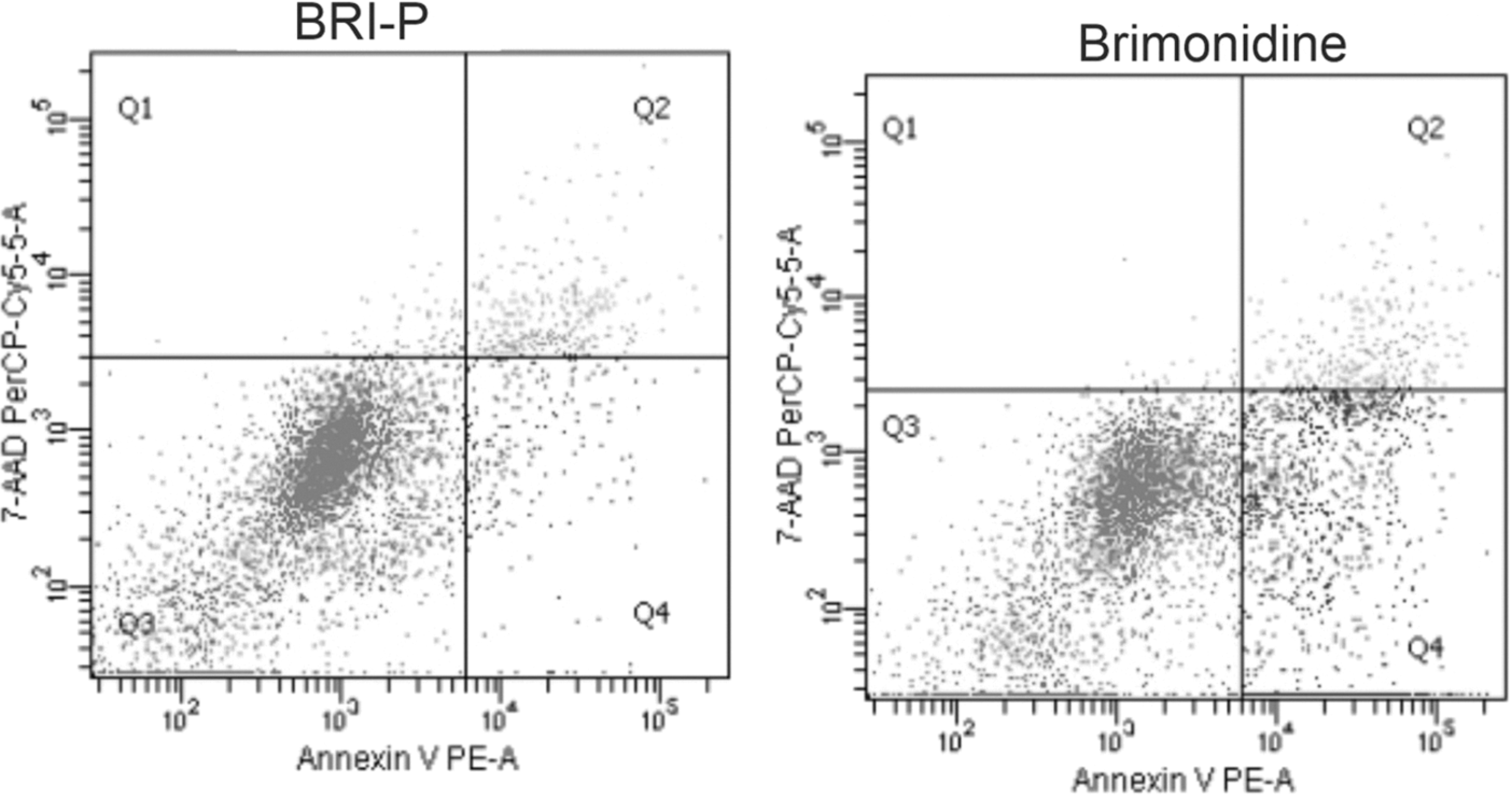
The effects of preserved and preservative-free 0.15% brimonidine preparations on programmed cell death (apoptosis). The cells were examined by flow cytometry following PE Annexin V-APC and 7-AAD staining; Q1: necrotic zone (PE Annexin V−/7-AAD+); Q2: late apoptotic zone (PE Annexin V+/7-AAD+); Q3: viable cells (PE Annexin V−/7-AAD−); Q4: early apoptotic zone (PE Annexin V+/7-AAD−). 7-AAD, 7-amino-actinomycin.

Cumulative graph of necrosis, early apoptosis, and late apoptosis rates compared with the control group of preserved and preservative-free 0.15% brimonidine preparations. All experiments were repeated 3 times. The results were presented as mean ± SD, n = 4 (*P < 0.05, **P < 0.001).
Reactive oxygen species
As seen in Fig. 4, increased ROS levels were observed in the cells treated with the preparations compared with the negative control group, and decreased levels compared with the positive control group (P > 0.05). ROS increased 1.1-fold in cells treated with preserved brimonidine and no increase was observed in cells treated with preservative-free brimonidine (P > 0.05). The results for both of the preparations were statistically insignificant.
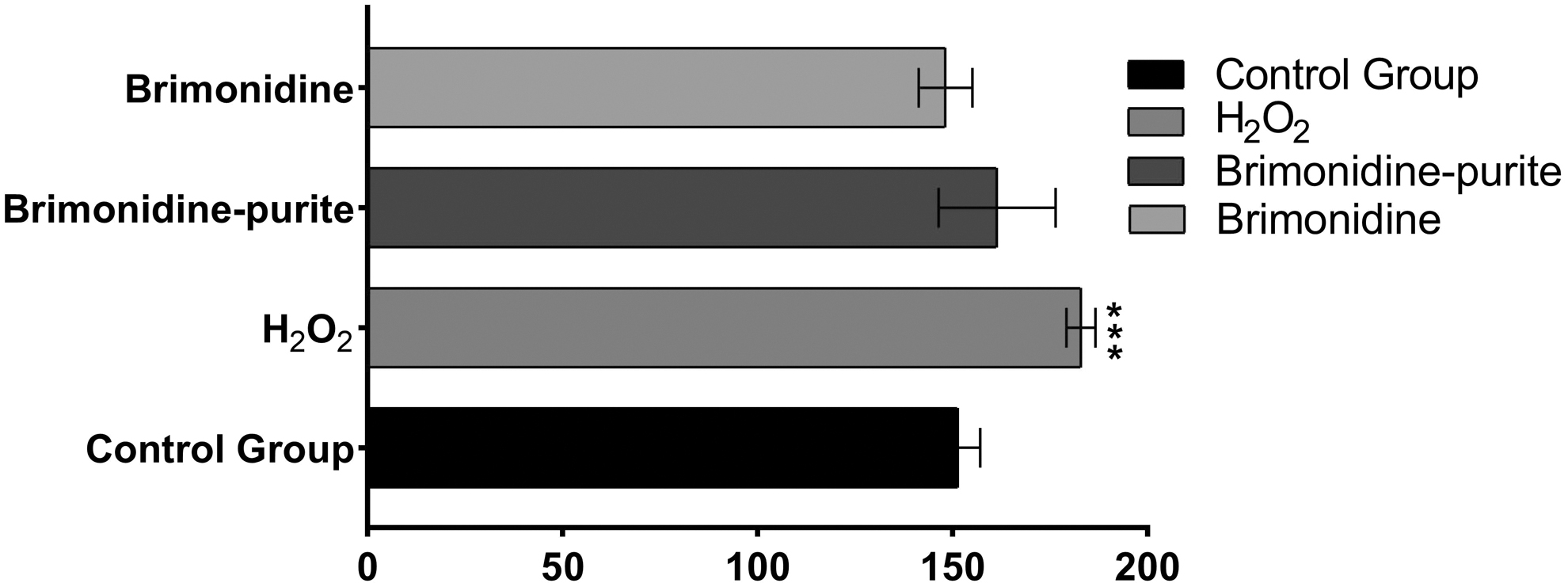
Reactive oxygen levels in HCECs treated with preserved and preservative-free brimonidine 0.15% preparations. Increased ROS levels were observed in cells treated with brimonidine and brimonidine–purite compared with the negative control group, and decreased ROS levels compared with the positive control group. All experiments were repeated 3 times. The results were presented as mean ± SD, n = 4 (***P < 0.0001). ROS, reactive oxygen species.
Gene expression studies
Compared with the control group, the preserved preparation decreased Bax gene expression to 0.62 of the control group (P > 0.05), whereas the preservative-free preparation increased it 1.88-fold (P < 0.0001). Preserved brimonidine increased gene expression of Bcl-2 by 1.34-fold (P < 0.001), whereas preservative-free brimonidine decreased it to 0.68-fold (P > 0.05). Preservative-free brimonidine increased gene expression of Caspase-3 by 1.64-fold (P > 0.05), whereas preserved brimonidine decreased it to 0.18-fold (P < 0.001). Preservative-free brimonidine increased gene expression of Caspase-9 by 2.40-fold (P < 0.0001), whereas preserved brimonidine decreased it to 0.51-fold (P < 0001). Preserved brimonidine increased gene expression of Caspase-12 by 2.97-fold (P < 0.0001), whereas preservative-free brimonidine increased it by 2.36-fold (P < 0.0001) (Fig. 5).
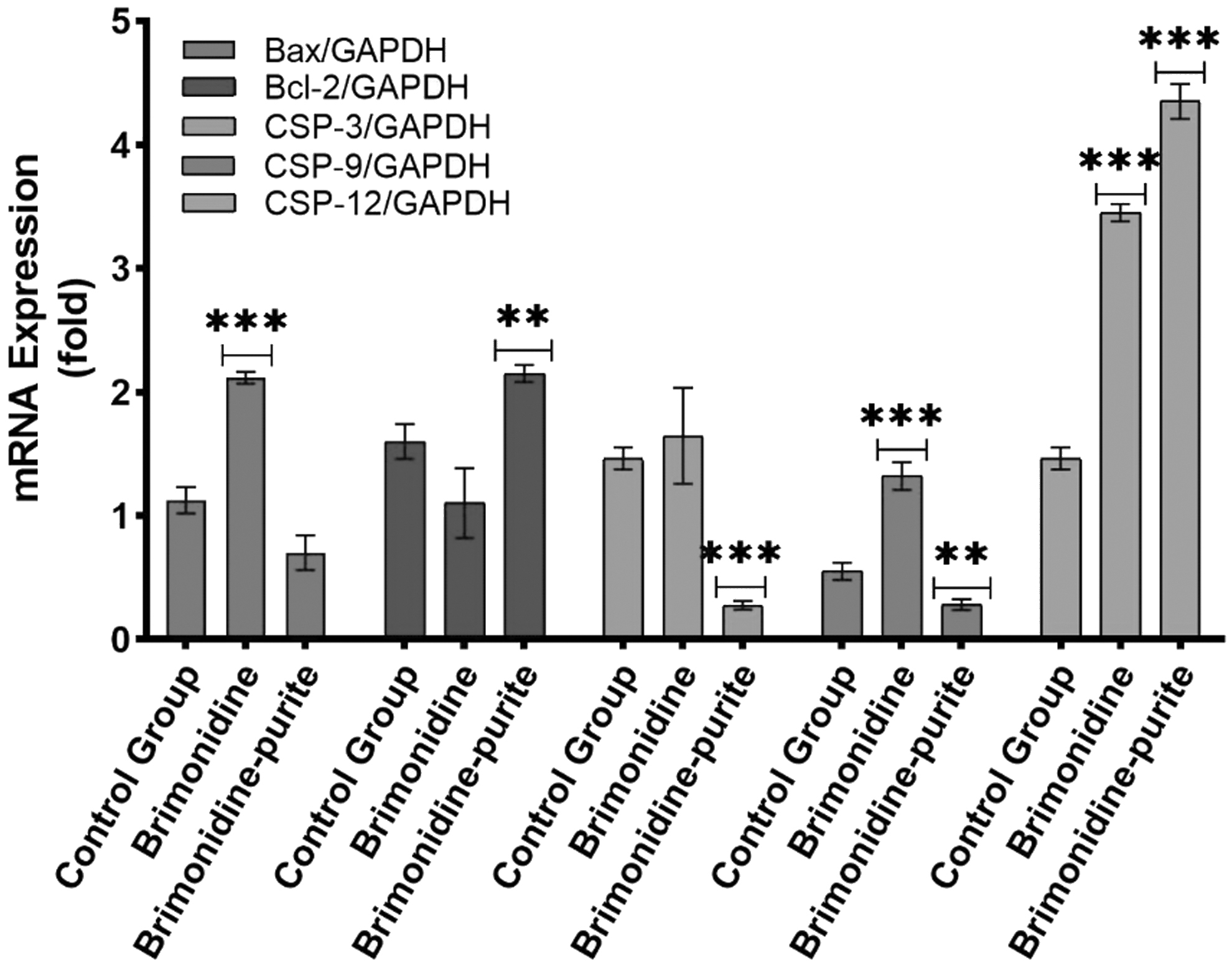
Effect of preserved and preservative-free 0.15% brimonidine on mRNA expression of Bax, Bcl-2, Caspase-3, -9, and -12 gene. All experiments were repeated 3 times. The results were presented as mean ± SD, n = 4 (**P < 0.001, ***P < 0.0001). mRNA, messenger RNA.
Among investigated genes, Bax and Bcl-2 families encode proteins that play a key role in the regulation of apoptosis and show pro- and antiapoptotic effects, respectively. In addition, caspase-3 and -9 were evaluated with the aim of examining the effector and initiator caspase subgroups, respectively. Besides, caspase-12 was investigated, which is associated with the endoplasmic reticulum and triggered by inflammatory processes. These results show that the preservative-free preparation significantly increases the proapoptotic components in the mitochondria-dependent apoptotic pathway compared with brimonidine–purite. However, the expression of the Caspase-12 gene, which is involved in the mitochondria-independent apoptotic pathway, was induced by both preparations, with brimonidine–purite being more potent.
Protein expression studies
According to analyses of the intensities of the immunoreactive protein bands (Fig. 6) the preserved preparation increased Bax protein expression 1.01-fold (P > 0.05), whereas the preservative-free preparation increased it 2.48-fold (P < 0.0001). Bcl-2 protein expression was decreased 0.55-fold with preserved brimonidine (P < 0.0001) and 0.57-fold with preservative-free brimonidine. (P < 0.0001) (Fig. 7). It was observed that both solutions suppressed the antiapoptotic Bcl-2 protein expression; however, the preservative-free solution increased the proapoptotic Bax protein expression, whereas the preserved solution did not cause a significant difference. Bax/Bcl-2 ratio was found to be higher with the preservative-free solution.
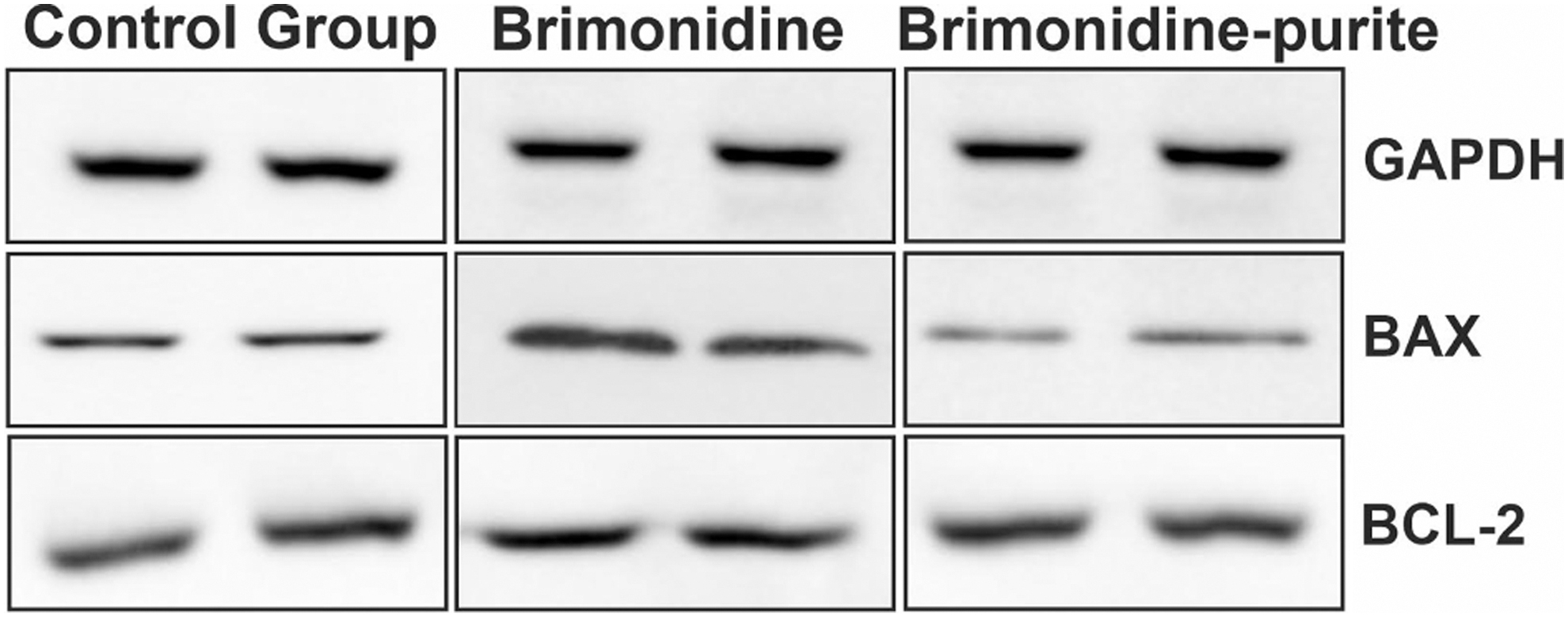
Effects of preserved and preservative-free 0.15% brimonidine on protein expressions apoptosis-related genes. Images of immunoreactive bands of Bax and Bcl-2 proteins after treatment with preserved and preservative-free brimonidine.
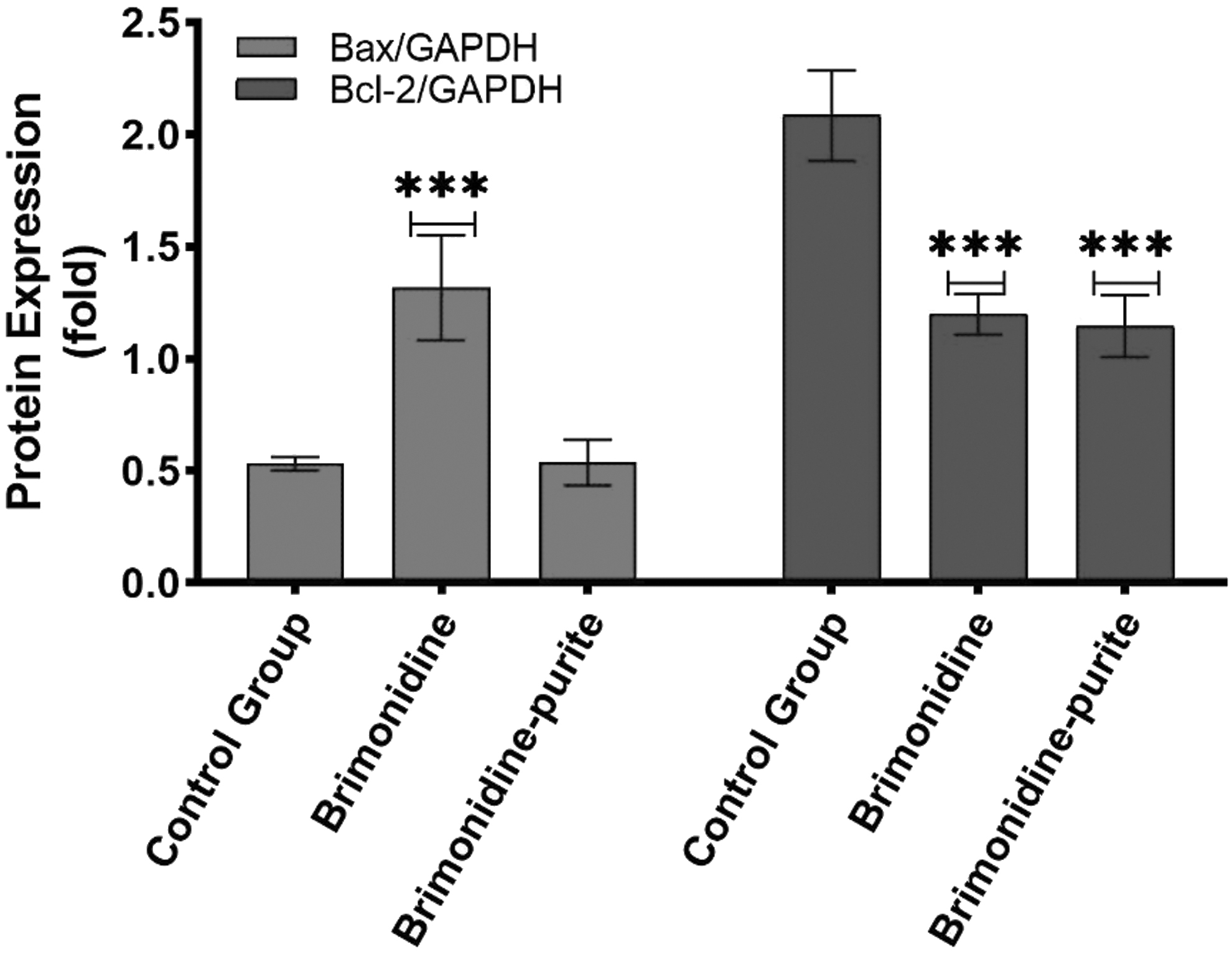
Relative protein expressions of Bax, Bcl-2 after treatment with preserved and preservative-free brimonidine. All experiments were repeated 3 times. The results were presented as mean ± SD, n = 4 (***P < 0.0001).
Discussion
Various studies have shown that long-term use of ocular hypotensive drugs can cause significant changes to the ocular surface.13,14 Because corneal epithelial cells comprise the first-line barrier of the eye, they constitute the cell group most exposed to the toxic effects of topical treatments. Problems of the ocular surface such as dry eye and allergies can be seen in a significant portion of glaucoma cases. 15 Preservative agents in antiglaucomatous preparations may exacerbate the condition and increase epithelial damage and ocular surface disease. The HCEC culture system we used in this study allows rapid evaluation of the toxic effects of ophthalmic solutions.
As previously stated by Friedlaender et al, eye drops are removed from the ocular surface within a few minutes. 16 Therefore, short-term applications are useful in terms of representing a single installation. According to the results of our cytotoxicity studies, it was observed that the preservative-free preparation had a greater cytotoxic effect at 5 min; there was no significant difference at 15 min and the following time periods (1, 6, 24 h). Alamar Blue has been shown to be highly sensitive to preservative toxicity in the evaluation of metabolic activity, even after exposure as short as 5 min. 7 The findings of this study do not coincide with the cellular toxicity studies conducted with preserved and preservative-free preparations in the literature.17–20
In a similar study, Xu et al 7 reported that polyquad, purite, sodium perborate, and sorbic acid do not cause significant cytotoxicity at short exposures of 5 and 15 min; preservative-free solutions were shown to have the lowest toxicity among the groups. Noecker et al 5 conducted an in vivo study by applying artificial tears carboxymethyl cellulose 0.5% containing purite BID, and various antiglaucomatous agents including brimonidine–purite 0.15% BID on New Zealand rabbits. According to the results of their study, there was no significant difference between brimonidine–purite 0.15% and artificial tears in terms of corneal damage. It was remarkable that the study demonstrated that the toxic effect of brimonidine on its own was indistinguishable from artificial tears containing purite as a preservative.
Evaluation of apoptosis with flow cytometric methods revealed a significant increase in early apoptotic activity (PE Annexin V+/7-AAD−), especially with preservative-free brimonidine. Similar to the cytotoxicity results, early apoptotic activity was observed more excessive in unpreserved brimonidine 0.15% compared with brimonidine–purite 0.15%.
Apoptosis has unique morphological and biochemical features, and caspases play a central role in the final phase. Alterations or anomalies in the apoptosis process contribute to the development of many diseases. 21 Apoptosis is stimulated in 2 different pathways, intrinsic (mitochondrial) and extrinsic.22,23 Apart from that, although Caspase-12 does not play a role in the mitochondria-related apoptotic pathway, it is thought to be involved in apoptotic mechanisms triggered by endoplasmic reticulum (ER) stress. 24 In terms of ophthalmological diseases, ER stress and misfolded protein response have been shown to play a major role in retinitis pigmentosa, macular degeneration, and glaucoma, which is the area of interest of our study. 25
To the best of our knowledge, this is the first study to investigate the effects of ophthalmic solutions containing brimonidine on genes and proteins involved in apoptosis. The preservative-free solution–treated cells showed higher expression of pro-apoptotic Bax, Caspase-9 and -12 genes, and Bax protein, whereas antiapoptotic Bcl-2 gene expression was insignificantly decreased and Bcl-2 protein expression was significantly decreased. On the contrary, brimonidine–purite insignificantly lowered the levels of pro-apoptotic Bax gene expression, significantly stimulated the antiapoptotic Bcl-2 gene expression, inhibited Caspase-3 and -9 gene expressions, and stimulated Caspase-12 gene expression. Bax protein expression was shown not to be affected and Bcl-2 protein expression was reduced. It was determined that the preservative-free solution triggers mitochondria-dependent apoptosis more prominently by activating the proapoptotic genes and caspase cascade stronger than the preserved solution (Fig. 8).
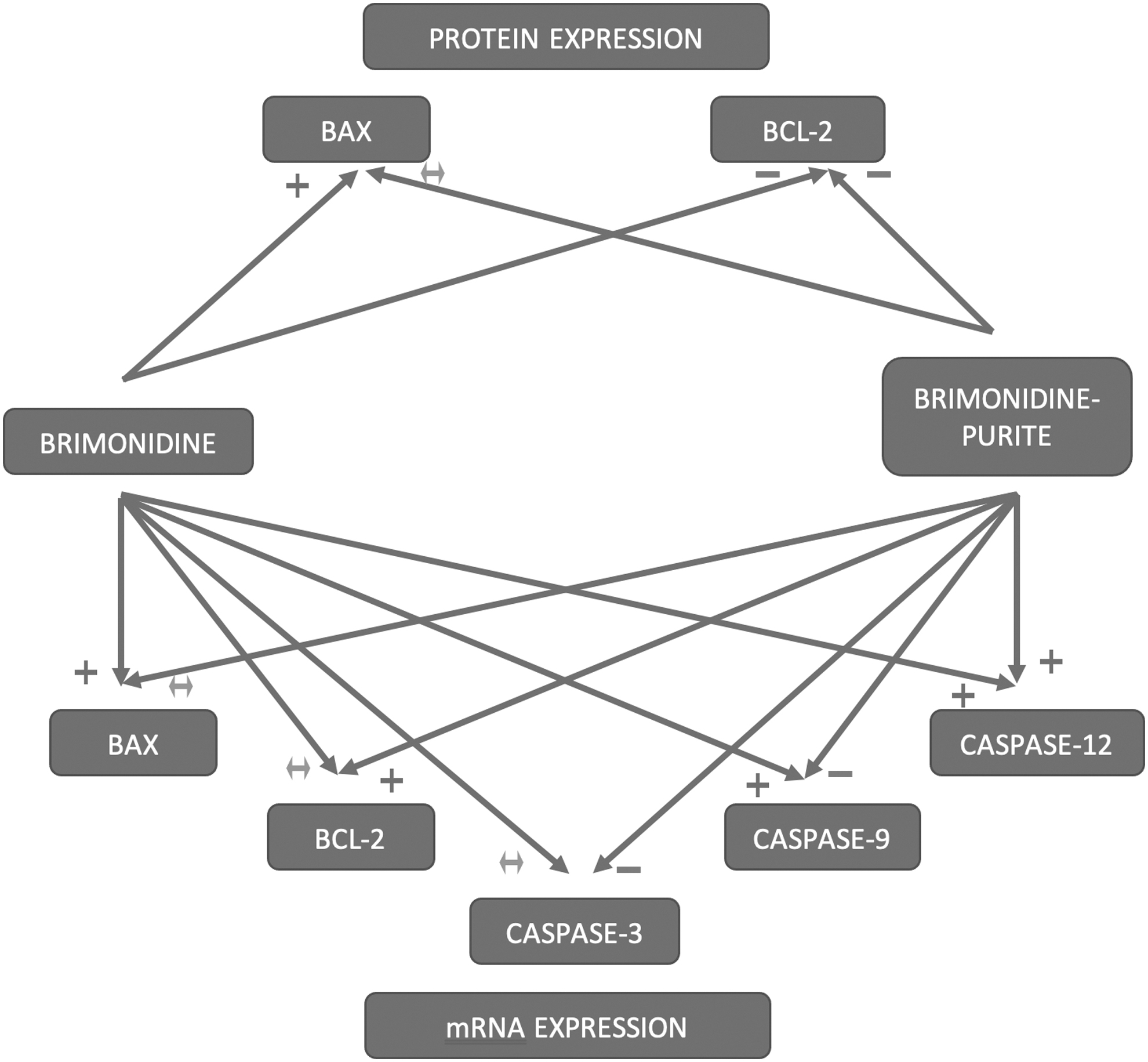
Summary diagram of the effects of brimonidine and brimonidine–purite on the mRNA and protein expressions involved in apoptosis. (+: stimulation of mRNA or protein product, P < 0.05; −: inhibition of mRNA or protein product, P < 0.05; ↔: change in mRNA or protein product is not statistically significant P > 0.05).
In addition, it seems that caspase-12 activation caused by the solutions may also trigger apoptosis through mitochondria-independent pathway. Brimonidine-purite–treated cells show significantly higher expression of Caspase-12 gene compared with preservative-free brimonidine. This may contribute to the increased apoptotic effect of brimonidine–purite compared with the control group. However, it could be said that both solutions trigger apoptosis through the mitochondria-dependent signaling pathway by significantly reducing the expression of anti-apoptotic Bcl-2 proteins and mitochondria-independent signaling pathway by increasing Caspase-12 gene expression. When flow cytometry, gene, and protein expression results are evaluated together, proapoptotic activity seems to be more apparent with the preservative-free solution.
The Bcl-2 is the key antiapoptotic protein that prevents the early apoptosis of the cells. Treatment with brimonidine–purite increased the expression of the Bcl-2 mRNA expression 2-fold compared with brimonidine group. Besides, this treatment also inhibited the expression of the central apoptotic protein, Bax. Brimonidine–purite was found to significantly decrease the Bax/Bcl-2 mRNA ratio. Thereby, brimonidine–purite may possess a protective effect against early apoptosis. Herein, the difference between the increase in gene expression and the level of protein encoded by the gene may pose a question mark. There are at least 3 known reasons for this weak correlation. First, many complicated and variable post-translational mechanisms are involved in the conversion of mRNA to protein, which have not yet been fully clarified. Second, the in vivo half-lives of proteins can vary considerably. Third, errors and noise occur in both protein and mRNA experiments, which makes it difficult to obtain a clear viewpoint.26,27
However, when viability findings and the results obtained from flow cytometric analysis of apoptosis are evaluated together, we gain a clearer perspective. Despite few supportive studies in the literature,15,28,29 our results regarding apoptosis also seem inconsistent with the strong evidence that preservatives or glaucoma drugs containing preservatives have more cytotoxic, antiproliferative, and proapoptotic effects in vivo and in vitro.30–34 However, supporting our early-term toxicity and apoptosis results, Duru and Ozsaygili compared preserved and preservative-free brimonidine 0.15% solutions in terms of ocular symptoms and tear parameters in primary open-angle glaucoma or ocular hypertension cases, and they found that the burning sensation was higher in the group treated with preservative-free solutions. 8
SOC is a preservative consisting of a combination of chlorine dioxide, chlorite, and chlorate that causes oxidation of intracellular lipid and glutathione and disrupts vital enzymes for cell function. It is an effective oxidizer as it has a high tendency to form free radicals. However, it rapidly decomposes on the ocular surface and converts into sodium and chloride ions, oxygen, and water. In our study, ROS levels yielded by brimonidine–purite 0.15% were higher than the negative control group and lower than the positive control group. Although the results were not statistically significant, they appear to be consistent with the oxidative potential of purite. Considering that most glaucoma cases are of advanced age and already carry varying degrees of oxidative stress in their ocular tissues, we would like to emphasize the importance of clinician awareness of treatments that will increase oxidative stress in their treatment approach.
During our investigation of the causes of the inconsistency between our cytotoxic and apoptotic results and the literature, in the short product information of the preservative-free solution, we found that silver ions were used in the cap design to provide antibacterial activity. Silver has been used in the past in the form of silver nitrate to prevent neonatal ocular infections. Today, data on silver uptake and metabolism are not well documented, but it is known to cause ocular irritation. 35 Calvery et al 36 studied the effects of silver salts on rabbit eyes and found that all the salts were irritative. We believe that the contamination of the preservative-free eye drops used in our study with silver ions implemented in the cap design may be related to the results we obtained. Further studies are required to gain clarification.
Use of the HCEC line was among the limitations of our study. Although this culture system allows rapid evaluation in terms of toxicity, three-dimensional culture systems may provide results closer to real ocular tissues. 37 However, in vivo models have the potential to provide a more accurate view of the corneal surface in terms of addressing both the interactions between epithelial and immunological cells and the relationship between epithelial cells and tear film. Despite this, in vitro models still maintain an important role in comparative drug studies, as these models also have disadvantages such as cost and difficulty in simulating chronic use.
Furthermore, the presence of a clinical study parallel to our study in the literature 8 provided a perspective in terms of in vivo studies. Second, eye drops were used in their commercial presentation in this study. Topical formulations contain preservatives and buffers as well as active ingredients. We were unable to identify the individual effects of each of these components in this study. Toxicity is usually caused by preservatives, but other components may also increase or decrease these effects.
In conclusion, the results of our study indicate that the brimonidine 0.15% solution containing purite was less cytotoxic to HCEC in the early period, had less early apoptotic activity, and did not cause a significant increase in ROS levels compared with the preservative-free solution. Further investigations of individual components are necessary to fully understand the toxicity of these ophthalmic solutions.
Footnotes
Authors' Contributions
A.K.: Conceptualization (lead); writing—original draft (lead); formal analysis (lead); writing—review and editing (equal). İ.M.D.: Software (lead); writing—review and editing (equal); methodology (lead); data curation (lead); investigation (lead). M.A.: Conceptualization (equal); review and editing (equal). S.K.: Conceptualization (supporting); Writing—review and editing (supporting); supervision.
Author Disclosure Statement
The authors have no conflicts of interests to declare that are relevant to the content of this article.
Funding Information
The authors declare that no funds, grants, or other support were received during the preparation of this article.