
Abstract
Purpose:
To examine the survival of neural progenitors (NPs) cells derived from human embryonic stem cells (hESCs) following subretinal (SR) transplantation in rodents.
Methods:
hESCs engineered to express enhanced green fluorescent protein (eGFP) were differentiated in vitro toward an NP fate using a 4-week protocol. State of differentiation was characterized by quantitative-PCR. NPs in suspension (75,000/μl) were transplanted to the SR-space of Royal College of Surgeons (RCS) rats (n = 66), nude-RCS rats (n = 18), and NOD scid gamma (NSG) mice (n = 53). Success of engraftment was determined at 4 weeks post-transplant by in vivo visualization of GFP-expression using a properly filtered rodent fundus camera. Transplanted eyes were examined in vivo at set time points using the fundus camera, and in select cases, by optical coherence tomography imaging, and after enucleation, by retinal histology and immunohistochemistry.
Results:
In RCS rats, cell rejection was observed in 29% of eyes at 6 weeks, rising to 92% at 8 weeks. In the more immunodeficient nude-RCS rats, the rejection rate was still high reaching 62% of eyes at 6 weeks post-transplant. Following transplantation in highly immunodeficient NSG mice, survival of the hESC-derived NPs was much improved, with 100% survival at 9 weeks and 72% at 20 weeks. A small number of eyes that were followed past 20 weeks showed survival also at 22 weeks.
Conclusions:
Immune status of recipient animals influences transplant survival. Highly immunodeficient NSG mice provide a better model for studying long-term survival, differentiation, and possible integration of hESC-derived NPs. Clinical Trial Registration numbers: NCT02286089, NCT05626114.
Introduction
Photoreceptor cell death is the major hallmark of a wide group of human blinding retinal degenerations such as retinitis pigmentosa (RP) and age-related macular degeneration (AMD) that are characterized by loss of photoreceptor cells in the outer nuclear layer (ONL).1–3 Mammalian photoreceptor cells are unable to regenerate, but recent studies have shown that exogenous sources of cells could provide a promising therapeutic strategy to replace lost photoreceptors.4–10 Such cell-based therapy to support/replace the degenerating cell type(s) may delay, halt, or perhaps even reverse degeneration, improve retinal function, and prevent blindness in these conditions.
Potential replacement sources of photoreceptors include retinal progenitors,11–13 primary photoreceptor progenitors,4,5,9,14–16 and photoreceptor progenitors derived from embryonic stem cells (ESCs)6,17–21 or induced pluripotent stem cells (iPSCs).7,22–28 In recent years, significant progress has been made in the differentiation of pluripotent cells toward retinal neural lineages, including photoreceptor cells,5–7,14,18,19,25,26,29 retinal ganglion cells,22–24,30–34 bipolar cells, and amacrine cells.8,35,36
Studies suggested that PSC-derived photoreceptors might engraft in the ONL after transplantation in retina-degenerated mice,18,25,37 form outer segment structures,6,7,12,18 and express rod photoreceptor markers.18,25,37,38 However, transplantation of ESC- or iPSC-derived photoreceptors is not yet being tested within human trials.
Our group is studying the potential of human embryonic stem cells (hESCs) to serve as a platform for the treatment of retinal and macular degenerations. One of our first initiatives was the development of a protocol that allowed directed differentiation of hESCs into retinal pigmented epithelium (RPE) cells in vitro. RPE cells play a crucial role in supporting and maintaining the photoreceptors and are the first cell type to fail in AMD. They are also involved in subtypes of RP and in a hereditary macular degeneration called Best disease, and in many ways an easier first target for cell-based retinal therapy than photoreceptors. Indeed, the hESC-derived RPE cells grown in vitro using our protocol, when transplanted into the subretinal (SR) space of Royal College of Surgeons (RCS) rats that serve as a model of retinal degeneration caused by RPE dysfunction, showed the ability to attenuate the disease process. 39
Following extensive in vitro and in vivo preclinical testing, a Phase I/IIa clinical trial in patients with advanced dry AMD was approved by the FDA and the Israeli Ministry of Health, and this trial was recently completed, leading to an optimization Phase IIa trial now being initiated.
Following this, we turned to explore the possibility of deriving retinal and photoreceptor progenitors from hESCs. A number of in vitro protocols were developed, and in the present study we addressed the ability of cells grown according to one of these protocols to survive following SR transplantation in different animal models. Thus, the purpose of the current study was to examine the survival and integration of premature neural progenitors (NPs) cells derived from hESCs following SR transplantation in rodents with a different immune status, in preparation for long-term experiments that will allow examining their ability to differentiate, integrate, and affect the course of retinal degeneration in rodent models.
Methods
Cell culture
Undifferentiated hESCs (cell line HADC102) were cultured on human cord feeders and were passaged every 7 days using collagenase A (Worthington Biochemical, Lakewood, NJ). 40 hESCs were genetically modified by a lentiviral vector to constitutively express enhanced green fluorescent protein (eGFP) to facilitate identification of the cells following SR transplantation in vivo. 41 This expression is strongly maintained also later during differentiation.
To obtain floating clusters of NPs in vitro, hESC colonies were picked up using collagenase A and cultured in a HydroCell plate (DiagnoCine, Hackensack, NJ) for 4 weeks as spherical bodies (SBs) in NutriStem hPSC XF medium (Biological Industries, Beit Haemek, Israel) supplemented with 50 U/mL penicillin and 50 μg/mL streptomycin (Gibco-BRL, Gaithersburg, MD). The media were changed twice a week and contained nicotinamide (NIC, 10 mM; Sigma, St. Louis, MO) and basic fibroblast growth factor (bFGF, 20 ng/mL; PeproTech, Inc., Rocky Hill, NJ) during the entire time. CHIR99021 was added to the culture (CHIR, 3 μM; PeproTech, Inc.) in the last 2 weeks of differentiation.
Before transplantation, the SBs were dissociated to single cells using TrypLE Select (Gibco-Thermo Fisher Scientific). After washing the cells in phosphate-buffered saline (PBS, Biological Industries), they were resuspended and filtered in 20 mL of balanced salt solution plus (BSS plus; Biological Industries). Using trypan blue dye (0.4%; Gibco-Thermo Fisher Scientific), the cells were stained, counted, and their viability was determined.
Quantitative-PCR analysis
Total RNA was extracted from undifferentiated HADC102 and from SBs differentiated for 4 weeks using the Quick RNA™ MicroPrep kit (ZYMO Research, Irvine, CA). cDNA was synthesized with Moloney murine leukemia virus reverse transcriptase (M-MLV RT) and specific primers, according to the manufacturer's instructions (Promega Corporation, Madison, WI). For quantitative-PCR (qPCR), TaqMan Assays-on-Demand Gene Expression Products, TaqMan Universal PCR Master Mix, and ABI Prism 7900HT Sequence Detection System (Applied Biosystems, Foster City, CA) were used. B-glucuronidase (GusB) was an internal reference for normalization.
The following TaqMan Assays-on-Demand were used: Oct4, assay ID Hs01895061; Pax6, assay ID Hs00240871; Six3, assay ID Hs00193667; Six6, assay ID Hs00201310: Rx1, assay ID Hs00429459; Chx10, assay ID Hs01584048; Crx, assay ID Hs00230899; NRL, assay ID Hs00172997; rhodopsin, assay ID Hs00892431; GusB, assay ID Hs99999908.
Immunocytochemical staining
After 4 weeks in suspension, SBs were dissociated using TrypLE Select, cultured on poly-D-lysine and laminin, and immunophenotyped with primary antibodies including anti-b-tubulin III (mouse IgG 1:1,000; Sigma), anti-nestin (rabbit polyclonal,1:300; Chemicon, Temecula, CA), and anti-Rx (rabbit polyclonal, 1:100; Abcam, Boston, MA). Primary antibodies were detected by using goat anti-rabbit Ig conjugated to Texas red (1:500) and goat anti-mouse IgG conjugated to Cy3 (1:500; both from Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, www.jacksonimmuno.com), Nuclei were counterstained with 4, 6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA).
Animals
All animal experiments were conducted according to the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research and approved by the institutional committee for animal research.
The following animal models were used in the present study:
RCS rats (n = 66) have a mutation in the Mertk gene that causes inability of the RPE to phagocytose shed photoreceptor outer segments. Accumulation of interphotoreceptor matrix proteins at the basal outer segment zone occurs, leading to apoptosis of photoreceptor cells and retinal degeneration. Although the RCS rat model does not fully recapitulate AMD pathophysiology, this model is widely used for testing novel therapeutic approaches for AMD as in AMD, dysfunction of the RPE is also a major cause of disease. Nude-RCS rats (n = 18): The congenitally athymic nude rat (rnu/rnu) is a genetic mutant found in Rattus norvegicus characterized by thymus aplasia and a severe alteration in the cell-mediated branch of immunity. Altered cellular immune responses are due to lack of appropriate development and maturation of functional T cell (thymus-derived) lymphocytes. In contrast to the defective T cell-mediated immune responses, natural killer (NK) cells and phagocytic function are not impaired in athymic nude rats. We performed crossbreeding between RCS rats and nude rats hoping to derive a model that on the one hand will manifest a clinically relevant retinal degeneration, and on the other will allow long-term survival that is crucial for differentiation and maturation of hESC-derived NPs. The NOD scid gamma mouse (NSG mouse) (n = 53) carries NOD.Cg-Prkdcscid and Il2rgtm1Wjl mutations and is among the most immunodeficient animals described to date. NSG mice lack mature T cells, B cells, and NK cells and they are also deficient in multiple cytokine signaling pathways. Deep immunodeficiency may potentially provide long-term engraftment of human cells.
SR transplantation of hESC-derived NPs
The study included animals of both genders. At 3 weeks of age, the animals were anesthetized with an intraperitoneal mixture of ketamine (85 mg/kg; Vetoquinol, Lure, France) and the relaxing agent xylazine (15 mg/kg; Eurovet Animal Health, Bladel, Netherlands). Local anesthetic drops (benoxinate HCl 0.4%; Fischer Pharmaceuticals, Israel) were administered. The pupils were dilated with 1% tropicamide and 2.5% phenylephrine HCl (all ocular drops from Fisher Pharmaceuticals, Tel-Aviv, Israel).
To deliver the cells subretinally, we used a transvitreal approach in mice and a transscleral approach in rats. This is necessary because of the different sizes of the eyes in these 2 species. All procedures were performed under direct visualization of a dissecting microscope.
In mice, a small incision was made 1 mm posterior to the limbus using a 30G needle. A 33G blunt needle connected to a Hamilton syringe (Hamilton, Reno, NV) was advanced through the vitreous and the retina, and 1 μL (75,000 cells) of hESC-derived NPs in BSS was injected into the SR space. The fellow eye served as an untreated control.
In rats, a transscleral route of injection was used. A self-sealing scleral incision was made using the tip of a 30G needle with the bevel pointed up. A scleral tunnel through the sclera was made avoiding penetration of the retina. The cornea was contralaterally punched decreasing intraocular pressure to prevent reflux of the cells. A 33G blunt needle connected to a Hamilton syringe was then advanced via the scleral tunnel into the SR space and 1.5 μL of 75,000 cell/μL suspension was injected.
Following injections, chloramphenicol 5% antibiotic ointment (Rekah Pharmaceutical, Holon, Israel) was topically applied, and animals were placed on the heating pad for recovery.
Successful SR delivery with SR bleb formation was visually confirmed immediately after injection, and animals with failed delivery were excluded from the study. Furthermore, at 4 weeks post-transplant, all animals underwent retinal imaging as outlined below to examine eGFP expression, and only eyes with a clear SR GFP-positive graft were maintained for further follow-up and included in the survival analysis. Successfully transplanted eyes with viable grafts at 4 weeks post-transplant were followed in vivo by serial GFP-filtered funduscopy and optical coherence tomography (OCT) imaging in addition to postenucleation histological and immunohistochemical analysis. Follow-up time was dependent upon evidence of graft survival in the different animal models studied, with some NSG mice followed until 22 weeks post-transplant.
After week 4, disappearance of the eGFP signal, development of new retinal detachment, and/or mass formation as evident on in vivo imaging were suspected as representing graft rejection and this was then confirmed by histological and immunohistochemical analysis.
Retinal imaging and OCT
Color fundus imaging, as well as fluorescent fundus photography to identify GFP expression, was performed under general anesthesia using a Micron III system designed to take images of the rodent retina (Phoenix Technology Group, Pleasanton, CA).
In vivo high-resolution OCT was performed using a Heidelberg Spectralis OCT system (Heidelberg Engineering, Inc., Carlsbad, CA) modified for use in rodents.
Histology and immunohistochemistry
At different time points, according to the in vivo imaging results, animals were euthanized, and eyes were enucleated, fixed in Davidson solution (glacial acetic acid, 95% alcohol, 16% paraformaldehyde, and distilled water in a ratio of 4:12:5:15, respectively) for 24 h, embedded in paraffin (Paraplast Plus; Leica Biosystems Nussloch GmbH, Nußloch, Germany), and serially sectioned at 5 μm.
Some sections were then stained with hematoxylin and eosin for descriptive histology. For immunohistochemistry, deparaffinized sections were incubated in a decloaking chamber (Biocare Medical, Pacheco, CA) with 10 mM citrate buffer (pH 6.0) at 125°C, blocked with PBS solution containing 1% bovine serum albumin, 0.1% Triton X-100, and 10% normal donkey serum, and subsequently incubated for 1 h with one of the following primary antibodies: mouse anti-human nuclei (Millipore, 1:80), goat anti-GFP conjugated to FITC (Abcam; 1:400), rabbit anti-recoverin (Millipore, 1:350), mouse anti-rhodopsin (Thermo Fisher, 1:40), rabbit anti-synapsin (Abcam; 1:175), mouse anti-PKC-α (Protein kinase C alpha; Santa Cruz, 1:100), mouse anti-CD3 (Bio-Rad Laboratories, 1:10), rabbit anti-Ki-67 (Thermo Fisher, 1:75), mouse anti-β-tubulin III (Sigma; 1:6,000), and rabbit anti-nestin (Sigma-Aldrich; 1:1,000).
After washing in PBS, specimens were incubated for 1 h at room temperature with proper secondary antibodies. The specificity of immunohistochemical staining was determined by negative control, replacing the primary antibody with serum.
All observations and photography were performed using a fluorescent microscope (BX41; Olympus Corp., Tokyo, Japan) equipped with a digital camera (DP70; Olympus Corp.). Image processing and quantification were performed using a raster graphics editor (Photoshop CS3; Adobe Systems, Mountain View, CA).
Results
Derivation of NPs from hESCs
To induce differentiation toward an anterior forebrain and eye fate, we cultured the hESCs in suspension as SBs for 4 weeks in the presence of NIC and bFGF, with CHIR added to the media during the last 2 weeks of differentiation. qPCR demonstrated that the NPs did not express Oct4, a marker of undifferentiated hESCs, and showed high expression of anterior forebrain and early eye field transcription factors such as Pax6, Rx, Six3, and Six6 42 compared with undifferentiated hESCs (Fig. 1A).
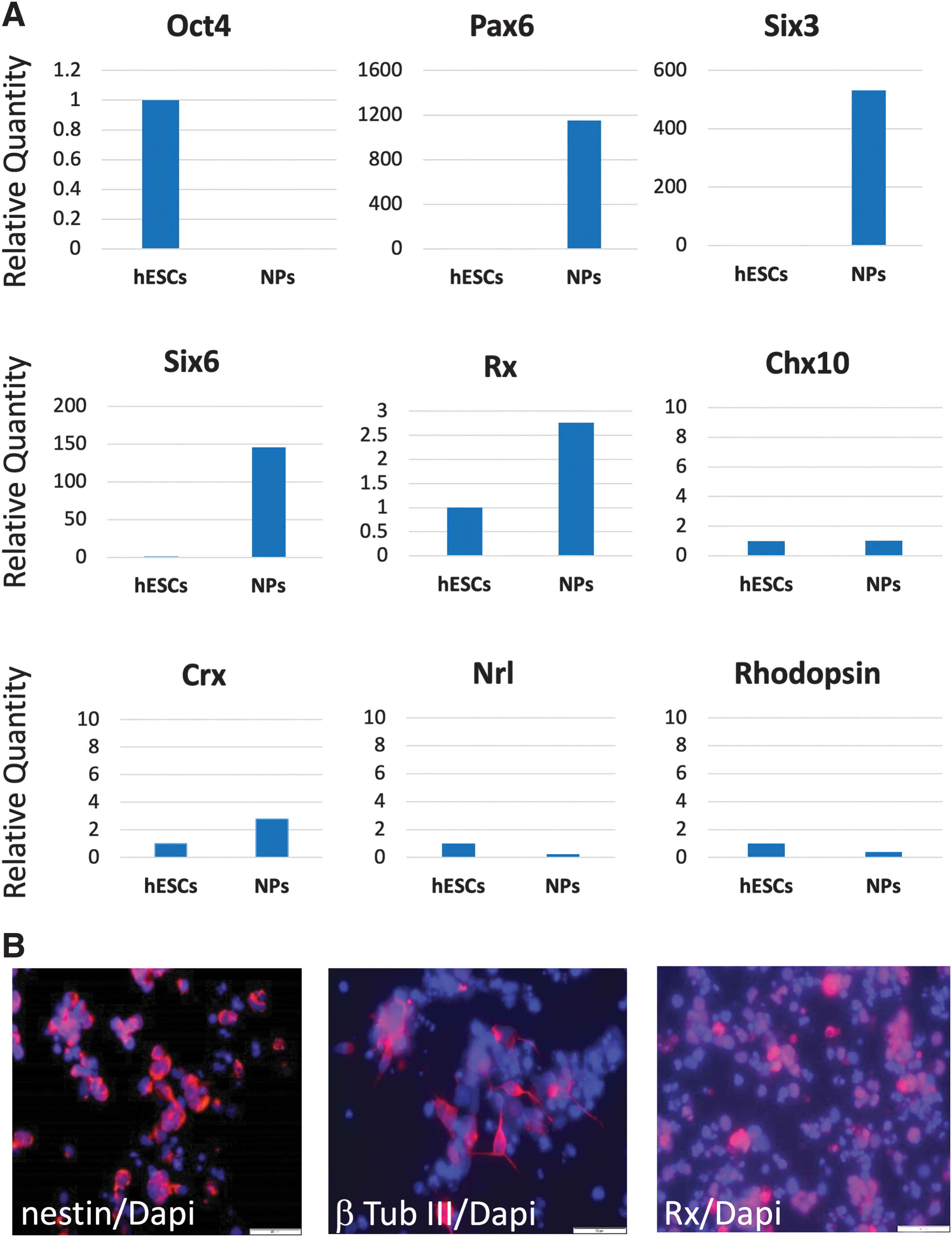
In vitro differentiation of hESCs toward an anterior forebrain and early eye field fate in the presence of NIC, bFGF, and CHIR.
However, there was no expression of the more mature neural retina transcription factor visual system homeobox 2 (VSX2/Chx10), 43 and transcripts of the retinal and photoreceptor markers such as Crx, Nrl, and rhodopsin were also not expressed in the NPs. Immunofluorescent staining after 4 weeks of differentiation in vitro using anti-nestin, anti-β-tub III, and anti-Rx antibodies revealed hESC differentiation toward a neural fate (Fig. 1B). 44 After SB dissociation for transplantation, the viability of cells was 90.2% ± 2.9%.
Survival and differentiation of hESC-derived NPs following SR transplantation in dystrophic RCS rat eyes
1.5 μL of hESC-derived NP cell suspension at a concentration 75,000 cells/μL was delivered into the SR space of 66 RCS rat pups at the age of 3 weeks. Retinal structure, cell survival, and engraftment were studied in vivo and histologically at different time points up to 9 weeks post-transplant. In vivo OCT performed 4 weeks following transplantation did not reveal a retinal detachment (defined as separation of the photoreceptors from the RPE/SR graft) in transplanted eyes, whereas SR grafts were distinctly visualized (Supplementary Fig. S1A).
These findings well correlated with histological results as shown in Supplementary Fig. S1B. To verify the human origin of engrafted cells and specificity of GFP expression, retinal sections were double stained with anti-GFP and anti-human nuclei primary antibodies. Supplementary Figure S1C shows high levels of expression of both markers mainly restricted to the SR grafts. Importantly, no GFP expression was found in the host ONL adjacent to the transplanted cells. Thus, GFP labeling provides a reliable and specific tool for recognizing the transplanted cells in retinal sections.
At 4 weeks post-transplant, the hESC-derived NPs in the SR grafts did not express the photoreceptor-specific marker recoverin, the synaptic marker synapsin, or the rod bipolar cell marker PKC-α (Fig. 2, upper row). In contrast, at 7 weeks post-transplant, immunohistochemical studies showed that the transplanted hESC-derived NPs further differentiated in vivo as recoverin and synapsin expression was clearly seen in the SR grafts. PKC-α was again not expressed in the grafts (Fig. 2, lower row).
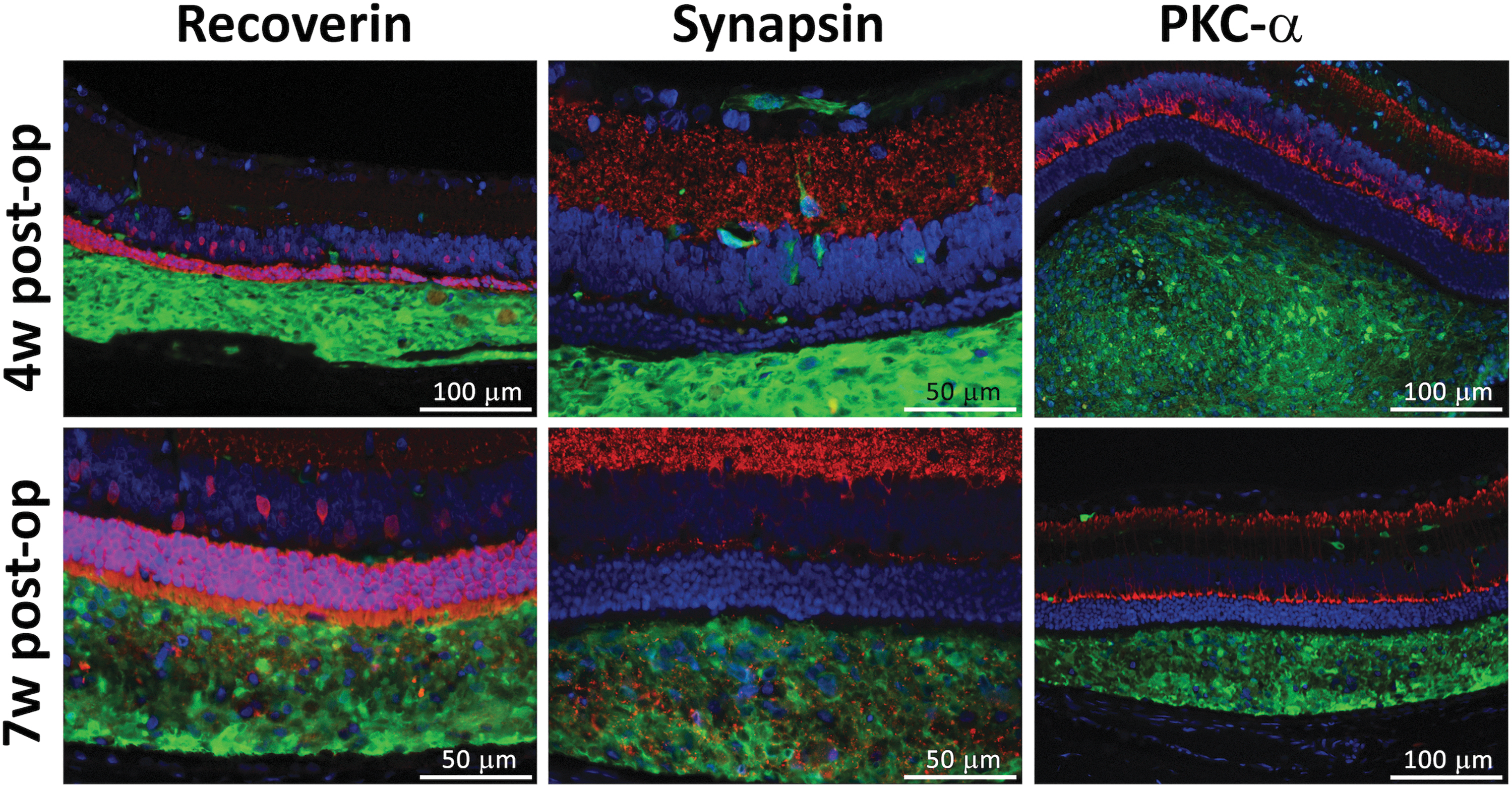
Following SR transplantation, hESC-derived NPs show further differentiation in vivo over time. At 4 weeks post-transplantation (upper row), the hESC-derived NP cells expressed GFP confirming their identity, but did not express the photoreceptor-specific protein recoverin, the synaptic marker synapsin, or the marker of rod bipolar cells PKC-α (red fluorescence). These markers are seen in the cells of the host retina at the relevant locations, but they do not colocalize with the anti-GFP staining marking the transplanted cells. At the 7-week time point (lower row), expression of recoverin and synapsin was observed within the SR grafts colocalizing with GFP expression, suggesting differentiation of the transplanted cells in the SR space in vivo over time. GFP, green fluorescence; NP, neural progenitor; PKC-α, protein kinase C alpha; SR, subretinal.
Survival of transplanted cells in vivo was assessed by using a Micron III system equipped with a proper set of filters for GFP visualization. Serial follow-up examinations of the transplanted RCS eyes in vivo revealed attenuation of GFP expression over time, which signifies the loss of the SR hESC-derived NPs (Fig. 3A). Rejection of the transplanted NPs often manifested as development of retinal detachment clearly seen in color and fluorescent fundus images with progression over time, including detachment of the retina from the RPE and/or from and around the SR graft (Fig. 3B). Detachment of the retina with accumulation of cells and debris was also confirmed by histology (Fig. 3C). Loss of the GFP signal and signs of rejection were observed in 29% of RCS rat eyes at 6 weeks post-transplant, rising to 92% rejection by 8 weeks, as reflected in the summarizing, concluding graph shown in Fig. 7 (blue line).
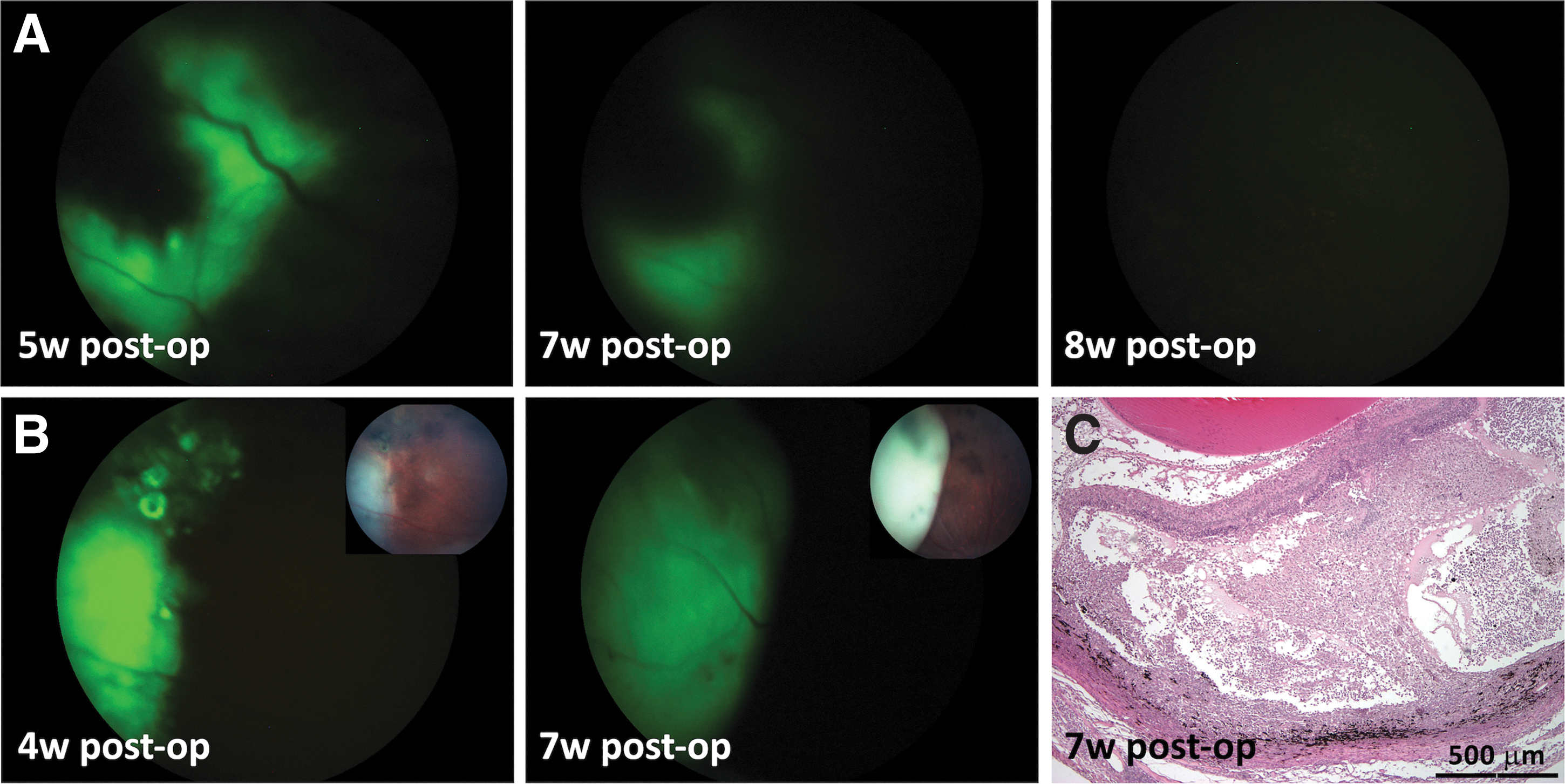
Rejection of hESC-derived NPs following SR transplantation in RCS rats.
In some cases, the presence of a large number of cells in the SR space was monitored using in vivo OCT imaging (Fig. 4A). However, these SR grafts did not reflect the accumulation of human-derived transplanted cells, since no GFP fluorescence was observed by Micron III GFP-filtered imaging (Fig. 4B) as well as by negative anti-GFP and anti-human nuclei immunohistochemistry (Fig. 4C–E). We presumed that this kind of SR collection of cells may reflect a rejection process and consists of host immunocompetent cells. Indeed, we found that many cells were positive for CD3, a marker of T cells, indicating a host T cell-mediated inflammatory responses in the SR graft area (Fig. 4F).
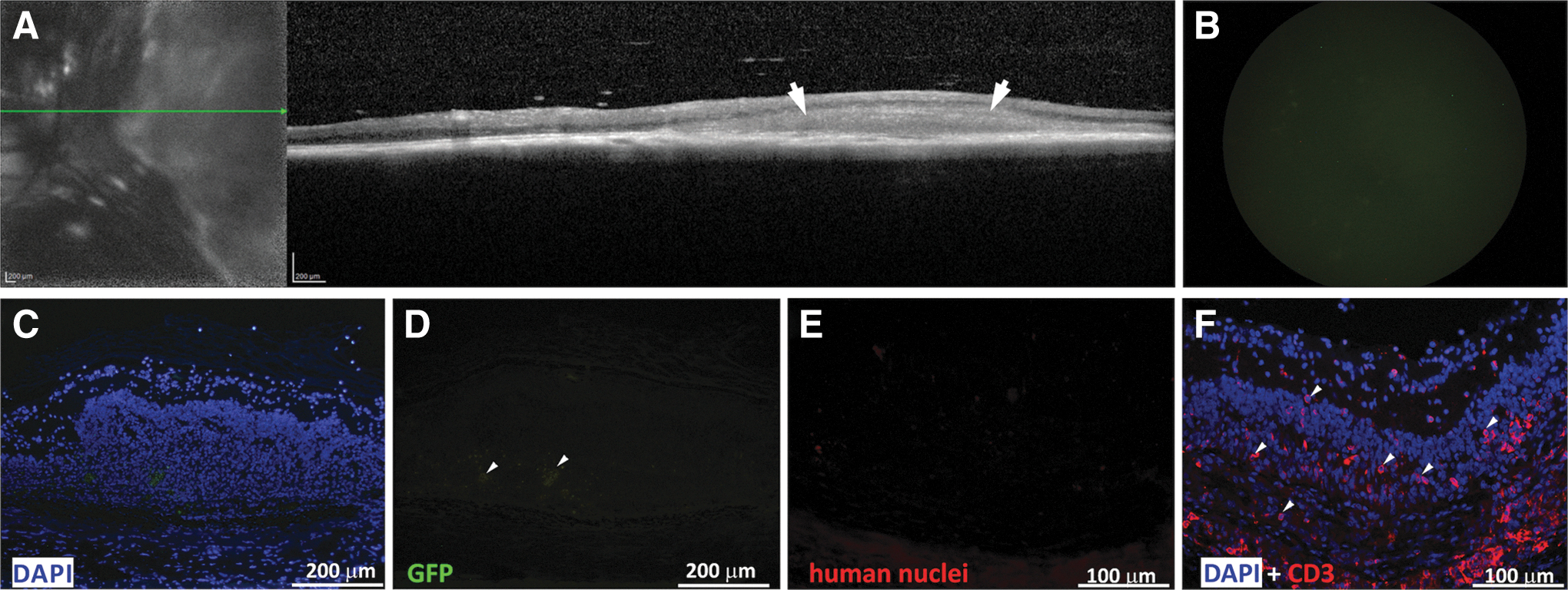
T cell-mediated rejection of an SR graft in RCS rat eye.
Thus, in contrast to RPE cells derived from the same HADC102 hESC line that survived for 16 weeks and more following SR transplantation in RCS rats, 39 hESC-derived NPs underwent cell-mediated rejection within <9 weeks. This occurred despite the impaired ability of RPE cells of the RCS model to perform phagocytosis and even when cyclosporin A (210 mg/L) was supplied in the drinking water as an immunosuppressant medication.
To allow longer term survival, one possible solution was to attempt to reach an animal model combining the same disease features, with deeper impairment of the immune system function. For this purpose, we crossbred RCS rats with congenitally athymic nude rats lacking mature T cells and used the new nude-RCS rat model for further transplantation experiments.
Survival of hESC-derived NPs cells after SR transplantation in nude-RCS rat eyes
Transplantation in nude-RCS rats was performed in the same way described for RCS rats. Despite lacking mature T cells, this model did not provide better survival of transplanted hESC-derived NPs. Moreover, rejection of 62% of grafts was already noted at 6 weeks post-transplant (summarizing Fig. 7, brown line), often associated with retinal detachment. In vivo OCT imaging 6 weeks post-transplant showed retinal detachment with an SR hyporeflective space, most probably fluid (Fig. 5A). In another eye, serial in vivo GFP-filtered and color fundus imaging taken at 4 and 7 weeks post-transplant showed rapid and significant expansion of the GFP signal accompanied by elevation of the retina (Fig. 5B, C).
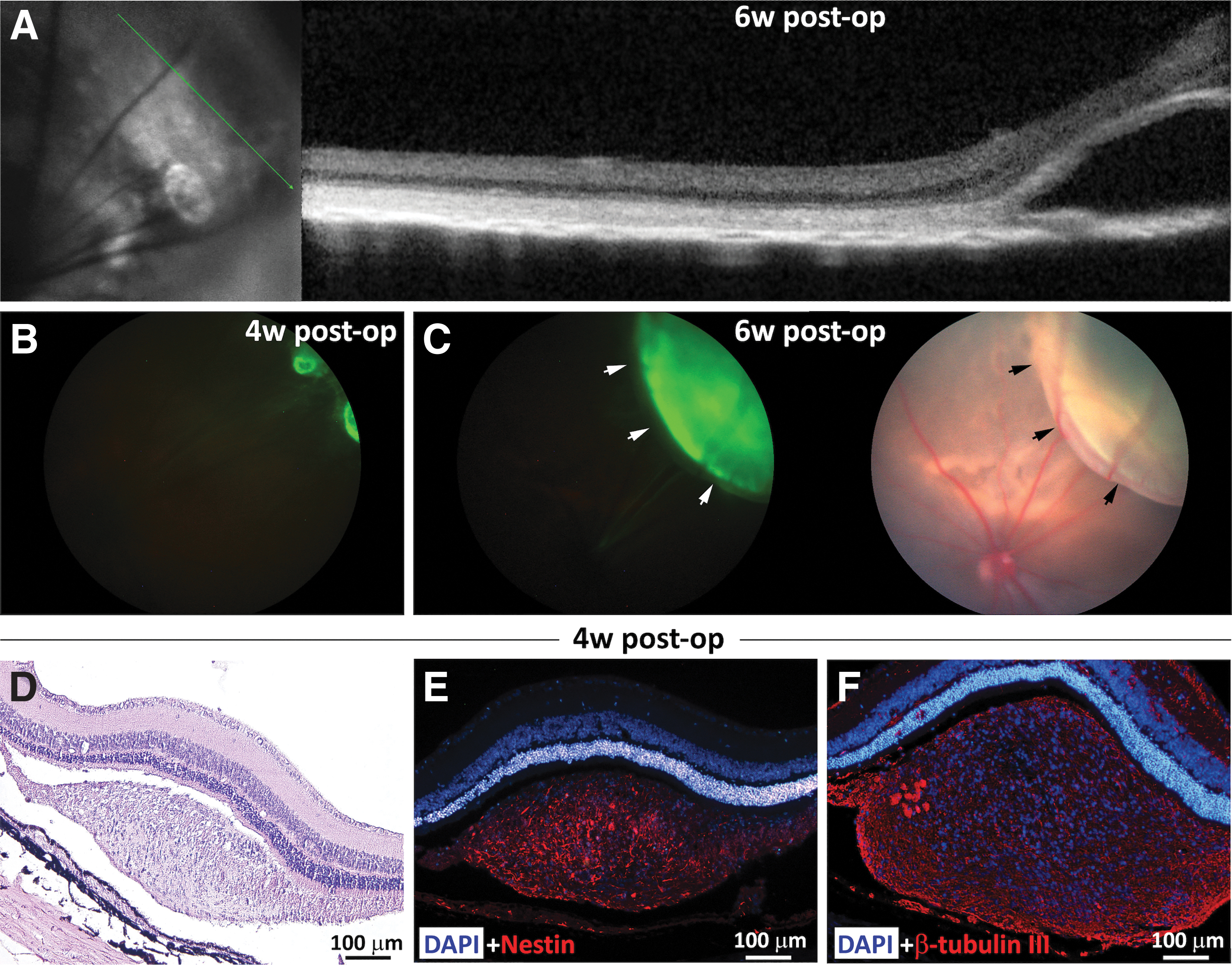
Development of retinal detachment as a sign of rejection of hESC-derived NPs in nude-RCS rats.
On histological evaluation, retinal detachment was sometimes associated with proliferation of cells assuming a “tumor-like” formation as evident in a hematoxylin and eosin-stained histologic section (Fig. 5D). Transplanted cells expressed high levels of the neural stem/progenitor cell marker nestin (Fig. 5E) and the neuronal marker beta-tubulin III (Fig. 5F). Costaining with anti-GFP antibody showed strong GFP expression (Supplementary Fig. S2A, B), confirming hESC origin of the cells.
Thus, the nude-RCS rat also did not provide a suitable model for long-term survival of hESC-NPs following SR transplantation.
Survival and incorporation of hESC-derived NP cells after transplantation in NSG mice
Following failure of the nude-RCS model to provide long-term survival of hESC-derived NPs, we decided to use for transplantation the most immunodeficient animal model available to date, namely NOD.Cg-Prkdcscidll2rtm1Wji (NSG) mice. These animals lack mature T cells, B cells, and NK cells and also have multiple impaired cytokine signaling pathways.
Indeed, SR transplantation of hESC-derived NPs in NSG mice showed 72% graft survival at 20 weeks (summarizing Fig. 7, green line). Serial in vivo GFP-filtered fundoscopy demonstrated engraftment and survival of hESC-derived NPs up to 22 weeks post-transplant (Fig. 6A). Positive anti-GFP immunostaining of retinal sections then confirmed the hESC origin of the engrafted cells (Fig. 6B). In a small number of eyes, in vivo OCT imaging demonstrated formation of “tumor-like” structures (Fig. 6C). Immunohistochemistry demonstrated strong anti-GFP expression (Fig. 6D) and a high level of cell proliferation within the “tumor-like” structure as determined by anti-Ki-67 immunostaining (Fig. 6E).
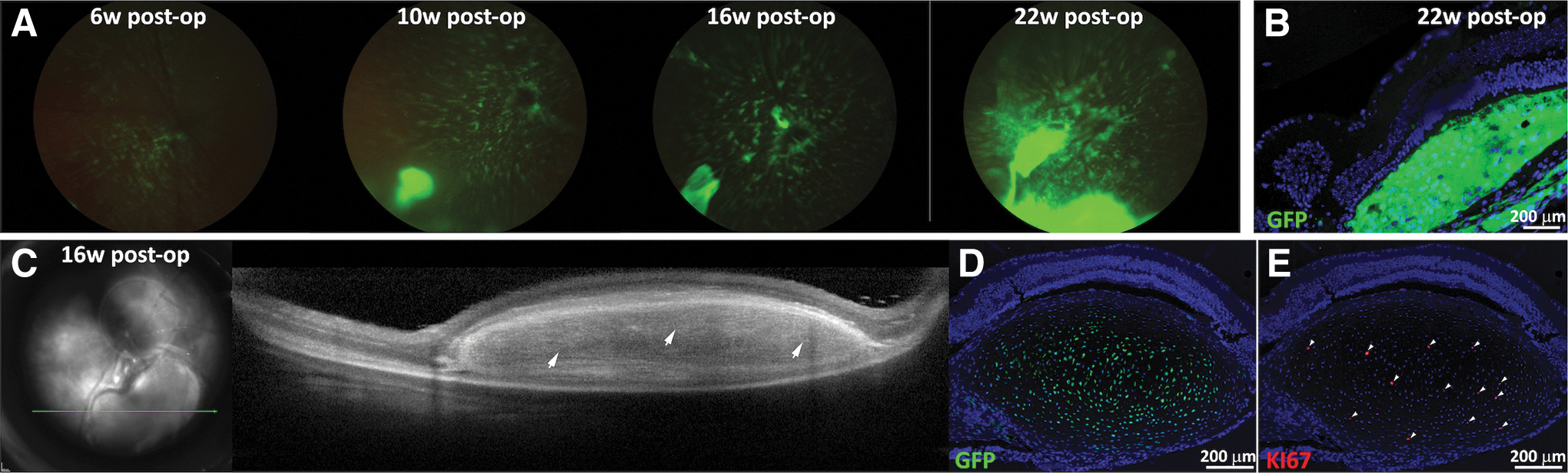
Long-term survival of hESC-derived NPs in NSG mice.
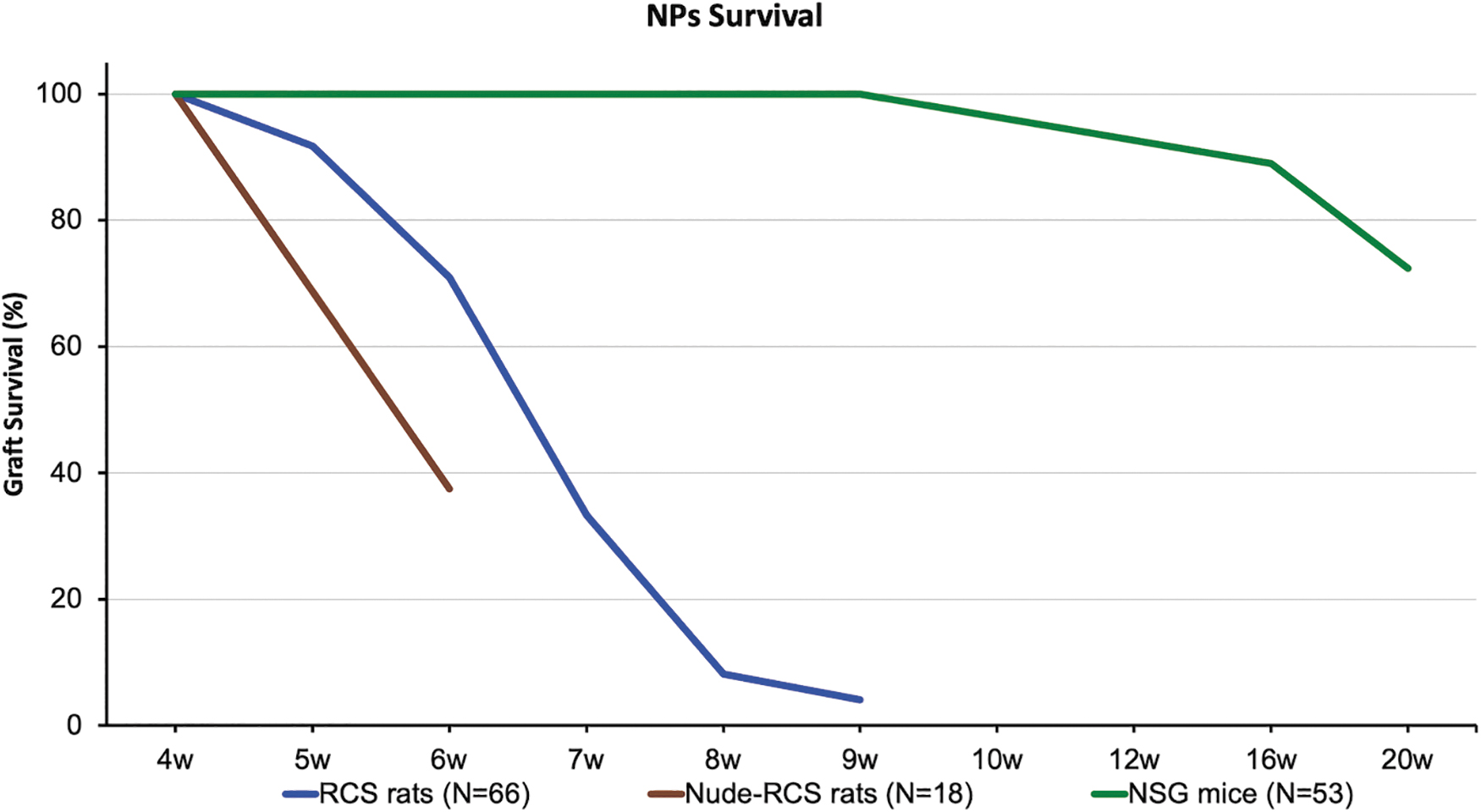
Summarizing figure: survival rate of hESC-derived NPs following transplantation into the SR space of RCS, nude-RCS, and NSG mice.
The survival rate of hESC-derived NPs following transplantation into the SR space of RCS, nude-RCS, and NSG mice is demonstrated in the summarizing graph (Fig. 7).
Discussion
One strategy to restore vision in degenerative retinal diseases such as RP and AMD is the application of cell-based therapy, aimed at supporting/replacing the failing cells of the host. Typically, patients lose vision when the outer retinal photoreceptor layer is lost. At present, there are no proven effective means of restoring vision to patients with these diseases, but research in this field is accelerating and retinal cell replacement through transplantation is being actively explored.7,8,45–55 The therapeutic goal would be to preserve vision by supporting host photoreceptor cells that are still surviving, as well as perhaps restore and replace lost cells. This would require survival, maturation, and connection of the transplanted cells with the inner retinal cells of the host.
Several studies utilized human hESC-derived retinal cells in different animal models.6,10,33,45,56–58 It places transplanted human cells in a more challenging environment (as they are xenogeneic grafts) than would be expected in the actual clinical transplantation (where they will be allogeneic grafts). This raises several questions regarding the optimal animal models as well as optimal configuration of donor cells (cell suspension vs. sheets), barriers to integration and donor cell function, and the need and role of immunosuppression to accompany transplantation.10,45,56,58–60
Before moving toward human trials, the potential of hESC-derived NPs to integrate and function must be tested in animal models of disease. This requires models that would allow the transplanted cells to survive despite the xenograft conditions, which could lead to cell rejection manifesting as ocular inflammation, retinal detachment, and ultimately death of the transplanted cells. It is thus necessary to identify permissive models and conditions that would provide the required time for integration, differentiation, and establishment of connectivity of the transplanted cells without cell rejection being evoked.
Among cell transplantation sites, the eye and particularly the SR space are considered to be advantageous as they are viewed as immune-privileged sites.61,62 Several reports showed that allografts of fetal retina or retinal progenitor cells survive in the SR space without immunosuppression.63–65 Indeed, our group has shown that hESC-derived RPE cells can survive in the SR space of RCS rats for months and such cells can induce immune tolerance.39,66 In our previous study, hESC-derived RPE cells transplanted to the SR space of RCS rats survived up to 16 weeks, and may have immunosuppressive effects through inhibition of T cell responses.39,66 This may be mediated by inhibition of interferon-γ secretion and reduced proliferation of activated T cells.
Furthermore, RPE cells enhanced T cell apoptosis and secretion of the anti-inflammatory cytokine interleukin-10, while they altered the expression of the T cell activation markers CD69 and CD25. 66 Thus, RPE cells may be able to induce an “immune-tolerant” state. In contrast, even though derived from the same HADC102 hESC line, we found that the transplanted NPs were not able to survive in the long term following transplantation into the SR space of RCS rats: while at 6 weeks the survival rate was still 71%, by 8 weeks this decreased to 8% with the development of retinal detachment in most of the cases.
The occurrence of transient neurosensory retinal detachment following SR injection of cells in suspension is a sign of successful transplantation and the fluid component is usually absorbed within a few days. Then, survival of the cells delivered can be identified by in vivo GFP imaging and OCT imaging, which demonstrates a graft between the host RPE and overlying host retina with the different layers adherent and in close proximity to each other. Indeed, we confirmed successful transplantation by this method in all animals 4 weeks postinjection and only animals with positive engrafting continued follow-up to determine longer term survival. Later on, if retinal detachment reoccurred and progressed, this was often a sign of immune response and rejection.
In our previous studies with SR delivery of hESC-derived RPE cells, we did not see such detachments. Thus, HADC102 hESC-derived NPs probably stimulate a more significant immune response in the host retina than RPE cells derived from the same source.
To improve survival of the hESC-derived NPs, we performed crossbreeding between RCS rats and athymic nude rats (Foxn1rnu/rnu), which are often used to test xenografts of human tissue.65,67 This allowed creating an immunodeficient rat model that manifests the dystrophic features of RCS rats and lacking T cells. Surprisingly, despite this immune deficiency, the rate of NP survival in this animal model was even lower than in the RCS rats, being 38% at 6 weeks with a prominent host immune response to the transplanted graft. These animals do possess NK cells and phagocytic function, and these may be mediating the rapid rejection.
Zhu et al. demonstrated that photoreceptor integration improved up to 9 months post-transplantation in IL2RG-deficient mice which carry a mutation in the interleukin 2 receptor-γ chain gene on the X chromosome resulting in a 10-fold reduction in the absolute number of lymphocytes and a complete absence of NK cells, which mediate cell rejection. 68 Therefore, we proceeded and investigated the ability of hESC-derived NPs to survive in the SR space of NSG mice that lack mature T cells, B cells, as well as NK cells. In this study, we were indeed able to achieve 89% survival of the NPs at 16 weeks post-transplantation and 72% survival at 20 weeks.
Following these results, we initiated crossbreeding between NSG mice (that have a normal retina) and rd10 mice 54 (that manifest severe retinal degeneration caused a mutation in the PDE gene) to obtain an aggressive model of retinal degeneration. We are also trying to develop NSG-FAM161A mice, which combine NSG-level immunodeficiency with a milder course of retinal degeneration caused by FAM161A gene knockout. 69 We are hoping that these 2 mice models will allow us to better assess the ability of hESC-derived NPs to differentiate, integrate, and perhaps attenuate the rate of disease following long-term survival in the microenvironment of a degenerating retina.
In summary, NSG mice allow long-term survival of NPs derived from our HADC102 hESC line without the need of additional immunosuppression, and can serve for testing maturation, integration, and function of retinal and photoreceptor precursors in preparation for future clinical trials.
Footnotes
Authors' Contributions
H.A. conceived, planned, and executed the experiments, analyzed data, prepared the figures, and wrote the article. C.M. planned and executed the experiments, and analyzed data. A.O. planned and executed the experiments, analyzed data, prepared the figures, and wrote the article. A.E., H.K., M.I., and B.R. executed the experiments and analyzed data. E.B. Supervised the work, conceived and planned the experiments, analyzed data, and wrote the article. All authors read the final version of the article and approved it.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Moxie Foundation, the Yedidut 1, and the GT Foundation Research Grants.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.