
Abstract
Background:
Prostaglandin (PG) receptor agonists are the first-line eyedrop medication treatment for glaucoma. The pathophysiology of this disease is not completely known, and elevated intraocular pressure (IOP) is the key risk factor. The membranes of the axons (of the retinal ganglion cells) passing through the optic nerve (ON) head experience significant damage. Lipids are an essential component of the cell's membranes, and their profile changes owing to neurodegeneration. In this investigation, three agonists for distinct PG receptors were used to lower IOP and to determine their effect on the ON lipids. We utilized DBA/2J mice as a model of progressive IOP increase and C57BL/6J mice as a model of ON crush.
Methods:
DBA/2J and C57BL/6J mice were treated daily for 2 weeks with Latanoprost, PF-04217329, or Rivenprost. The IOP was measured every 2 days and pattern electroretinogram was conducted for DBA/2J throughout the study. Lipidomics of ONs were performed for each model and treatment group.
Results:
Of the tested compounds, Latanoprost and Rivenprost were the most effective agents decreasing IOP in DBA/2J mice. Triglyceride levels increased in the ONs of DBA/2J mouse model, but phosphatidylethanolamine levels underwent highest level changes in the C57BL/6J mouse model when treated with Latanoprost.
Conclusions:
Topical ocular FP- and EP4-receptor agonists appreciably lowered IOP in the DBA/2J mice representing pigmentary glaucoma. The observed changes in ON lipidomics in the different models of neurodegeneration suggest possible use of such measures in the development of more effective medicines for both IOP reduction and ON protection.
Introduction
Glaucoma is a debilitating eye disease characterized by the progressive loss of retinal ganglion cells (RGCs) and their axons resulting in serious visual impairment that can lead to irreversible blindness. Owing to the differences in etiologies linked with glaucoma, it is often regarded as a group of diseases, broadly categorized as open-angle glaucoma and angle closure glaucoma, with the former accounting for most cases in the United States. 1 The pathophysiology of glaucoma is not completely known, but current treatments strive to lower intraocular pressure (IOP), the leading risk factor and the only modifiable factor that slows down the disease progression. The ciliary body secretes aqueous humor (AH), which is dispersed in front of the iris and behind the cornea. 2
In glaucoma, the anterior eye chamber's production and drainage of AH become unbalanced, leading to an increase in IOP and subsequent damage to the optic nerve (ON). In open-angle glaucoma, AH drainage is reduced owing to the resistance at the trabecular meshwork (TM), whereas in closed-angle glaucoma, outflow is limited by iris apposition to the cornea resulting in TM obstruction. 3
Glaucoma research has focused on medications regulating IOP dynamics, with prostaglandin (PG) analogues serving as the first-line treatment options and retarding disease progression.4,5 Along with Travoprost and Bimatoprost, Latanoprost, a PG prodrug that is hydrolyzed to release the free acid active moiety, which binds to and activates the prostaglandin F (FP) receptor, is one of the most common ocular therapeutics utilized to treat ocular hypertension and glaucoma. Although FP receptor agonists are highly effective in reducing IOP, they can have considerable adverse effects such as hyperpigmentation of the iris and periocular skin, deepening of the upper eyelid sulcus, irregular elongation and thickening of eyelashes, and hyperemia. This has led to the quest for other therapeutically useful drugs, including agonists of other prostanoid receptors and their subtypes. 6
Under other circumstances, alpha-2-adrenergic agonists (e.g., brimonidine) and carbonic anhydrase inhibitors (e.g., brinzolamide) are utilized to lower IOP when the patient's IOP is recalcitrant to PGs or if the patient adversely reacts to the later drugs.6–8
Most of the early research on PG receptors in the eye has been conducted on cell cultures of various tissues, as opposed to in vivo animal studies that analyze both the anterior chamber tissues and the ON. The ON is composed of the axons of RGCs and is responsible for transmitting visual information from the retina to the brain. As part of the central nervous system (CNS), the axons of the ON are largely incapable of regeneration after injury in mammals. Clinically, the loss of axons at the optic nerve head (ONH) results in loss of visual acuity, which is central to glaucoma diagnosis. 9 In glaucoma, the membranes of ONH cells undergo a distinct perturbation, which manifests as altered lipid content.10,11 One of the goals of this study was to determine whether selective PG receptor agonists delivered by eyedrops to the anterior segment of the eye decrease IOP and simultaneously influence the lipid composition of the ON of mice.
In this study, the effects of three different PG receptor agonists on the reduction of IOP in two mouse models were compared. Latanoprost free acid is an agonist for FP receptors, whereas free acids of PF-04217329 and Rivenprost are agonists for EP2 and EP4 receptors, respectively. Although the FP-receptor mediates its biological actions by increasing phosphoinositide metabolism and generation of inositol phosphates (which cause release of intracellular Ca2+) and diacyglycerol (which act as second messengers), both the EP2 and EP4 receptor are positively coupled to adenylate cyclase and enhance intracellular cAMP level and use it as their second messenger. We wanted to compare the effects of these prostanoids that have different molecular and cellular mechanisms of action6,12 to better understand their role in modulating IOP in naive normal mice with and without optic nerve crush (ONC; C57BL/6J mice) and those with pigmentary dispersion glaucoma (DBA/2J mice).
The DBA/2J mouse is a well-described model for chronic angle-closure secondary glaucoma. 13 About 5% DBA/2J mice are found to retain open-angle, little to no pigment dispersion and lack of anterior segment. 14 However, this study evaluated a general cohort of DBA/2J mice cohort. Around 7–9 months of age, these mice spontaneously develop anterior segment defects, accompanied by a gradual increase in IOP. 15 Similar to glaucoma patients, DBA/2J suffer from ON degeneration and impaired eyesight. To supplement this study and gain a deeper understanding of the function of each compound, we used C57BL/6J mice after ONC. This model is frequently utilized in neuroregeneration research because it lacks the mutations present in DBA/2J. This study analyzes the effects of three PG analogs on IOP and the lipidome of the ON in two of the most widely used mouse models in ophthalmology. ON axonal transport has been reported to be impaired in glaucoma. This study also evaluated axonal transport for each treatment compared with untreated controls.
Methods
Animals and husbandry
All procedures were performed in compliance with the Association for Research in Vision and Ophthalmology (ARVO) statement for use of animals in ophthalmic and vision research. The experimental protocol was approved by the Animal Care and Use Committee of the University of Miami. A total of 80 DBA/2J mice and 60 C57BL/6J mice (Jackson Labs, Bar Harbor, ME), aged 8–10 months, were originally randomly assigned to the treatment groups with equal gender distribution. DBA/2J mice have been shown to have a gender difference in the time course of glaucoma development. To disregard any normotensive mice, we measured IOP previous to the beginning of the study and included only the animals with 20 mm Hg or above. 13
Mice were maintained in a cyclic light environment (12:12 h light [50 lux]–dark) and fed ad libitum. A group of C57BL/6J mice had ONC in one eye, as follows. Mice were first given an intraperitoneal injection (0.1 μl) of ketamine (100 mg/kg) and xylazine (9 mg/kg) provided by our veterinary division. Once confirmed to be fully anesthetized, one drop of 0.5% proparacaine hydrochloride was added to the eye that was going to be operated. A small incision was made in the conjunctiva at the superior region while carefully moving the eye forward until the head of the ON was observed. The nerve was crushed ∼1 mm from the globe with extra-fine self-closing forceps. Animals were allowed 2 days to recover and excluded from the study if experienced bleeding or unexpected damage.
The animals were treated with eyedrops containing either Latanoprost, PF-04217329 (Item No. PZ0369; Millipore Sigma, Burlington, MA), Rivenprost (Item No. 13618; Cayman Chemical, Ann Arbor, MI), or vehicle. Latanoprost is sold at a formulation of 50 μg/mL (0.005%). To establish dose consistency, the other two drugs were prepared in the same manner and at the same concentration.
For DBA/2J mice, eyedrops were given to both eyes, once daily for 14 days. Physical restrain rather than anesthesia was used to administer the drops on the days when no IOP or pattern electroretinogram (PERG) was performed. On the days of IOP and PERG measurements, the drops were given after the test between 11 AM and 12 PM to keep the schedule consistent. The C57BL/6J mice had the same schedule but the duration for IOP and PERG was 8 days, whereas the lipidomics cohort were treated for 14 days as the DBA/2J.
IOP and PERG measurements
To measure the IOP, a handheld tonometer (TonoLab, Colonial Medical Supply, Franconia, NH) was used. The mice were anesthetized, one at a time and placed in a separate cage until it failed to respond to touch before the measurements were taken. The anesthesia was administered by intraperitoneal injection (0.1 μl) of ketamine (100 mg/kg) and xylazine (9 mg/kg). To ensure the least variation, we measured IOP between 9 AM and 11 AM. The TonoLab was held by a clamp at the same height, and the mouse was placed on a platform that prevented any vibrations to compromise the results. Three IOP measurements were obtained, and a mean value calculated for each eye.
PERG was performed using a commercial PERG instrument (Jorvec Corp, Miami, FL). For PERG recordings, mice were anesthetized as previously described. The mouse was placed on a modified stereotaxic apparatus with a heating pad to keep body temperature at 37°C. The head was stabilized by using a mouth bite. The method used for recording is the same as described in a previous report. 16 In brief, two (15 × 15 cm) light emitting diode displays were placed in front of each eye (Jorvec Corp). The visual stimulations were black and white horizontal gratings with full contrast and different spatial frequencies (0.047–0.571 cycles/degree).
Multiple comparison analyses were conducted using analysis of variance (ANOVA) with Tukey's post hoc test or Dunnett's multiple comparisons test. Results with P < 0.05 were considered statistically significant.
Lipidomics
Extraction
Mass spectrometric lipidomics analysis was performed on a subset of ONs after 2 weeks of daily eyedrop treatment. The animals were killed and the entire globe with nerves was removed and separated into different vials. The nerves were subjected to lipid extraction per our previous protocols. 17 In brief, nerves were vortexed and sonicated in ultrasonic bath for 2 min in chloroform methanol and a deuterium-labeled standard, to rupture the membranes. Samples were vortexed again for 2 min and centrifuge at 17,800 g, 4°C for 30 min to obtain phase separation. The lower phase was collected and dried completely in a centrifugal vacuum concentrator.
Once ready for mass spectrometry, the stored lower phase was resuspended in acetonitrile. Another full range deuterium labeled standard was added to perform relative quantification. Standards used were GM1-d5, GM3-d5 and Equisplash™(Avanti Polar Lipids, Part of Croda International Inc., Alabaster, AL).
Mass spectrometry
Samples were analyzed by using liquid chromatography electrospray tandem mass spectrometry (LC-MS/MS). The instruments used were the Accela HPLC system and an orbitrap mass spectrometer (Q-Exactive, Thermo Scientific, Waltham, MA). An Acclaim 120 C18 3 μm column (Thermo Scientific) was run with methanol:water 60:40 (v/v) and 10 mM ammonium acetate and methanol:chloroform 60:40 (v/v) with 10 mM ammonium acetate, as solvent A and B, respectively. Each sample was run in both positive and negative ion mode.
Identification and quantification
The RAW scans were analyzed using LipidSearch (Thermo Scientific). Positive and negative scans were aligned, and the deuterium labeled standards were identified and allowed for normalization of each lipid class within the sample. MetaboAnalyst 5.0, a freely available software, was used for statistical analysis and the generation of figures.
Image analysis
ONs were collected for confocal imaging and immediately deposited in 4% paraformaldehyde at 4°C for overnight storage. The animal cohort for imaging was injected intravitreally with cholera toxin subunit B (CTB)–conjugated Alexa Fluor 488 (Thermo Fisher Scientific) 2 days before the animal was killed. 18 This is a prominent anterograde label for regenerated RGC axons. The following day, a sucrose gradient (5%–20%) was applied, followed by embedding in optimal cutting temperature compound (Tissue Tek, Sakura Finetek USA, Inc.) for cyrosectioning. Longitudinal sections (10 μm thickness) of the ON were placed on slides. A coverslip was mounted after 3 h. The VECTASHIELD Vibrance® Antifade mounting medium with DAPI or 4′,6-diamidino-2-phenylindole for nuclear staining was applied to the tissue section before sealing with coverslip. The imaging was performed with a Leica Stellaris 5 Confocal Microscope (Deerfield, IL). To facilitate intrasample comparison, the laser intensity and gain were held constant throughout the imaging.
Results
IOP measurements
This study examined the effectiveness of three PG receptor agonists in reducing IOP. Before beginning administration, the DBA/2J mice in the study were examined to determine if their IOP was >20 mm Hg. This was followed by daily eyedrops and every 2 days IOP readings. Latanoprost produced eye distress (hyperemia, swollen eyes) in ∼20% of the mice and these had to be eliminated from the studies because IOP could not be measured. This issue was not observed in mice treated with Rivenprost or PF-04217329. Both Latanoprost and Rivenprost displayed a comparable IOP lowering profile (Fig. 2A). IOP was reduced with PF-04217329 for the first 6 days, but then returned to baseline levels.
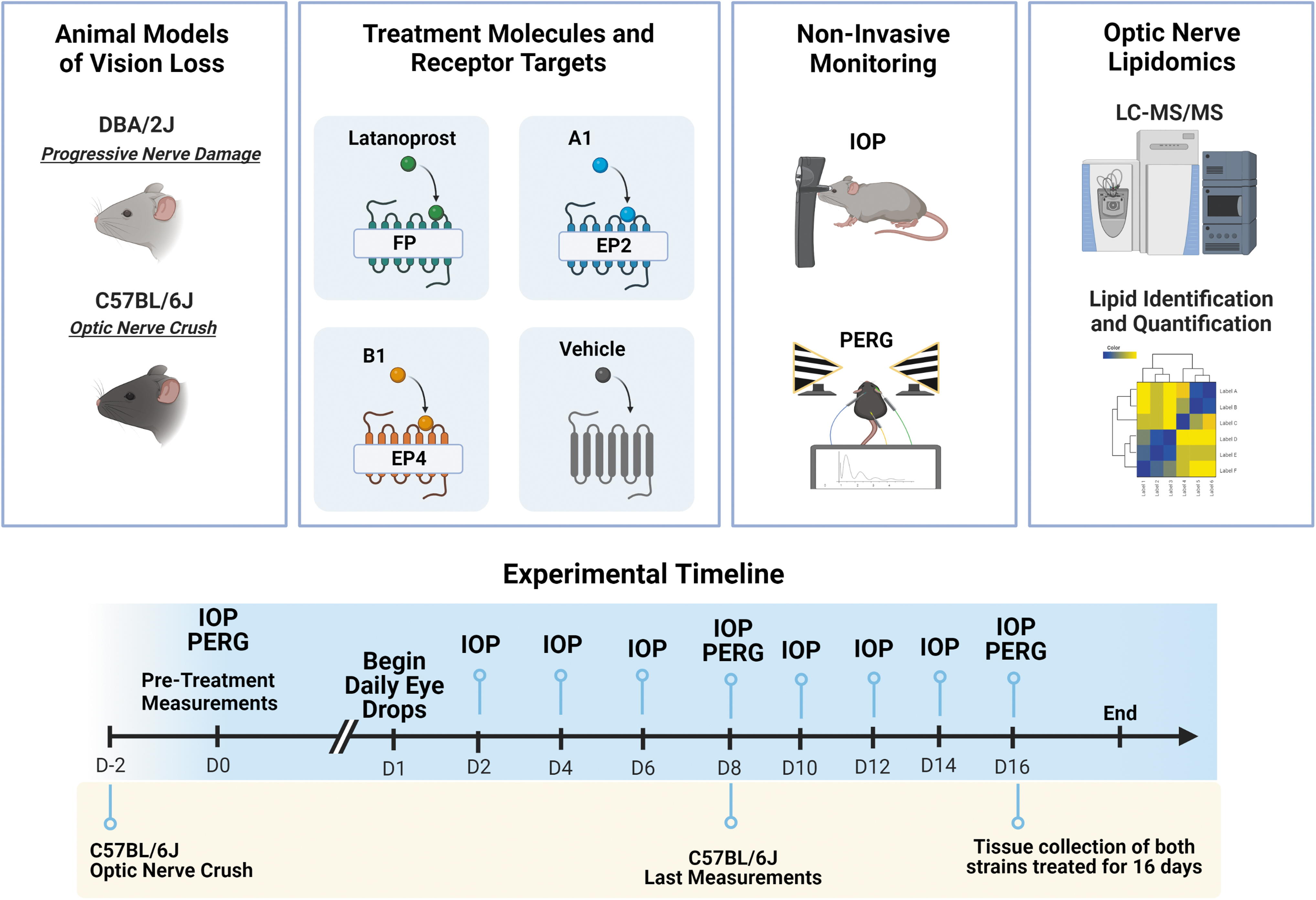
Schematic representation of the study overview. Two animal models were used, each treated daily for 2 weeks with one of three prostaglandin receptor agonists. The animals had their IOP and PERG measurements taken at different time intervals. The final analysis consisted of optic nerve lipidomics analyses and RGC axon regeneration imaging. Experimental timeline and measurement details have been described in the bottom. IOP, intraocular pressure; PERG, pattern electroretinogram; RGC, retinal ganglion cell.

DBA/2J mouse model responses to three prostanoids topical ocular treatment as indicated.
We also treated C57BL/6J animals with and without ONC for 8 days after ONC; the mice were given 2 days to recover. Following 2 days of therapy, the IOP of C57BL/6J rose and then recovered to the same level as the other groups (Fig. 3A). The noncrush control C57BL/6J mice exhibited an average IOP of 10–15 mm Hg during treatment (Fig. 3B).
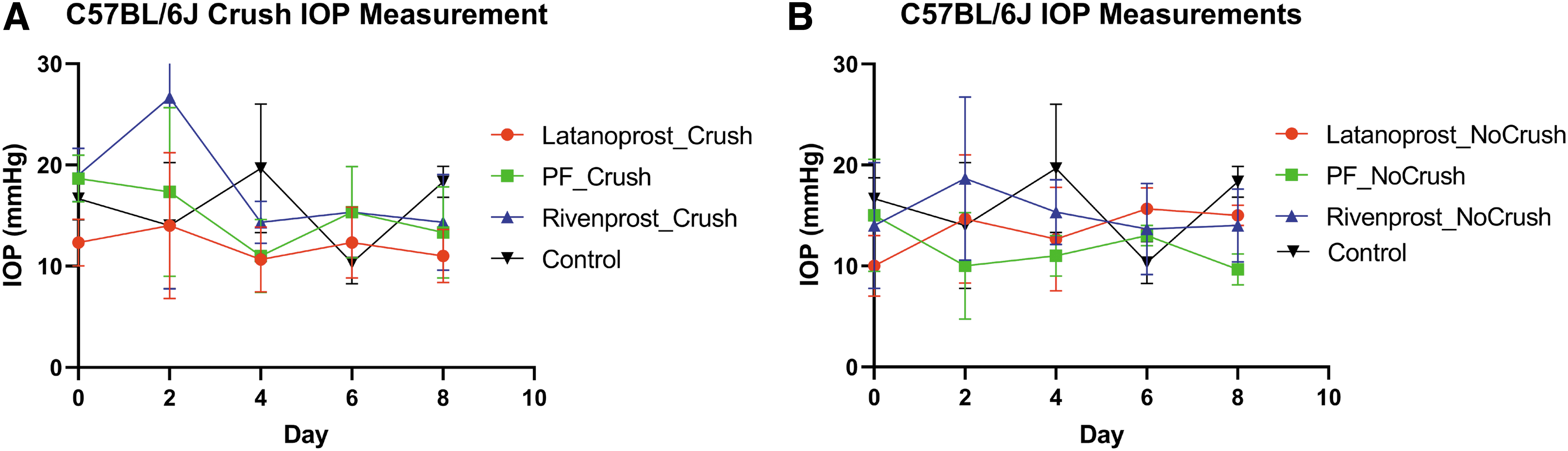
C57BL/6J mouse model intraocular pressure measurements before and during treatment with each of the prostaglandin analogs as indicated.
Pattern electroretinogram
To measure the effects of treatment on DBA/2J mice inner retinal function, we performed the noninvasive PERG analysis. This is an electrophysiologic ophthalmologic test that provides quantitative measurement of central retinal function. 16 PERG is the retinal response to a pattern-reversing, black-and-white checkerboard or stripped stimulus. We measured PERG in n = 6 mice, equal gender distribution, per treatment group. The PERG amplitude remained nonsignificant throughout the study (Fig. 2B).
ON lipidomics
All animals used for lipidomics had 2 weeks of daily eyedrop treatment. After killing, the eyes were dissected and stored at −80°C until analysis. For each category, three biological replicates were utilized. Figure 4A schematically provides the workflow utilized for analyses. After lipid identification and quantification with deuterium-labeled standards, the samples were further analyzed with the open-source software MetaboAnalyst. One-way ANOVA for multigroup analysis was performed for all treatment groups and both models. From the whole comparison, 46 of the 826 detected lipids were identified as significant (Fig. 4B).
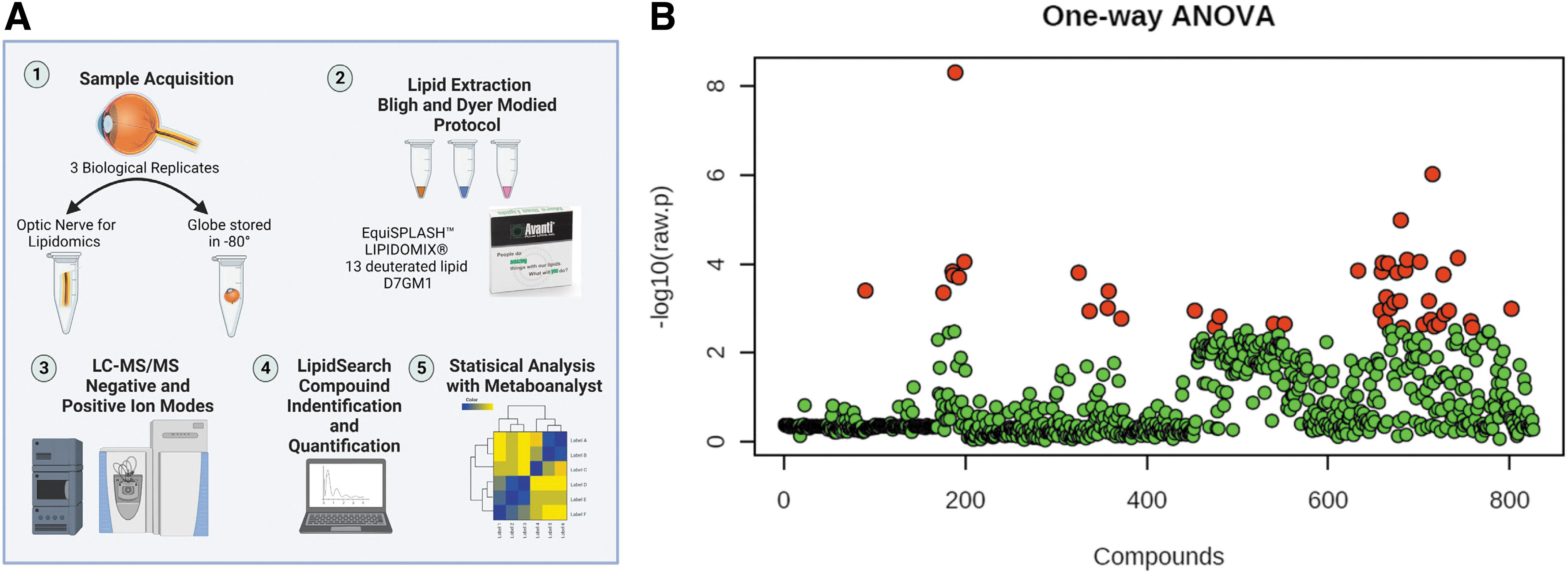
Overview of the lipidomics of all optic nerve samples.
Independent analyses of DBA/2J-specific lipidomics of the ON were conducted (Fig. 5). The partial-least squares discriminant analysis (PLS-DA) is a supervised approach that uses multivariate regression techniques to extract, via linear combination of the original variables (X), information that can predict class membership (Y). The scores plot explained 33.4% of the variance by treatment group and 24.3% of the variance by lipid species abundance (Fig. 5A). The correlation analysis of the individual lipids revealed that triglycerides (TG) are significantly elevated in the Latanoprost group, whereas Rivenprost had a minimal influence. Latanoprost and PF-04217329 exhibited parallel upregulation patterns in eight significant lipids, whereas Rivenprost exhibited the reverse pattern (Fig. 5B).
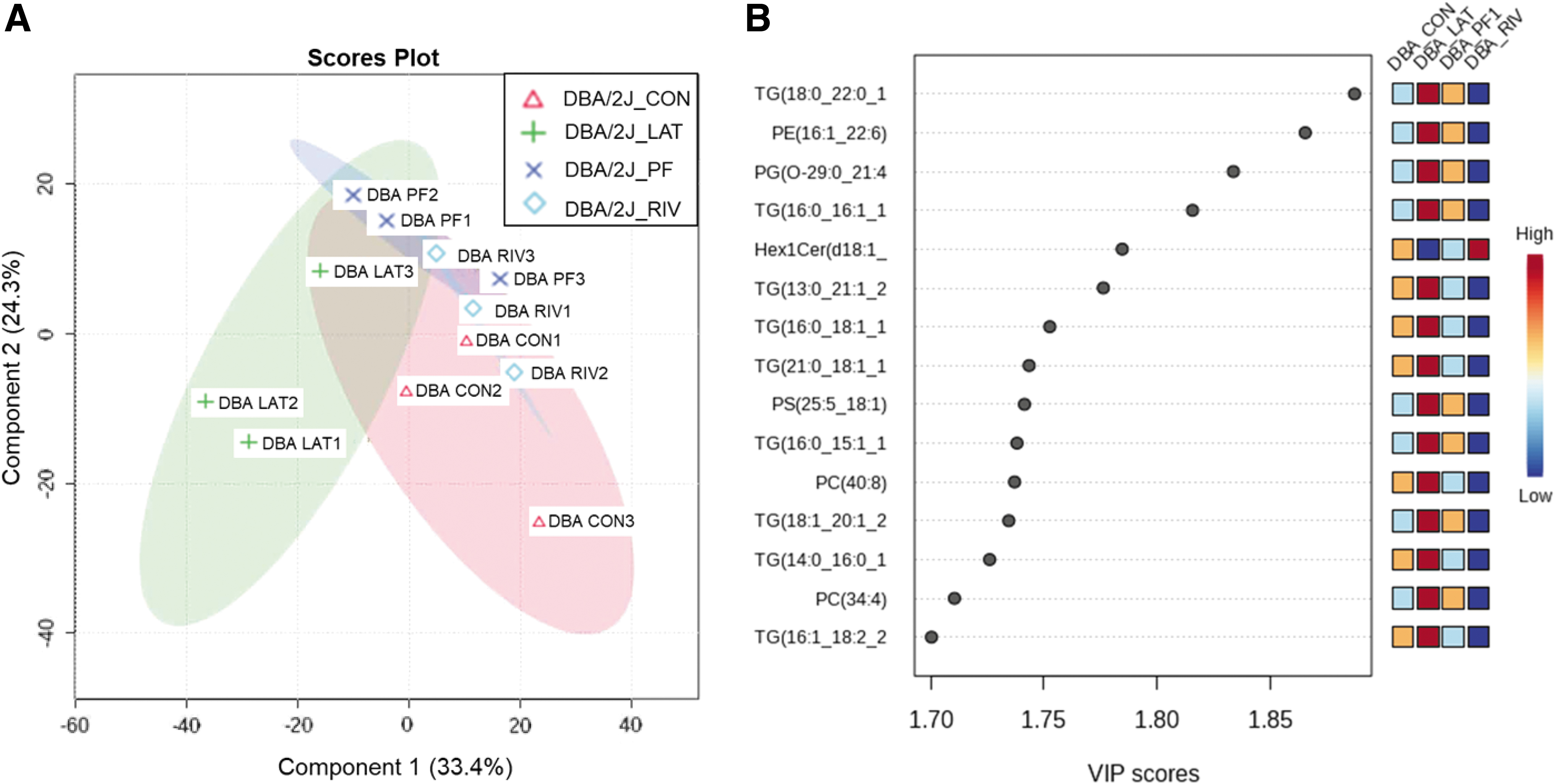
PLS-DA of DBA/2J optic nerve lipidomics.
The C57BL/6J samples were analyzed using the same techniques as the DBA/2J specimens. Using an ANOVA plot with a P-value cutoff of 0.05, two significant characteristics were identified. Hex2Cer(d18:1 14:0) and Hex1Cer(d18:2 24:1) were the species (Fig. 6A). The PLS-DA correlation revealed that the lipids of the control and Latanoprost were similarly elevated, whereas the lipids of Rivenprost and PF-04217329 were correspondingly downregulated. Phosphatidylethanolamine was unexpectedly the most significant class of phospholipids in C57BL/6J (Fig. 6B).
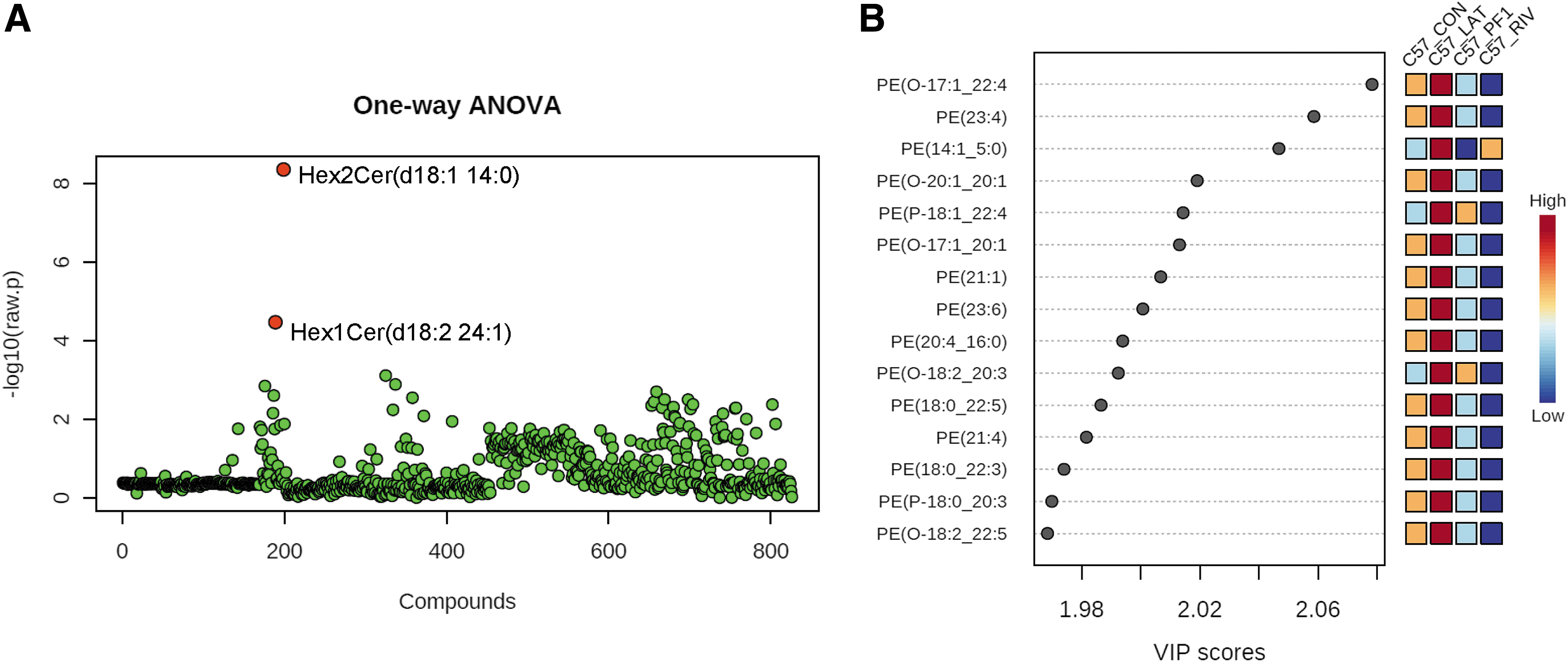
Analysis of C57BL/6J optic nerve lipidomics.
Comparison of both DBA/2J and C57BL/6J mice lipidomics revealed that both mice have similar lipid profile at baseline before any treatment (Fig. 7A). The topical ocular treatment with prostanoid analogs shifts the lipidome in a very distinct way among the two strains. This is where the genetic background of the models seems to play a role when metabolizing the medications (Fig. 7A). A study of gene expression in the brain tissue of both animal models (GSE4734) revealed that diacylglycerol kinase epsilon (Dgke) is substantially expressed in C57BL/6J mice but comparatively low in DBA/2J mice (Fig. 7B). This guided to a proposed schematic of how the genetic downregulation could have led to the high triacylglycerol found in the Latanoprost-treated DBA/2J (Fig. 7C).
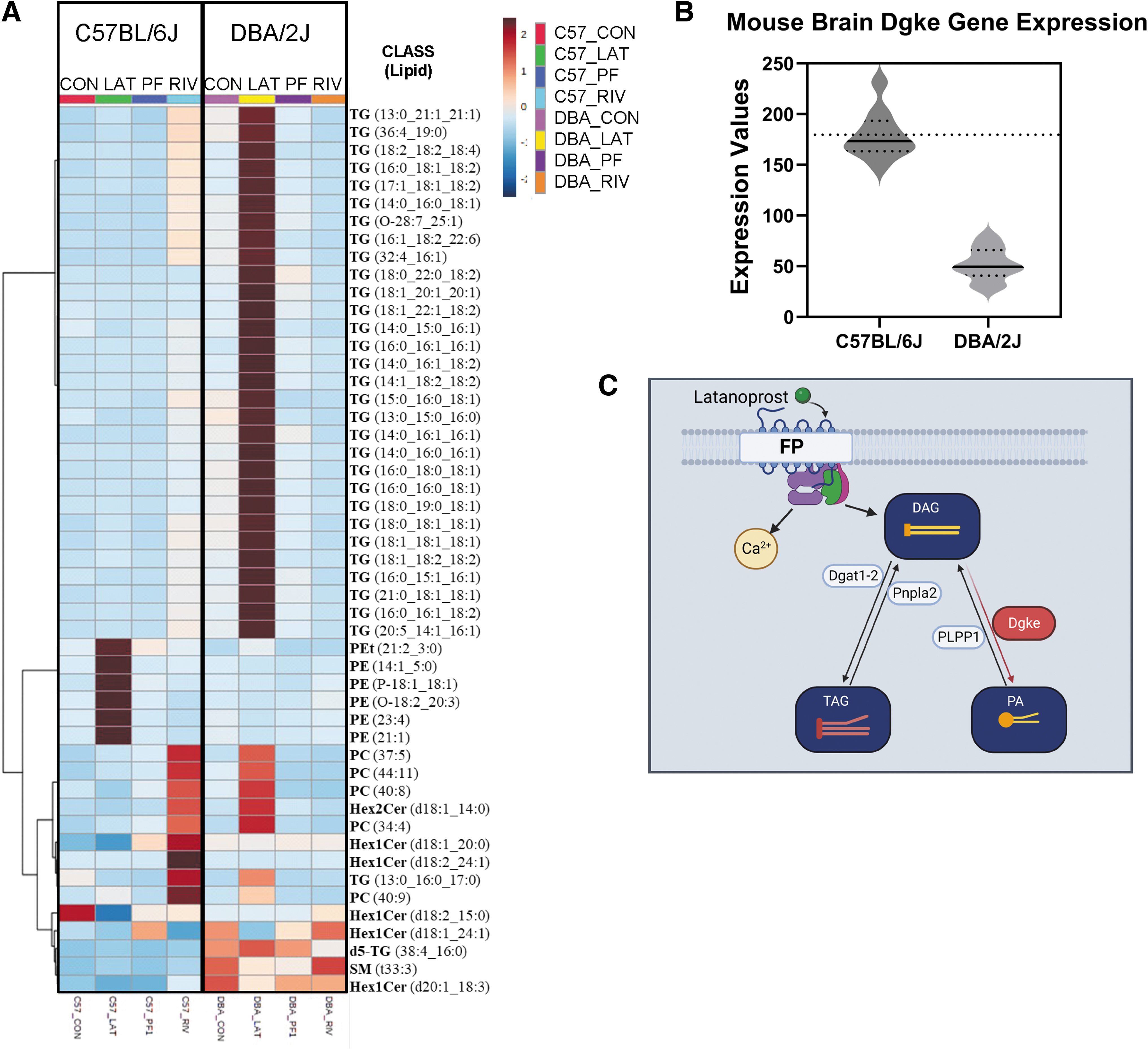
C57BL/6J and DBA/2J optic nerve lipidomics.
Axon analysis
To test the anterograde transport ability of the ONs, 2 μL of cholera toxin b-subunit (CTB) coupled with Alexa Fluor 488 was injected and imaged once the tissue was collected. CTB is also utilized as a marker of regeneration and lipid rafts because it binds to GM1, one primary component of growth cones. According to data given in Fig. 8, DBA/2J had the greatest overall transfer of CTB or regeneration (Fig. 8A). In this model, Latanoprost caused the most significant effect on axonal transport. C57BL/6J mice after ON crush did not show any axonal transport or axon growth/regeneration (Fig. 8B).
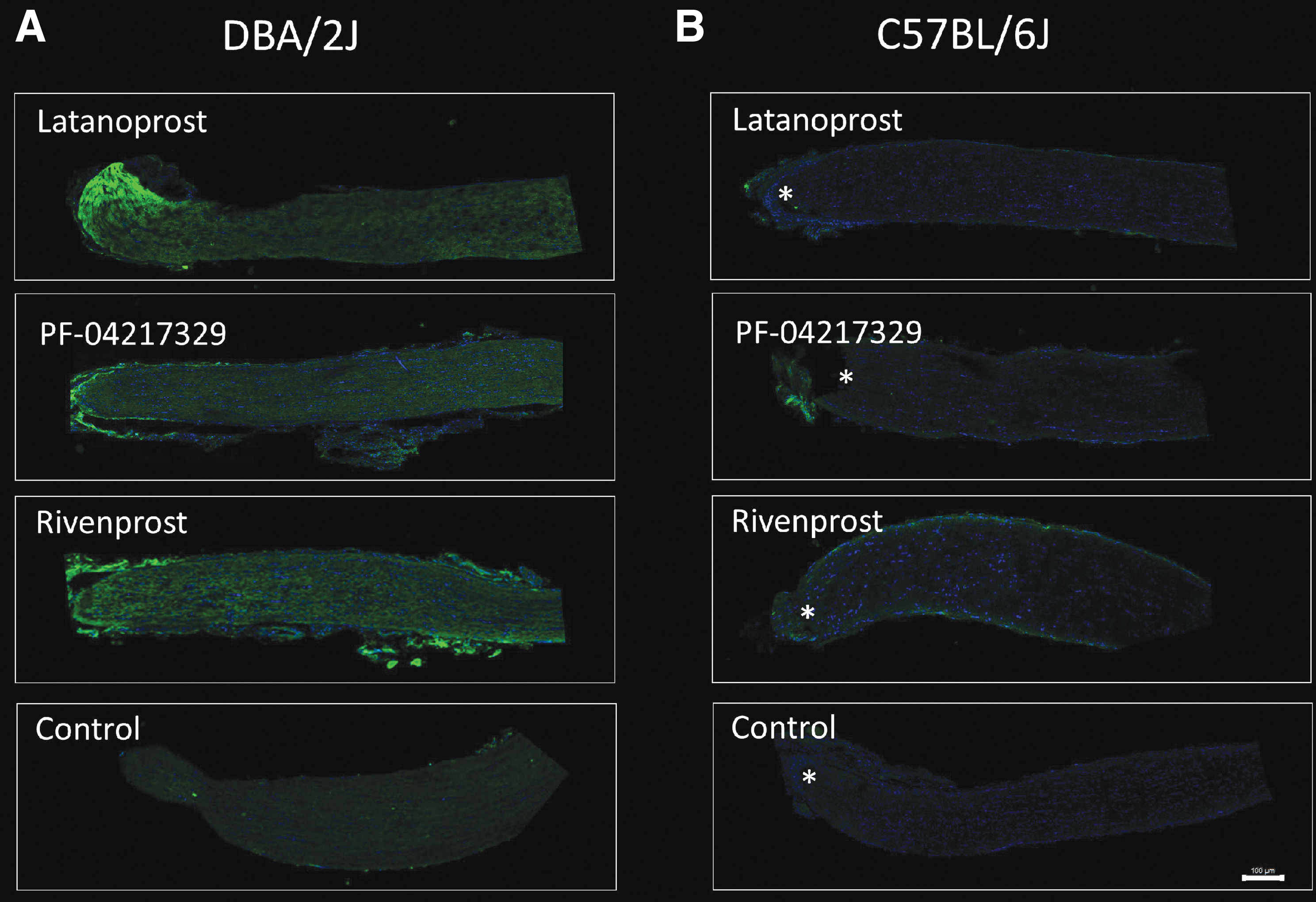
Axon transport analysis representative images of DBA/2J
Discussion
In this study, we used two mouse models to evaluate the capacity of three PG analogs to lower IOP and to identify their effects on the lipidome of the ON. The mice were treated with the clinically approved PG analog Latanoprost and two other PG analogs, Rivenprost and PF-04217329 prepared at the same 0.005% concentration (Fig. 1). Topical ocularly instilled Latanoprost is a commonly used drug to treat OHT and glaucoma, and its active free acid targets the PGF2a receptor (FP receptor). The free acid of Rivenprost is a small molecule agonist for the EP4 receptor, whereas PF-04217329 free acid targets and stimulates the EP2 receptor. 19 We have used the same concentrations for all the molecules to test the effect on IOP and lipidome of the ON. Further studies to establish the pharmacodynamics of these molecules would prove useful for translational purposes.
DBA/2J mice are a well-established model for chronic angle-closure secondary glaucoma, pigmentary dispersion glaucoma, 15 although a very small percentage of mice retain open-angle, normal anterior segment anatomy and little to no pigment dispersion. 14 Both males and females of the DBA/2J strain in our cohort developed glaucoma spontaneously between 8 and 10 months of age with an initial IOP of at least 20 mm Hg. 20 At this age, DBA/2J mice exhibit numerous physical and functional issues, prompting the withdrawal of 25% of the animals. 21 This colony displayed elevated levels of corneal calcification, neovascularization of the cornea and hyphema. Other research groups have observed the same problems and reported images of the complications. 22
At the midpoint of the research, Latanoprost and Rivenprost were the most effective drugs for reducing IOP in DBA/2J mice. On day 12, all treatments had increased IOP except for the Rivenprost group, whose IOP remained low until the end of the study (Fig. 2). Last measurement mean IOP for Rivenprost was 12 mm Hg (standard deviation [SD] = 5.4), Latanoprost was 13 mm Hg (SD = 4.2), PF-04217329 was 16 mm Hg (SD = 6.5), and the control was 17 mm Hg (SD 8.0). These results show the possibility of targeting the EP4 receptor to lower IOP in such pigmentary glaucoma conditions with Rivenprost, or as a combination with Latanoprost. However, these results need to be confirmed in additional studies.
To determine if the treatments had any effect on the functionality of RGCs, we utilized PERG amplitude measurement before the initial medicated eyedrops, at the midway, and at the conclusion of the testing paradigm. It is known that PERG is impaired in DBA/2J mice at an early stage of the disease, before observable axon loss occurs but while IOP rises steadily. Tested mice were at least 8 months old, an age at which the PERG amplitude is generally at baseline noise. 23 Our evaluation recorded similar finding, and none of the tested compounds were capable of restoring RGC function (Fig. 2B).
To broaden our understanding of PG-based therapy, we also utilized C57BL/6J mice, which lack the genetic alterations present in DBA/2J animals. The mice were 8–10 months old, and there was an equal distribution of genders. Axonal degeneration was induced by ONC in one eye. 18 The cohort with ONC had an increase in IOP 2 days after treatment started, which was more noticeable in the Rivenprost group (Fig. 3A). Rivenprost free acid is an agonist for the EP4 receptor, which activates adenylate cyclase and leads to an increase in the intracellular concentration of cyclic adenosine monophosphate (cAMP).
This activation of cAMP signaling can lead to the production of proinflammatory molecules. 24 Because the mice had surgery 4 days earlier, activation of the EP4 receptor could have caused further inflammation, leading to the abnormaly high IOP not seen in the ON noncrush mice (Fig. 3B). By the end of the study, all groups had IOP within the same ranges and there was no final difference in effect between ON crush and noncrush animals.
Despite FP-receptor PG agonist therapy in normotensive glaucoma patients, loss of RGCs and their axons at the ONH continues.9,19 The ONH is a delicate structure with the axon membrane experiencing distortion as the disease progresses. 15 Lipids are the most abundant component of the axon/cell's membrane, and their composition shifts in response to the onset of neurodegeneration.10,11 This prompted the question of whether the selected test drug eyedrops could modify the lipidome of the ONs of the various mice groups.
Three biological replicates were used for each group and model. One-way ANOVA of all the lipids identified, found 46 of the 826 total lipids to be significantly changed (Fig. 4). Analysis of the DBA/2J lipidome gave unique results where the Latanoprost-treated mice had a significant increase in triglyceride species, whereas Rivenprost had the opposite effect. PLS-DA indicated several TGs, one phosphatidylethanolamine (PE) and another phosphatidylglycerol species to be upregulated in Latanoprost- and PF-04217329-treated animals only (Fig. 5). It appears that once each drug treatment begins to impact the ON, the latter's metabolism appears to stimulate quite distinct pathways.
PLS-DA revealed that TG was insignificant in the lipid profile of C57BL/6J mice, resulting in a radically altered lipid profile. Latanoprost upregulated the most significant lipid class, PE, whereas PF-04217329 and Rivenprost downregulated it (Fig. 6). This could be related to the differences in the receptor coupling mechanisms and the second messengers associated with the FP receptor and the EP2 and EP4 receptors as mentioned previously.
When evaluating the heatmap of the combined mouse models, the most striking difference is how TG and PE increase from topical ocular Latanoprost treatment. Latanoprost is a synthetic PG analog in the form of a prodrug whose free acid has a high affinity for the prostaglandin FP receptor (FP receptor).5,25 The complicated interaction between PGs, their receptors, and lipid production and breakdown make it difficult to isolate precise mechanisms by which such changes are mediated. The exact manner by which Latanoprost increases TG in DBA/2J mice is not known. Because DBA/2J mice have many mutations that disrupt the biosynthesis of compounds, it is probable that a rise in TG is the result of an enzyme that cannot metabolize the cascade of changes caused by Latanoprost. PGs can be enzymically converted to arachidonic acid, which may serve as precursor for production of phospholipids or diacylglycerol (DAG). It appears that once the chemicals reach the nerve in DBA/2J, they are in the form of TG rather than phospholipids.
Activation of FP receptors has been shown to increase the levels of DAG in cells. 26 Comparison of gene expression in the CNS tissue of both mouse models (GSE4734) revealed a significant downregulation of Dgke. The protein coded by this gene, Dgke has substrate specificity toward arachidonoyl-DG to be converted to phosphatidic acid (PA). 27 A possible explanation for the high TG levels in DBA/2J ON is that the DAG to PA conversion is not as robust as in C57BL/6J mice (Fig. 7C). TGs are one of the main components of lipid droplets and are associated with axonal growth inhibition. On the contrary, PE is a major constituent of the cell membrane and is growth permissive. 28
The mice were injected with cholera toxin-B 488 to determine whether the lipid changes affected axonal transport or regeneration. Indicating the presence of lipid rafts, the DBA/2J mice displayed a stronger signal toward the ONH. Compared with the control group, Latanoprost and Rivenprost were the most active of the tested drugs. As a result of ONC, the C57BL/6J mice did not exhibit any fluorescence changes reflecting no change in axonal transport or axon regeneration. Increased IOP has been shown to cause deformation of the ONH and axonal transport deficiency in different animal models. The lack of nutrients passing through the axons could lead to the observed lipid changes in the ONs of the different mice.29,30
Our findings indicate the significance of evaluating ocular medications that mediate their actions via different molecular and cellular mechanisms of action using multiple animal models. Our study revealed that ocular hypotensive medications given topically to the eye do have an effect on the ONH and ON and adjacent tissues. These compounds were able to alter the lipids on the nerve despite being administered as a drop in the anterior chamber. When Latanoprost was administered to DBA/2J mice, the nerves contained elevated levels of TG, but C57BL/6J animals exhibited an increase in PE. Shifts in the nerve lipidome may be responsible for the RGCs' incapacity to renew, and this should be taken into account while prescribing glaucoma drugs and evaluating other therapies for the eye.
In addition, the role of PG receptor activation on lipid metabolism may influence the development of more effective medicines for both IOP reduction and ON protection. Small molecules are typically simpler to deliver and safer than gene therapies. Further research with these altered molecules and lipids could lead to the creation of a combination therapy for IOP and ON regeneration. We hope that future research in this arena will shed further light on the link between IOP changes, lipid metabolism, and ON (RGC axon) regeneration.
Footnotes
Authors' Contributions
J.A.: Experimentation, Data curation, Writing—original draft preparation. A.E.: Animal experimentation, Data acquisition. N.A.S.: Compound selection, Article preparation, Data interpretation. S.K.B.: Conceptualization, Methodology, Writing—reviewing and editing.
Disclaimer
Funders had no role in experimental design and overall interpretation of data.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a research agreement grant from Santen Inc., Research to Prevent Blindness (RPB), NIH grants EY031292 and EY14801.