
Abstract
Purpose:
To examine the effects of prostanoid FP and EP2 receptor agonists, PGF2α and Omidenepag (OMD), respectively, on the transforming growth factor beta (TGF-β2) induced conjunctival fibrogenesis.
Methods:
Two-dimension (2D) and three-dimension (3D) cultures of these fibroblasts were subjected to following analyses: (1) planar proliferation evaluated by transendothelial electron resistance (TEER) measurements, (2) real-time metabolic analyses, (3) subepithelial proliferation evaluated by 3D spheroid' size and stiffness measurements, and (4) the mRNA expression of extracellular matrix (ECM) molecules and their modulators.
Results:
TGF-β2 induced increase in the planar proliferation was significantly decreased or enhanced by PGF2α or OMD, respectively. The proportion of oxygen consumption required to drive ATP synthesis compared with that driving proton leakage was increased by PGF2α and OMD independently with TGF-β2. In contrast, maximal mitochondrial respiration was decreased by PGF2α and OMD, and the OMD-induced effect was further enhanced by the presence of TGF-β2. In addition, the TGF-β2 dependent increase in the glycolytic capacity was cancelled by PGF2α and/or OMD. Alternatively, subepithelial proliferation, as evidenced by the stiffness of the 3D spheroids, was substantially increased by both PGF2α and OMD, and these were differently modulated by TGF-β2. The expression of several related factors as above fluctuated among the conditions for both 2D and 3D and TGF-β2 untreated or treated cultures.
Conclusion:
The present findings indicate that the prostanoid FP or the EP2 receptor agonist may solely and differently induce the planar and subepithelial proliferation of HconF cells and these were also modulated by TGF-β2.
Introduction
The human conjunctiva functions to serve as a biological protecting barrier as well as to maintain the ocular surface homeostasis. 1 In a pharmacological point of view, the permeability beyond these conjunctival barriers is also known to be a critical factor for drug delivery to the posterior segments of the eye upon the instillation of several ocular drugs. 2 Alternatively, these conjunctival barrier functions can be impaired in cases of several ocular surface diseases as well as surgical intervention leading to both planar and subepithelial fibrogenetic changes. Therefore, the regulation of these conjunctival fibrosis during their wound healing process in the conjunctiva is of great importance to obtain the preferable surgical outcomes of ocular surface-related diseases, especially glaucoma filter surgery.3–7 In fact, uncontrolled conjunctival scarring at the operative site may lead to poor success results in the trabeculectomy.8,9
During the physiological and pathological wound-healing processes, it is well known that several cytokines and growth factors are importantly involved, and among these, transforming growth factor beta (TGF-β) regulates almost all processes of wound healing. 10 In fact, these TGF-β11,12 induced conjunctival wound healing responses are caused by 2 different processes; that is, (1) superficial re-epithelialization and wound contraction, and (2) subconjunctival fibrous scar formation. 13
Therefore, upon exposure to TGF-β, the transdifferentiation of the fibroblasts, the major sources of extracellular matrix (ECM) proteins, into myofibroblasts can be evoked.14–16 During the normal healthy wound repair process, such myofibroblasts are removed from the wound area by apoptosis. However, in case that such wound repairing process does not work, progressive fibrosis by myofibroblasts may cause permanent scar formation.14,17 Therefore, the strategy to prevent such progressive conversion of fibroblasts to myofibroblasts should be extremely important for maintaining appropriate barrier functions of conjunctiva.8,18,19
A characteristic optic nerve head damage called “glaucomatous optic neuropathy (GON)” is defined to be associated with the progressive visual field deterioration upon elevated levels of intraocular pressure (IOP).20,21 As possible mechanisms responsible for such elevated levels of IOPs, increased out flow resistance of aqueous humor, presumably due to excess levels of deposits of ECM proteins within the trabecular meshwork.22–24 Therefore, hypotensive therapy to decrease IOP at suitable levels by antiglaucoma medication, laser treatment or surgery is currently the only evidence-based strategy for the treatment of GON. 25 Among initial antiglaucoma medications, prostaglandins derivatives (PGs), prostanoid FP receptor agonists are frequently used as first-line antiglaucoma medications due to their substantial hypotensive efficacy with minimal systemic side effects.26–28
However, recently, within a considerable number of long-term patient users of FP receptor agonists, local adverse effects so-called “prostaglandin-associated periorbitopathy (PAP) which includes deepening of the upper eyelid sulcus (DUES)” have been recognized.29,30 For alternative option to avoid such adverse effects by FP receptor agonists, a selective prostanoid EP2 agonist prodrug, Omidenepag isopropyl (OMDI), has been used in the treatment of patients with ocular hypertension (OH) and glaucoma.31,32 In fact, our recent studies using in vitro three-dimensional (3D) spheroid models replicating DUES etiology suggested that EP2 agonist, Omidenepag (OMD), but not FP receptor agonists, may not induce DUES.33,34
Thus, these findings rationally suggest that topically used FP and EP2 receptor agonists may differently affect orbital fatty tissues. Since such topically administrated drugs should be transported thorough ocular surface including conjunctiva, both drugs may also affect the physiological and pathological states of conjunctival tissues. In fact, previous studies indicated that both receptors are expressed within conjunctival tissues.35,36 Furthermore, since both antiglaucoma drugs are also frequently used before and after glaucoma filter surgery, it is of great interest to know whether or not both drugs affect wound healing processes within conjunctiva. However, as of this writing, our knowledge regarding the drug-induced effects of FP and EP2 receptor agonists toward conjunctival fibrogenesis remains quite limited.
Therefore, in this study, to obtain insights into this issue, we examined the effects of the FP receptor agonist, PGF2α and the active form of EP2 agonist, OMD on two-dimensional (2D) and 3D human conjunctival fibroblasts (HconF) cell cultures that had been treated and untreated with TGF-β2. These cultures were previously shown to be suitable in vitro conjunctival models for fibrogenetic changes on the plane or in the spatial space, respectively. 37
Methods
2D and 3D cultures of HconF
HconF cells purchased from ScienCell Research laboratories (Carlsbad, CA) were 2D cultured and subjected to the planar proliferation analysis by transendothelial electron resistance (TEER) measurements and the measurement of real-time cellular metabolic functions as described hereunder. In brief, 2D cultures of HconF cells were cultured in 150 mm 2D culture dishes at 37°C in the Fibroblast Medium (FM; Catalog No. 2301, ScienCell Research laboratories). 37 Alternatively, they were further maintained by changing the medium every other day or subjected to 3D spheroid cultures, as described in our previous study. 37
In brief, 2D cultured HconF cells were washed with phosphate buffered saline (PBS), detached by treatment with 0.05% Trypsin/EDTA, resuspended in the FM supplemented with 0.25% methylcellulose (Methocel A4M) at a level of ∼20,000 HconF cells in 28 μL, and each well was subjected to hanging drop culture plates (No. HDP1385; Sigma-Aldrich, St Louis, MO) (Day 0). On each subsequent day until Day 6, half of the medium (14 μL) was exchanged with fresh medium.
For evaluating drug-induced effects, 5 ng/mL solutions of TGF-β2, 500 nM solutions of PGF2α or OMD, or both were added to 2D or 3D cultured Hcon F cells daily for 6 days starting on day 1. The dosages of the TGF-β2, PGF2α, and OMD used in this study were based on data reported in a previous study.37,38
TEER of 2D cultured HconF cells
The TEER values for HconF cell monolayers were determined as described in a previous study.37,39 In brief, HconF cells prepared in 150 mm 2D cultured dishes as aforementioned were washed with a PBS, and the cells were detached using 0.25% Trypsin/EDTA. After centrifugation for 5 min at 300 g, the cell pellet was resuspended in the FM and HconF cells were seeded on 12-well plates for TEER (0.4 μm pore size and 12 mm diameter; Corning Transwell, Sigma-Aldrich) at a density of 2.0 × 104 cells per well. In each well of the TEER plate, the apical side (inside of the membrane inserts) and basal side (outside of the membrane inserts) were maintained in 0.5 and 1.5 mL of the FM, respectively.
When cells had reached ∼80% confluence, 5 ng/mL TGF-β2 was added to the FM of the apical side in the absence or presence of 500 nM PGF2α or OMD (Day 1). This culture medium of the apical side in each experimental group was changed every other day. At Day 6, TEER (Ωcm2) was measured using an electrical resistance system (Kanto Chemical Co. Inc., Tokyo, Japan) according to the manufacturer's instructions after washing twice with PBS.
Measurement of real-time cellular metabolic functions
The rates of oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) of 2D HconF cells were simultaneously measured using a Seahorse XFe96 Bioanalyzer (Agilent Technologies) as described previously with minor modifications.40,41 In brief, 20 × 103 2D HconF cells were placed in wells of a 96-well assay plate as follows: (1) nontreated control (CON), (2) treated with TGF-β2 (TGF), (3) treated with PGF2α or OMD, and (4) treated with TGF-β2, and PGF2α or OMD. After replacing the culture medium with Seahorse XF Dulbecco's modified Eagle medium assay medium (pH 7.4, No. 103575-100; Agilent Technologies) supplemented with 5.5 mM glucose, 2.0 mM glutamine, and 1.0 mM sodium pyruvate, the baseline OCR and ECAR values were determined using a Seahorse XFe96 Bioanalyzer.
The samples were further analyzed after supplementation with 2.0 μM oligomycin (complex V inhibitor), 5.0 μM carbonyl cyanide p-trifluoromethoxyphenylhydrazone (FCCP, a protonophore), 1.0 μM rotenone/antimycin A (complex I/III inhibitors), and 10 mM 2-deoxyglucose (a hexokinase inhibitor).
Measurement of the size and stiffness of 3D HconF spheroids
The physical properties, size, and hardness of the HconF 3D spheroids were characterized as reported in our previous studies.42,43 In brief, the mean sizes of the 3D spheroids were measured using an inverted microscope (Nikon ECLIPSE TS2; Tokyo, Japan). Alternatively, for the hardness measurement, a single living 3D spheroid was placed on a 3 × 3-mm plate and compressed until a 50% deformation was achieved during 20 s using a micro-compressor (MicroSquisher, CellScale, Waterloo, ON, Canada). The force required (μN) was determined, and force/displacement (μN/μm) was calculated.
Other analytical methods
As previously reported,44,45 total RNA was extracted from 2D or 3D HconF cells using an RNeasy mini kit (Qiagen, Valencia, CA) was then subjected to reverse transcription using the SuperScript IV kit (Invitrogen) as per the manufacturer's instructions. Each respective gene expression was quantified by real-time polymerase chain reaction (PCR) with the Universal Taqman Master mix using a StepOnePlus machine (Applied Biosystems/Thermo Fisher Scientific) using specific primers and probes (Supplementary Table S1). cDNA quantities were normalized to the expression of 36B4 (Rplp0) and are shown as fold-change relative to that for the control.
Experimental data are shown as the arithmetic mean ± the standard error of the mean. All statistical analyses were performed using GraphPad Prism 8 (GraphPad Software, San Diego, CA) as described in a recent report.44,45 Statistical difference between experimental groups were determined by analysis of variance followed by a Tukey's multiple comparison test.
Results
Effects of PGF2α and OMD on the planar proliferation and metabolic properties of 2D cultured HconF cells.
Using TGF-β2 treated 2D and 3D cultured HconF cells, we recently developed suitable in vitro models that mimic the TGF-β2-related myofibroblast-induced changes on the surface plane as well as in the subepithelial spatial space, respectively. 37 In this study, we examined the effects of 500 nM PGF2α or OMD on these models. Before this study, the expression of receptors for PGF2α and EP2 within these HconF preparations was confirmed (Fig. 1).
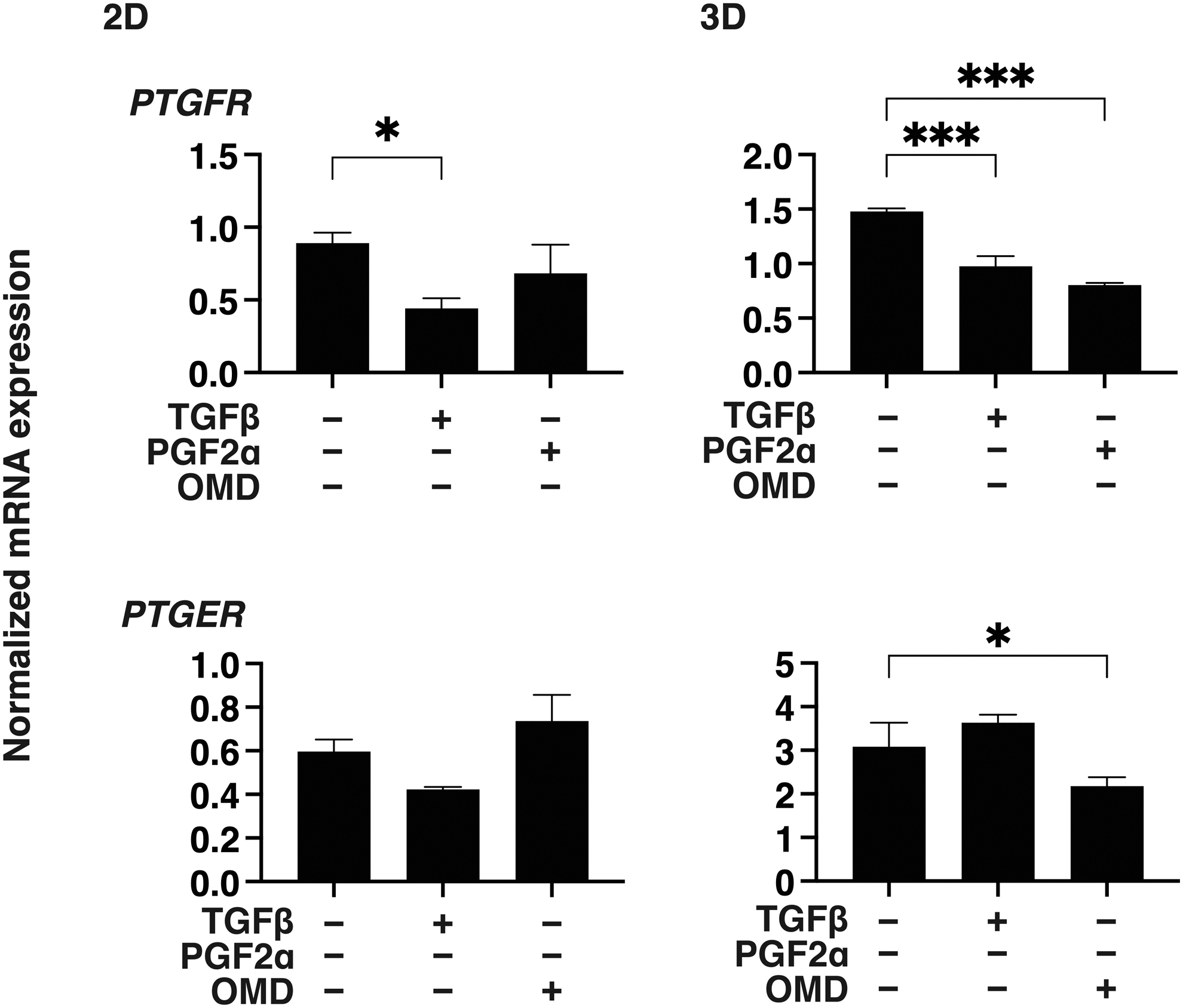
mRNA expression the FP or the EP receptor in 2D and 3D cultured HconF cells. 2D or 3D cultures HconF cells were subjected to qPCR analysis to estimate the mRNA expression of FP (PTGFR) or EP2 (PTGER) receptor. All experiments were performed in duplicate using fresh preparations. *P < 0.05, ***P < 0.005. 2D, two-dimensional; 3D, three-dimensional; HconF, human conjunctival fibroblasts qPCR, quantitative polymerase chain reaction.
To study these drug-induced effects on planar proliferation, the TEER measurement of 2D HconF monolayers in the presence of and in the absence of TGF-β2 were evaluated. As shown in Fig. 2, a significant increase in TEER values were observed upon an exposure of the cultures to TGF-β2. PGF2α or OMD did not change TEER values alone. However, and quite interestingly, both drugs caused different effects toward TGF-β2 treated HconF cell monolayers. That is, TGF-β2-induced increase in TEER values was decreased in the presence of PGF2α but not OMD, and in turn, the OMD but not PGF2α treated TEER values were significantly increased by TGF-β2.

Effects of PGF2α or OMD on TEER of TGF-β2 untreated or treated 2D HconF monolayers. The 2D HconF cell monolayer was untreated or treated with a 5 ng/mL solution of TGF-β2 in the absence or presence of 500 nM PGF2α or OMD. The 2D cultures of HconF monolayers at Day 6 were subjected to planar proliferation analyses by TEER electric resistance (Ωcm2) measurements. All experiments were performed in triplicate using fresh preparations. *P < 0.05, **P < 0.01, and ****P < 0.001. OMD, Omidenepag; TEER, transendothelial electrical resistance; TGF-β2, transforming growth factor beta 2.
The effects of PGF2α or OMD on the cellular metabolic properties of the TGF-β2 untreated or treated 2D HconF cells were also examined (Fig. 3). The coupling efficiency, defined as the ratio of oxygen consumption required for driving ATP synthesis to that driving proton leakage, of both TGF-β2 untreated and treated 2D cells was increased by PGF2α and/or OMD. This suggests that the oxygen consumption required for ATP synthesis is increased by PGF2α and OMD independent of TGF-β2, although maximal mitochondrial respiration was decreased by PGF2α and OMD. In contrast, the increase in glycolytic capacity induced by TGF-β2 was canceled by both PGF2α and OMD irrespective of whether TGF-β2 was present or not.
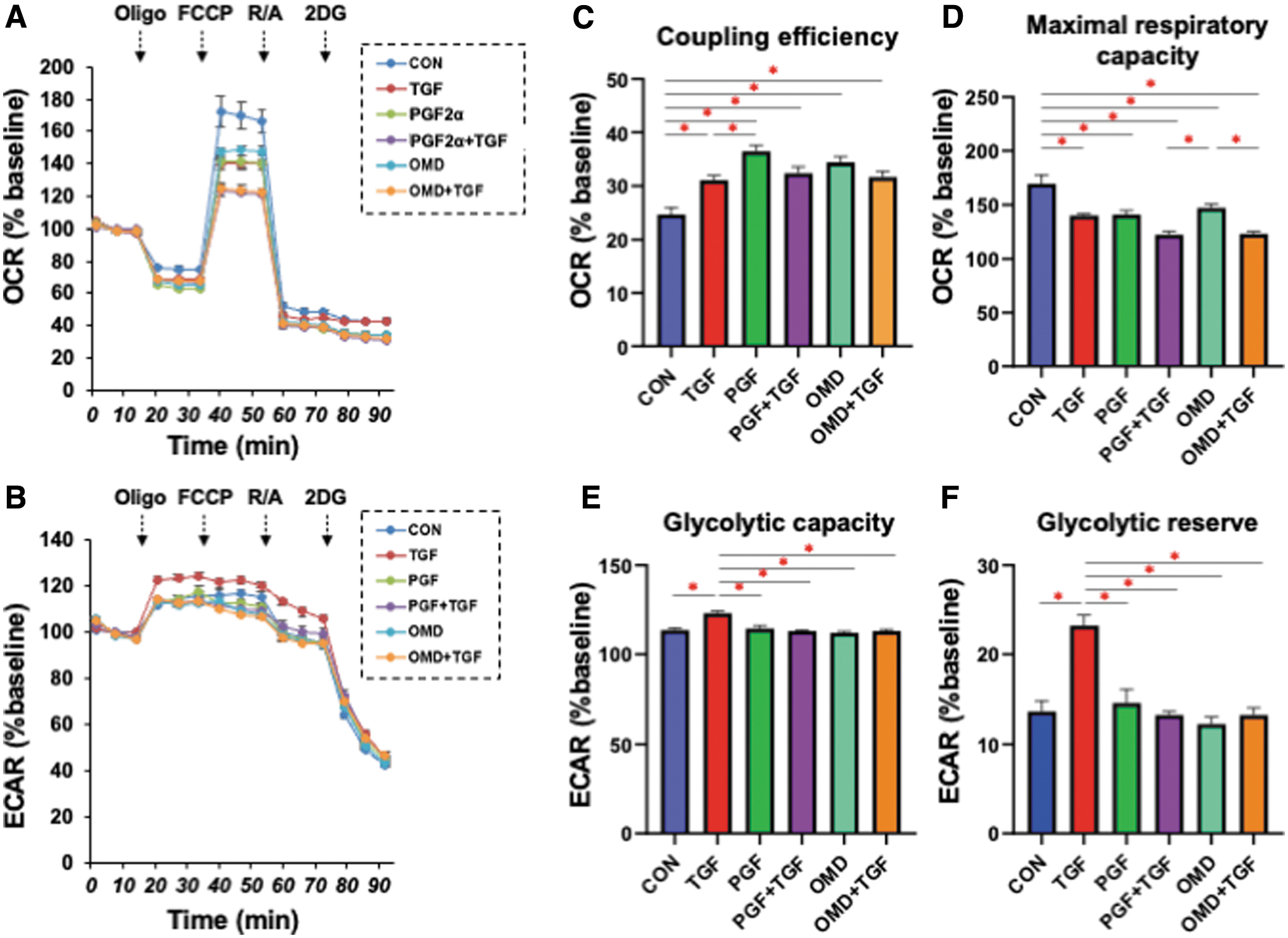
Effects of PGF2α or OMD on the cellular metabolic phenotype of 2D HconF cells in the absence or presence of TGF-β2. In addition to an untreated control (CON), 2D HconF cells were untreated or treated with a 5 ng/mL solution of TGF-β2 (TGF) in the absence or presence of 500 nM PGF2α (PGF) or OMD. At Day 6, the samples were subjected to a real-time metabolic function analysis using a Seahorse XFe96 Bioanalyzer. Values of OCR
These results indicate that PGF2α and OMD significantly affect the TGF-β2-induced planar proliferation of the 2D HconF cell monolayers in different manners and that these effects may be associated with alterations in mitochondrial respiratory capacity.
Effects of PGF2α or OMD on the fibrogenetic properties of 3D HconF spheroids.
To examine the PGF2α- or OMD-induced effects on myofibroblast-induced changes in the subepithelial spatial space, the physical characteristics, including the size and stiffness of the TGF-β2 untreated or treated 3D HconF spheroids were evaluated. As shown in Fig. 4, although the mean sizes of the 3D HconF spheroids were not changed significantly by TGF-β2 and/or PGF2α or OMD, the physical stiffness of the 3D HconF spheroids was significantly increased by a monotreatment with PGF2α or OMD, and PGF2α also enhanced TGF-β2-induced effects. These results indicate that PGF2α and OMD significantly facilitate the TGF-β2-unrelated myofibroblast-induced changes in the subepithelial spatial direction, although these changes are modulated differently by TGF-β2.
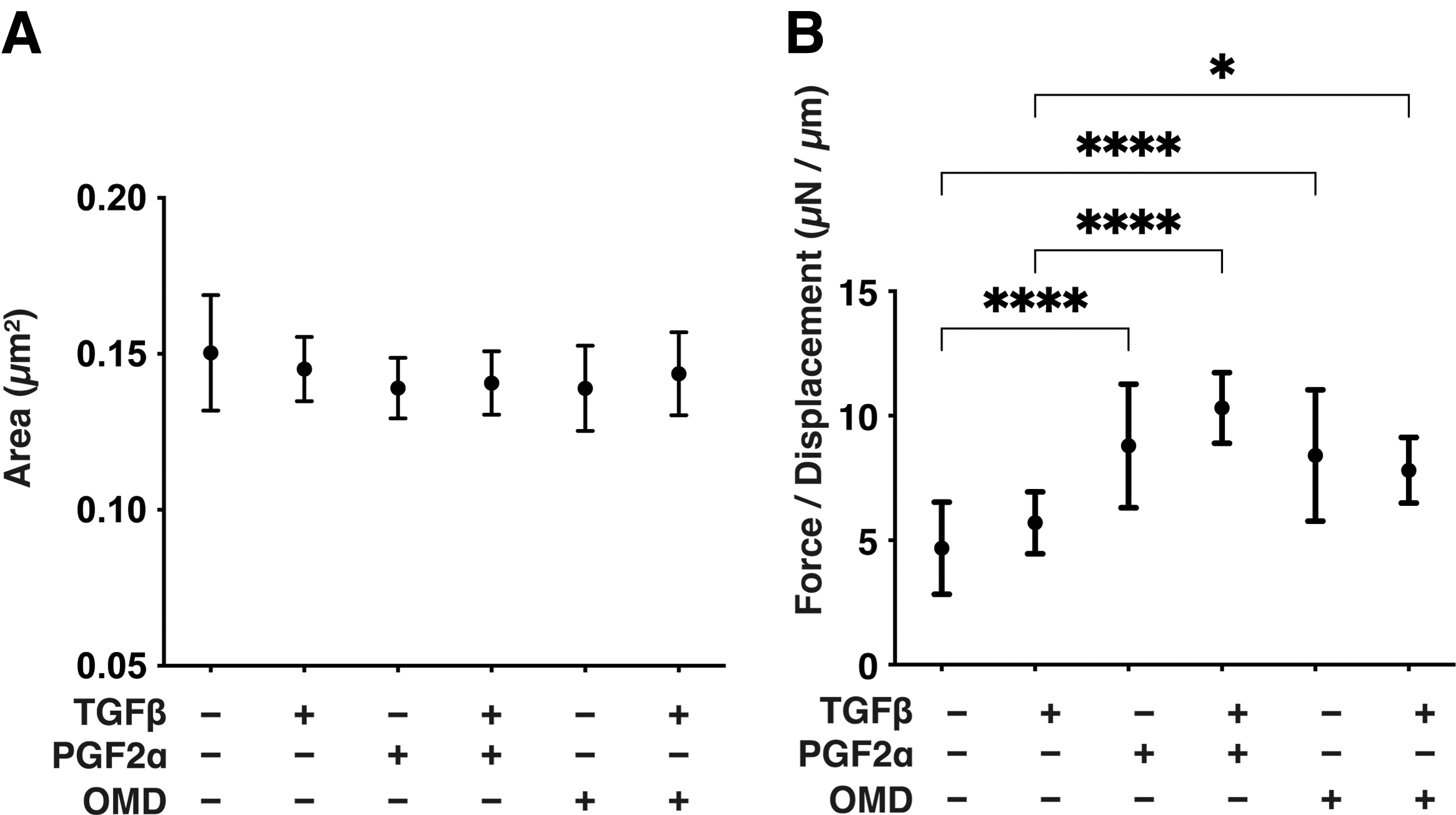
Effects of PGF2α or OMD on the physical properties, sizes, and stiffness of 3D HconF spheroids in the absence and presence of TGF-β2. 3D HconF spheroids were untreated or treated with a 5 ng/mL solution of TGF-β2 in the absence or presence of 500 nM PGF2α or OMD. At Day 6, the mean sizes were measured and the plotted data are shown in panel A. The physical solidity was analyzed by a micro-squeezer, and the force required to produce a 50% deformity of a single spheroid during a period of 20 s was plotted in
Effects of PGF2α or OMD on the expression of several ECM proteins, their modulators, and ER stress-related factors on 2D and 3D cultured HconF cells
To study further these issues in more detail, the PGF2α or OMD-induced effects on the expression of some major ECM proteins, including collagen1 (COL1), COL4, COL6, fibronectin (FN), and α smooth muscle actin (αSMA) of TGF-β2 untreated or treated HconF cells, were determined by qPCR analysis (Fig. 5). A monotreatment of TGF-β2 but not PGF2α- or OMD-induced significant upregulations in COL1, COL4, and αSMA and the addition of TGF2α to PGF2α or OMD caused substantial downregulations of COL6 in the 2D HconF cells. Whereas in contrast TGF-β2 alone had no affect, but PGF2α or OMD induced relative or significant upregulations of all 5 ECMs independently with TGF-β2 in the 3D HconF spheroids.

Effects of PGF2α or OMD on the mRNA expression of ECMs in 2D and 3D cultured HconF cells in the absence and presence of TGF-β2. 2D and 3D HconF cells were untreated or treated with a 5 ng/mL solution of TGF-β2 in the absence or presence of 500 nM PGF2α or OMD. At Day 6, each sample was subjected to qPCR analysis and the expression of mRNA in ECMs, COL1, COL4, COL6, FN, and aSMA were estimated. All experiments were performed in duplicate using 3 different confluent 6-well dishes (2D) or 15 freshly prepared 3D HconF spheroids (3D) in each experimental condition. *P < 0.05, **P < 0.01, and ***P < 0.005. ECM, extracellular matrix.
Similarly, the PGF2α or OMD induced effects on the mRNA expressions of ECM modulators, TIMPs and MMPs were also different between the 2D and 3D cultured HconF cells (Fig. 6). That is, the gene expression of both TIMP 1 and 4 or MMP2 were significantly downregulated or upregulated, respectively, and that for MMP14 was relatively downregulated by TGF-β2 independent of PGF2α or OMD in the 2D HconF cells. However, in contrast, the values for TIMPs or MMPs were relatively or significantly downregulated by TGF-β2, or PGF2α or OMD, respectively, in the 3D HconF spheroids.

Effects of PGF2α or OMD on the mRNA expression of TIMPs and MMPs of 2D and 3D cultured HconF cells in the absence and presence of TGF-β2. 2D and 3D HconF cells were untreated or treated with a 5 ng/mL solution of TGF-β2 in the absence or presence of 500 nM PGF2α or OMD. At Day 6, each sample was subjected to qPCR analysis to estimate the expression of mRNA in TIMP1–4
Therefore, the observed diversity in the expression of ECM protein molecules as well as ECM modulators between 2D and 3D HconF cells again suggest that PGF2α or OMD may affect epithelial or subepithelial fibrogenesis in different manners, as suggested earlier.
Discussion
Previous studies have reported on positive expressions of FP, a PGF2α receptor, within not only the ciliary body, but also in ocular surface tissues, including scleral and subconjunctival tissues, suggesting that FP receptor agonists might affect ocular surface tissues.35,36 Alternatively, in terms of PGE2 linked receptors, it is well known that 4 types of G protein-coupled receptors, that is, prostaglandin receptors EP 1 through 4 are present. 46 Functionally, EP1 functions to enhance the intracellular levels of free calcium, 47 and EP2 and EP4 or EP3 stimulate or inhibit cAMP production among various systemic organs. 48 In a previous study, quantitative real-time RT-PCR revealed that the EP2 receptor levels were higher than those of other EP receptors within conjunctival fibroblasts, and in fact, the EP2 agonist, butaprost inhibited the proliferation of conjunctival fibroblasts without toxicity against Tenon's tissue. 49
Therefore, taken together, it was suggested that the underlying mechanism involved the activation of the EP2 receptor, leading to an enhancement in cAMP cascade signaling. Since it is also well known that stimulation of the FP receptor increases [Ca2+]i by the activation of inositol triphosphate and diacylglycerol synthesis through coupling to the Gq protein, 50 we rationally speculate that FP receptor agonists and the EP2 agonist, OMD could also induce diverse effects on ocular surface tissues including conjunctival fibrogenetic changes. In fact, in terms of the cosmetically annoying local side effects referred to as “upper eyelid sulcus (DUES)” presumably caused by FP receptor agonists that induce orbital fat atrophy,44,45,51,52 our group recently reported that the EP2 agonist, OMD did not induce such adipocyte atrophy in 2D and 3D cultures of 3T3-L1 mouse preadipocytes 34 as well as in human orbital fibroblasts. 33
In this study, we examined some of the currently unidentified effects on conjunctival fibrogenetic changes by FP receptor agonists and/or the EP2 agonist, OMD, and the findings indicate that both agonists appear to modulate both planar and subepithelial fibrogenetic changes of HconF cells but in different manners. To support current observation, a recent crossover study indicated that the switch from PGF2ags to OMD significantly reduced the eyelid skin coloring and eyelash elongation in addition to DUES despite the lowering of IOP remained intact in glaucoma patients. 53
Since cellular metabolism is known to play an important role in the regulation of fibrotic processes, this study examined the effects of the prostanoid FP and EP2 receptor agonists, PGF2α and OMD, respectively, on metabolic properties in the absence or presence of TGF-β2 stimulation, a major fibrotic signal in HconF cells. TGF-β2 tended to shift the metabolism toward energetic states in HconF cells as we previously reported, 54 and PGF2α or OMD increased ATP-linked respiration regardless of whether TGF-β2 was absent or present, whereas these treatments resulted in decreased mitochondrial maximal respiratory activity and canceled TGF-β2-induced enhancement of glycolytic capacity.
Although the underlying mechanisms responsible for these effects may be complex, one possibility is that agonists that can increase intracellular calcium levels through Gq proteins, such as PGF2α, have the potential to enhance ATP-linked respiration through the calcium-dependent activation of the TCA cycle. 55 In addition, decreased maximal respiration by PGF2α can be explained by the increased production of reactive oxygen species (ROS) with mitochondrial respiration in response to metabolic demand, potentially leading to impairments in the electron transport chain system.
Indeed, it has been reported that a mitochondrial calcium overload can induce disintegration of the mitochondrial respiratory supercomplex. 56 Concerning OMD, it is also possible that calcium-independent ROS production may result in these metabolic phenotypes. 57 Nevertheless, these unique changes in cellular metabolism appear to be associated with the cellular properties of 3D spheroids of HconF cells that are altered by PGF2α or OMD in the presence or absence of TGF-β2.
ECM proteins serve as essential structural support in addition to having several cellular functions that are critically regulated by several factors, including TIMPs, MMP, PGs, and others.58,59 In this study, TEER measurements of the 2D cultured HconF cells and stiffness measurements of the 3D HconF spheroids suggested that both epithelial and subepithelial epithelial-mesenchymal transformation (EMT) changes were significantly stimulated by FP receptor agonists and the EP2 agonist, OMD. To support our current observations, previous studies have reported that PGF2α facilitates pulmonary fibrosis through PGF receptors (FP) in a murine bleomycin-induced pulmonary fibrosis model 60 and the selective EP2 agonist, butaprost, inhibits myofibroblast differentiation in the lungs, thereby being involved in the pathogenesis of fibrotic changes of the lungs and other organs.61–64
It should be noted that we observed similar effects on ECM proteins and their modulators, TIMPs, MMPs between FP receptor agonists, and the EP2 agonist, OMD of 2D or 3D cultured HconF cells. Those effects, however, were exclusively different between both culture systems, in that both drugs induced insignificant effects or marked enhancement effects toward ECM proteins of 2D or 3D HconF cells, respectively.
Therefore, the collective findings reported in this study suggest that FP receptor agonists or the EP2 agonist induced effects toward epithelial and subepithelial EMT changes may be caused by different mechanisms, including the metabolism of ECM protein molecules. Taking the relationship between conjunctiva fibrosis and failure of glaucoma filtration surgery into account, we speculate that the diverse effects on the metabolism of ECM proteins between FP receptor agonists and an EP2 agonist on fibrogenetic changes provide important information regarding the choice of appropriate glaucoma medications to be used, especially before glaucoma filter surgery.
Although only a few related studies regarding this issue have appeared so far, we have also observed several diverse effects of FP receptor agonists and an EP2 agonist on TGF-β2 related or unrelated fibrogenetic changes in conjunctiva. However, the precise underlying mechanisms responsible for causing these diverse effects toward ECM protein metabolisms by both agonists as observed in the biophysical analyses of the aforementioned 2D and 3D HconF cells remain to be elucidated. Therefore, to better understand the results related to the role of both agonists toward fibrogenetic changes of 2D and 3D cultured HconF cells, further investigations, including an RNA-Sequencing experiment, will be needed to elucidate possible mechanisms linked with several other complex signaling networks to confirm our present observations, together with several observational clinical studies of patients for which both agonists were used.
Footnotes
Authors' Contributions
Y.T. performed the experiments, analyzed the data, and wrote the article. T.S., M.W., and M.F. performed the experiments and analyzed the data. H.O. designed the experiments, analyzed the data, and wrote the article.
Data Availability Statement
The data sets used and/or analyzed during this study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.