
Abstract
Purpose:
This study clarifies the beneficial effects of MG132, a proteasomal inhibitor, on retinal vascular injury mediated by diabetes-induced oxidative stress through nuclear factor erythroid 2-related factor 2 (Nrf2).
Methods:
Diabetic rats and control animals were randomly assigned to receive MG132 or vehicle for 24 weeks, and human retinal endothelial cells (HRECs) were incubated with normal or high glucose with or without MG132. 26S proteasome activity in the rat retinas or cultured HRECs was measured using Suc-LLVY-7-amido-4-methylcoumarin. NADPH-quinone oxidoreduc-tase (NQO1), heme oxygenase (HO)-1, kelch-like ECH-associated protein 1 (Keap1) and Nrf2 were examined by Western blotting and real-time reverse transcription polymerase chain reaction. Cell apoptosis is measured through flow cytometry assay, mitochondrial reactive oxygen species (ROS) production, and retinal vascular leakage were assayed using CM-H2DCFDA fluorescent probes and Evans blue, respectively.
Results:
MG132 significantly inhibited the activation of 26S proteasome induced by diabetes or elevated glucose, and subsequently increased the expression of Nrf2, NQO1, and HO-1, and further reduced ROS accumulation. These changes were associated with a decrease of diabetes-induced retinal vascular leakage and retinal capillary cell apoptosis.
Conclusions:
MG132 decreases diabetes-induced 26S proteasome activation and exerts protective effects against retinal microvascular dysfunction in diabetic rats in association with the alleviation of retinal oxidative stress mediated by Nrf2.
Introduction
Diabetic retinopathy (DR), the leading cause of blindness in working-age population, is one of the most severe microvascular complications of diabetes mellitus (DM). 1 Retinal vasculature alterations, including increased vascular permeability, endothelial apoptosis, pericyte loss, and cellular capillary development are early events in DR and contribute to breakdown of the blood–retinal barrier (BRB) and subsequent diabetic macular edema and pathologic angiogenesis regarded as the leading cause of diabetes-associated vision loss. 2 Therefore, the identification of pharmacological targets for the prevention of damage to retinal vasculature from diabetes may provide a potential therapeutic strategy for diabetes-induced vision loss.
Overwhelming evidence implicates that oxidative stress plays a pivotal role in the etiology of diabetic complications.3,4 A unifying hypothesis of the pathogenesis of DR has been proposed, in which production of mitochondrial reactive oxygen species (ROS) in response to chronic hyperglycemia, and these ROS in turn activate stress-sensitive intracellular signaling pathways and might be the key initiator for all of the pathogenic pathways and lead to diabetes complications, such as retinopathy. 4 Thus, it has been hypothesized that ROS might be an important target and antioxidants will be beneficial in treating diabetic complications by preventing the accumulation of ROS. While results from clinical trials thus far using general antioxidants have been disappointing. 5 Therefore, new approaches for amelioration and treatment of DR, especially in the earlier retinal vascular injury, are thus urgently needed.
Researchers have shown that MG132, a cell-permeable proteasome inhibitor, has therapeutic effects on diabetes-associated renal,6,7 cardiac, 8 aortic, and retinal 9 diseases through inhibiting the increase of ROS generation. Despite considerable patent in MG132 performed in regulating intracellular redox status, the mechanism by which it acts is unclear. Prior studies have suggested that MG132 can upregulate nuclear factor erythroid 2-related factor 2 (Nrf2)-mediated antioxidative function by inhibiting its proteasomal degradation and stimulate Nrf2 translocation into the nucleus in vascular endothelial cells. 10 Nrf2 is considered to be one of the most important cellular defense mechanisms to cope with oxidative stress11,12_ENREF_12. In response to oxidative stress, Nrf2 translocated to the nucleus and binds to antioxidant response elements (AREs), resulting in the transcriptional activation of phase II detoxifying and multiple antioxidants, including NADPH-quinone oxidoreduc-tase (NQO1), heme oxygenase (HO)-1, catalase, superoxide dismutase, glutathione peroxidase, and glutamate–cysteineligase. 13
Despite evidence is accumulating regarding a potential protective role for Nrf2 in the retina and MG132 has been found to be protective in the context of diabetic disease,6–8,10,14,15 direct preventing effects of MG132 on retinal vascular injury in DR is still lacking.
Thus, to investigate the potential effect and molecular mechanisms of MG132 on retinal vascular injury in vivo and in vitro, the rat DR model and the human retinal capillary endothelial cells' high D-glucose (HG) model were constructed in the present study. We speculate that the upregulation of Nrf2 was associated with the protective effect of MG132 on DR, and our findings indicate that MG132 possesses a great potential for treatment of the disease.
Methods
Ethics statement
This study was carried out according to the recommendations of the Guide for the Care and Use of Laboratory Animals (National Institutes of Health, Bethesda, MD). The protocol was approved by the Committee on the Ethics of Animal Experiments of Wenzhou Medical University, Zhejiang, China. The experiments of this study were carried out at the Key Laboratory of Lishui Municipal Central Hospital and performed in accordance with the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmic and Vision Research. All surgical procedures were performed after administering sodium pentobarbital anesthesia, and all efforts were made to minimize the suffering of the rats.
Materials
All chemicals were of reagent grade and were purchased from Sigma Chemicals (St. Louis, MO), unless otherwise stated.
Animals
We used male Sprague–Dawley rats (8 weeks old) weighing 180–200 g (Shanghai Laboratory Animal Center, Chinese Academy of Sciences). Animals were kept under a 12-h light/12-h dark cycle; the room temperature was maintained at 23°C–25°C, and the humidity was maintained at 55%–60%. Food and water were available ad libitum. The rats were acclimatized for 1 week and subsequently divided into two groups. The first group of rats was administered a 60 mg/kg dose of STZ intraperitoneally and the second group of rats was administered citrate buffer. The first group of rats became diabetic as their blood glucose levels exceeded 16.7 mmol/L, at 48 h after STZ administration. The control animals (Con; n = 18) were age matched before being administered an equal volume of sodium citrate. Two weeks after inducing diabetes, the rats were divided randomly into two subgroups: diabetic rats (DM; n = 18) and diabetic rats treated with 0.5 mg/kg/day MG132 (DM+MG132; n = 18). MG132 (Calbiochem) was dissolved in dimethylsulfoxide and diluted with saline, and administered by intraperitoneal injections for 24 weeks.
Meanwhile, the Con and DM groups received intraperitoneal injections of the same volume of saline containing dimethylsulfoxide every day. Unless otherwise noted, rats were euthanized 24 weeks after the spontaneous onset of hyperglycemia, when their age was typically 26 weeks.
26S proteasome activity assay
The 26S proteasome activity was measured as described previously by measuring ATP-dependent degradation of proteasome fluorescence substrate.16,17 Briefly, cells were washed with phosphate-buffered saline (PBS) and then with buffer I (50 mmol/L Tris, pH 7.4, 2 mmol/L DTT, 5 mmol/L MgCl2, 2 mmol/L ATP). The cells were then made into pellets by centrifugation. Homogenization buffer (50 mmol/L Tris, pH 7.4, 1 mmol/L DTT, 5 mmol/L MgCl2, 2 mmol/L ATP, 250 mmol/L sucrose) was added, and cells were subjected to a vortex for 1 min. Cell debris was removed by centrifugation at 1,000 g for 5 min followed by 10,000 g for 20 min. Protein concentration was determined by the bicinchoninic acid protocol (Pierce, Rockford, Ill). Protein (100 μg) from each sample was diluted with buffer I to a final volume of 1,000 μL.
The fluorogenic proteasome substrate Suc-LLVY-7-amido-4-methylcoumarin (chymotrypsin-like, Sigma, St. Louis, Mo) was added at a final concentration of 80 μmol/L in 1% DMSO. Cleavage activity was monitored continuously by detection of free 7-amido-4-methylcoumarin with a fluorescence plate reader (Gemini, Molecular Devices, Sunnyvale, CA) at 380/460 nm at 37°C.
Permeability assay
Retinal vascular permeability was determined by the Evans Blue albumin leakage assay as described previously. 18 BRB breakdown was calculated and visualized under a confocal microscope (Eclipse E800; Nikon, Tokyo, Japan).
Cell culture and treatment
Human retinal endothelial cells (HRECs) and attachment factor were purchased from the Tongpai (Shanghai) Biotechnology and cultured according to our previous study. 19 Briefly, HRECs were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (Science Cell, San Diego, CA), 100 U/mL penicillin (Invitrogen), 100 mg/mL streptomycin (Invitrogen) at 37°C in a humidified atmosphere containing 5% CO2 and 95% humidified air. Cultured HRECs at passages 3 or 4 were used in the experiments that follow. Confluent HRECs were maintained in DMEM and supplemented with 0.4% BSA. The cells were incubated for 96 h in normal D-glucose (5.5 mM; NG), NG plus 0.5 μM MG132 (NG+ MG132), HG (30 mM), or HG in the presence of 0.5 μM MG132 (HG+ MG132).
Cell transfection
The ORF of Nrf2 cDNA was inserted into a pcDNA3.1 vector to obtain the Nrf2 expression vector. The vector was transfected into cells using Lipofectamine according to the manufacturer's instructions.
Small interfering RNA (siRNA) for Nrf2 (20 nmol/L) were from Genesil Biotechnology (Wuhan, China). Transfection of siRNA Predesigned siRNA against human Nrf2 and the negative-control siRNA were chemically synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China) using the following sequences:
#Con, 5′-GGAUUAUUAUGACUGUUAAAU-3′ (sense) and 5′-UUAACAGUCAUAAUAAUCCUU-3′ (antisense);
#1, 5′-GAGUUACAGUGUCUUAAUAUU-3′ (sense) and
5′-UAUUAAGACACUGUAACUCAG-3′ (antisense); and
#2, 5′-CAGUCUUCAUUGCUACUAAUC-3′ (sense) and
5′-UUAGUAGCAAUGAAGACUGGG-3′ (antisense).
The cells were transfected with 10 nM siRNA duplexes using Lipofectamine 2,000 reagent (Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions.
After 24 h incubation, fresh medium was added, and the cells were cultured for another 48 h. Then, we used western blot analysis to detect the efficiency of Nrf2 overexpression and Nrf2 silencing.
Detection of ROS
The cellular ROS generation was measured using 2′,7′-dichlorofluorescein diacetate (DCFH-DA) fluorescence assay with the ROS Assay Kit (Beyotime Biotechnology) according to the manufacturer's recommendation as described in our previous study.19,20
RNA extraction and real-time reverse transcription polymerase chain reaction
Total RNA was extracted from rat retinal tissue, harvested as described above, or from HRECs using TRIzol reagent (Invitrogen Life Technologies, Gaithersburg, MD) and stored at −80°C. The DyNAmo Flash SYBR Green qPCR Kit (Finnzymes Oy, Espoo, Finland) was used according to the manufacturer's instructions. The reaction conditions were 40 cycles of two-stage polymerase chain reaction (PCR) consisting of denaturation at 95°C for 30 s annealing at 60°C for 30 s after an initial denaturation step at 95°C for 5 s. The sequences of the primers (sense/antisense) used were: Nrf2, 5′-GAGACGGCCATGACTGAT-3′/5′-GTGAGGGGATCGATGAGTAA-3′;
NQO1, 5′-AGACAAACATTCAAGCCGCT-3′/5′-
CCATCTCTTGTTTGCTGCAG-3′;
HO-1, 5′-TGCTCGCATGAACACTCTG-3/5′TCCTCTGTCAGCAGTGCC-3′
β-actin, 5′-CGACAACGGCTCCGGCATGT-3′/5′-GGGGCCACACGCAGCTCA TT-3′). The specificity of the amplification product was determined by performing a melting curve analysis.
Standard curves were generated for each gene by using serial dilutions of known quantities of the corresponding cDNA gene template. Relative levels of target gene mRNA expression were calculated using the 2−ΔΔCT method. Amplification of the target gene cDNA was normalized to β-actin expression.
Western blot analysis
Total lysates from retinas or cells were extracted with RIPA buffer and western blot analysis of proteins were performed as described before. 18 Rabbit anti-kelch-like ECH-associated protein 1 (Keap1) antibody (SC365626, 1:500; Santa Cruz), rabbit anti-Nrf2 antibody (affinity, AF0639, 1:500), rabbit anti-NQO1 antibody (affinity, DF6437, 1:1,000), and mouse anti-HO-1 antibody (affinity, AF5393, 1:1,000) were used to detect each protein and blotted with a horseradish peroxidase-conjugated secondary antibody. ECL was used to detect the immunoreactive bands. Detection of GAPDH expression with anti-GAPDH antibody (ab9485, 1:1,000; abcam) was used as an internal control to confirm equivalent total protein loading. All measures are expressed relative to the signal intensities measured in the control lanes, which were assigned a value of 1.0. Western blots were repeated 3 to 5 times and qualitatively similar results were obtained.
Detection of apoptosis by flow cytometry
After 24 h, cells were washed in Annexin V binding buffer and were incubated in 500 μL Annexin V staining solution (1:50 dilution of Annexin V–phycoerythrin in Annexin V binding buffer) for 10 min in the dark at 37°C, 5% CO2. The cells were then washed with ice-cold PBS twice and resuspended in the binding buffer, then 10 μL of Propidium Iodide (PI) was added for 10 min at 4°C. Flow cytometry was used to collect 5,000 cells, which were sorted based on whether they were stained with PI (apoptotic or dying cells) or were unstained with either fluorochrome (live cells). All data were collected, stored, and analyzed by Multigraph software (Coulter, Miami, FL).
Statistical analyses
GraphPad Prism 8.0.1 software system (GraphPad, San Diego, CA) and the statistical software program SPSS version 22.0 (SPSS, Chicago, IL) were used for data analyses. Each experiment was repeated three times. The Shapiro–Wilk test was used to test normality. Differences among groups were analyzed using one-way ANOVA followed by Tamhane's test and the least significant difference method. The Kruskal–Wallis test was used when data did not conform to a normal distribution. Data are shown as standard mean ± standard deviation, P < 0.05 denoted significance in all cases.
Results
Animal data
Within 24 weeks of the onset of diabetes, nonfasting blood glucose was markedly higher in the diabetic rats than in the control group rats (21.8 ± 3.5 mmol/L vs. 5.7 ± 0.5 mmol/L; P < 0.01), and administration of MG132 showed no effect on hyperglycemia in diabetic rats (20.9 ± 3.1 mmol/L vs. 21.8 ± 3.5 mmol/L; P > 0.05). We also found that there was a significant decrease in the body weight of diabetic rats compared with the rats in the control group (314.6 ± 27.1 g vs. 552.7 ± 29.5 g; P < 0.01), and the MG132-treated group showed no effect on body weight (321.4 ± 26.2 g) when compared with the diabetic rats (P > 0.05).
MG132 alleviation of retinal vascular permeability in diabetic rats
Evans Blue was applied to retinal flat mounts to evaluate the effect of MG132 on retinal blood vessel leakage. In control retinas, Evans Blue fluorescence was located within blood vessels, whereas in STZ-treated rats, focal leakage of the dye from capillaries and larger vessels was detected, and treatment with MG132 prevented this effect (Fig. 1A–C). The Evans Blue levels in the retina were measured to assess BRB permeability. Evans Blue levels were elevated in the retinas of STZ-treated diabetic rats (2.88 ± 0.33 μg Evans Blue/g wet wt retina) as compared with control rats (0.96 ± 0.12 μg Evans Blue/g wet wt retina). In agreement with the reduced vascular leakage revealed by whole-mount imaging, this elevation was significantly reduced in STZ-treated rats administered MG132 (1.14 ± 0.31 μg Evans Blue/g wet wt retina) (Fig. 1D). The eyes in theMG132 treatment group showed significantly lower vascular permeability in the retina, compared with that in the STZ-treated eyes.
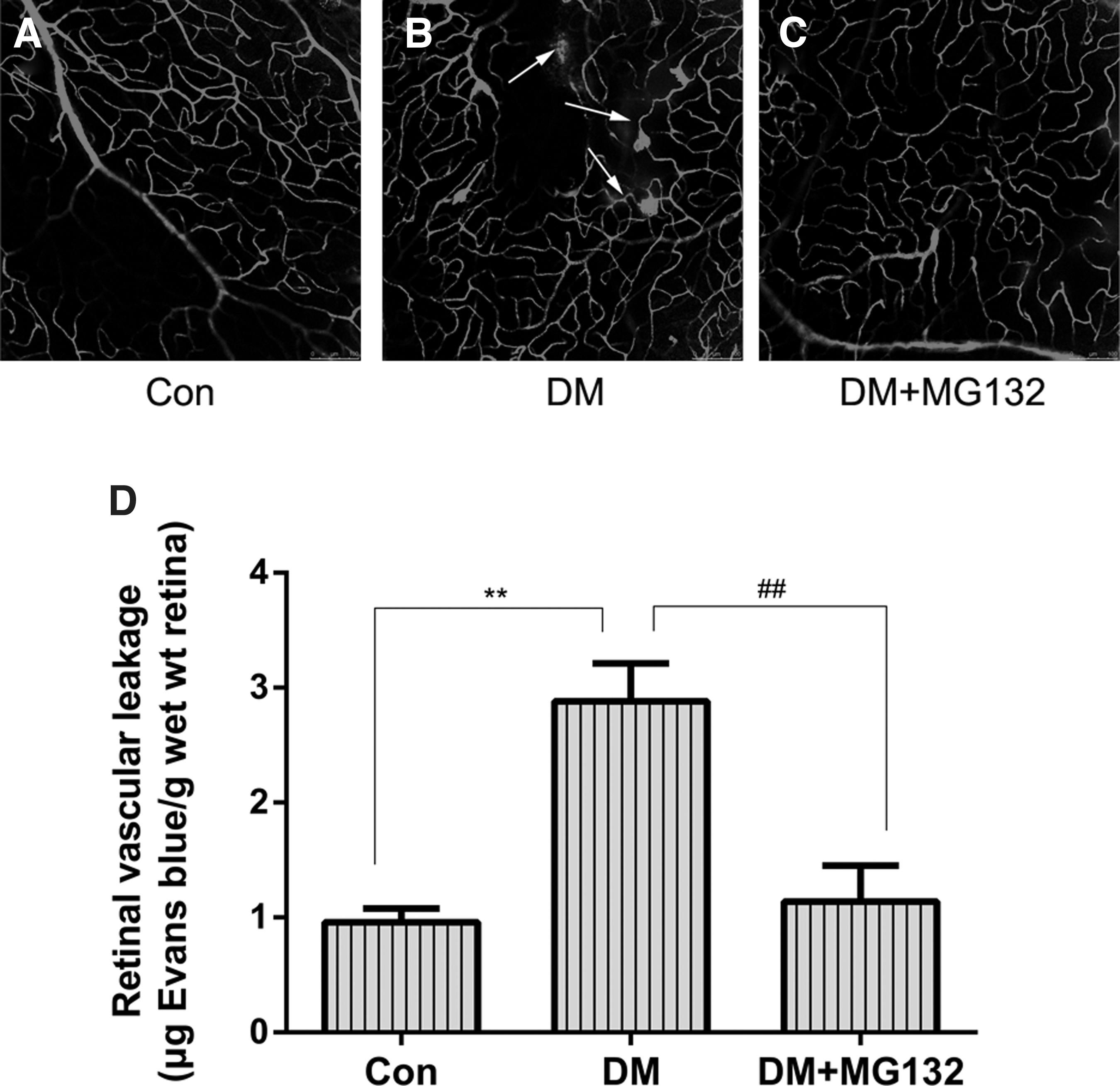
Visualization of retinal blood vessel leakage with Evans Blue.
MG132 alleviation of ROS production in diabetic rats
Spectrofluorometric analysis was used to quantify intracellular ROS levels. The 2′,7′-dichlorofluorescein (DCF) fluorescence intensity was increased by about 100% in the retinas of diabetic rats compared with control rats. MG132 treatment significantly reduced the DCF fluorescence intensity by about 33% as compared with that in STZ-treated rats (Fig. 2).
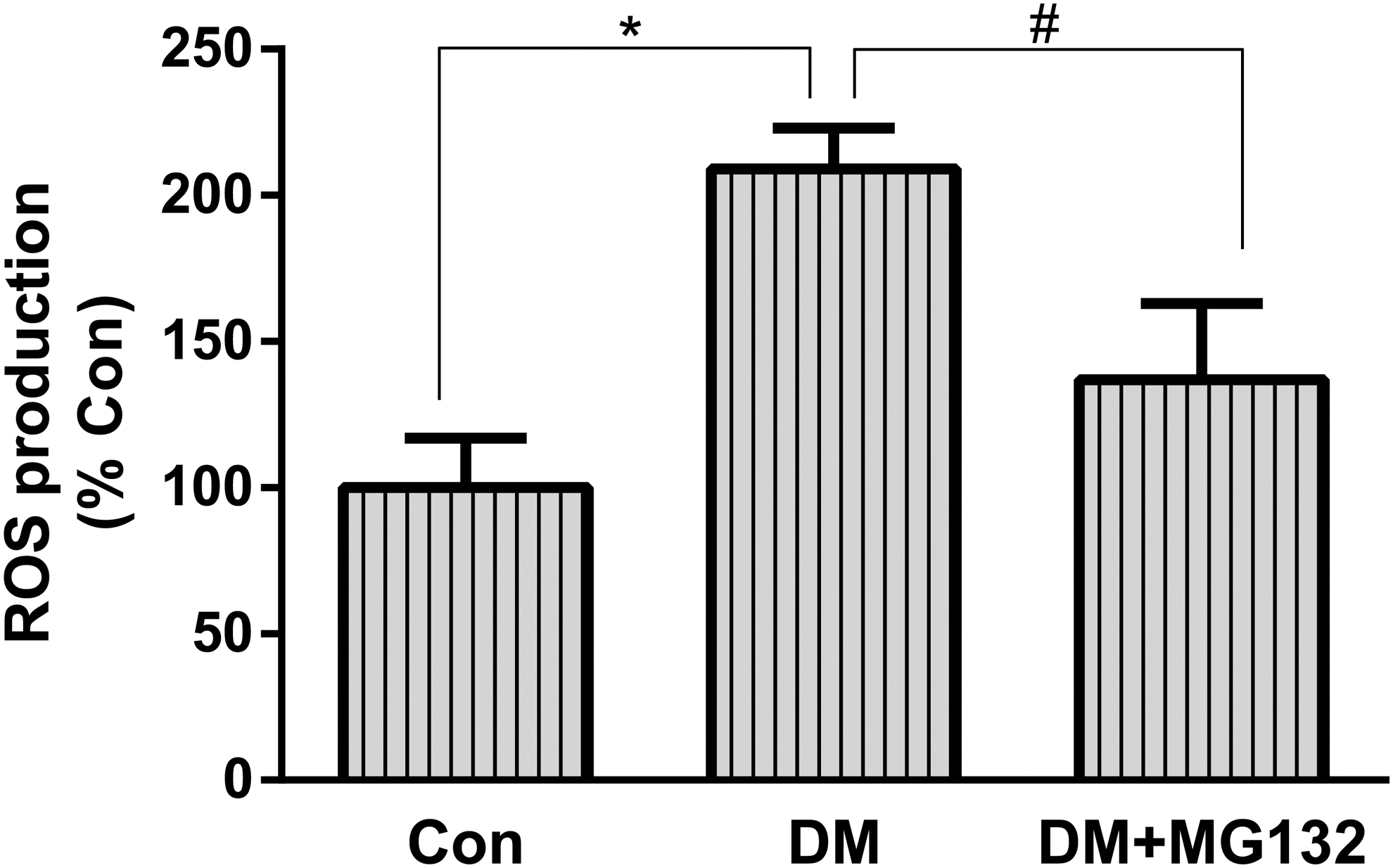
Effect of MG132 on ROS production in rat retinas. Freshly prepared rat retinal homogenates were incubated with DCFH-DA, and DCF fluorescence intensity was quantitated. Data are mean ± SD from six rats per group, and the experiments were repeated independently at least three times with similar results. *P < 0.05 versus control; #P < 0.05 versus diabetes. DCF, 2′,7′-dichlorofluorescein; DCFH-DA, 2′,7′-dichlorofluorescein diacetate; ROS, reactive oxygen species.
Effect of MG132 on retinal 26S proteasome activity
The 26S proteasome, as a key component in the ubiquitin–proteasome system (UPS), functions to recognize, unfold, and ultimately destroy proteins. To confirm that enhancement of functionality was due to activity elevation, we chose to measure 26S proteasome activity (ATP-dependent chymotrypsin-like activity) in retinal tissues. A representative figure showing that 26S proteasome activity was increased by about threefold in the retinas of diabetic rats compared with control rats, this elevation was significantly reduced in STZ-treated rats administered MG132 (Fig. 3).
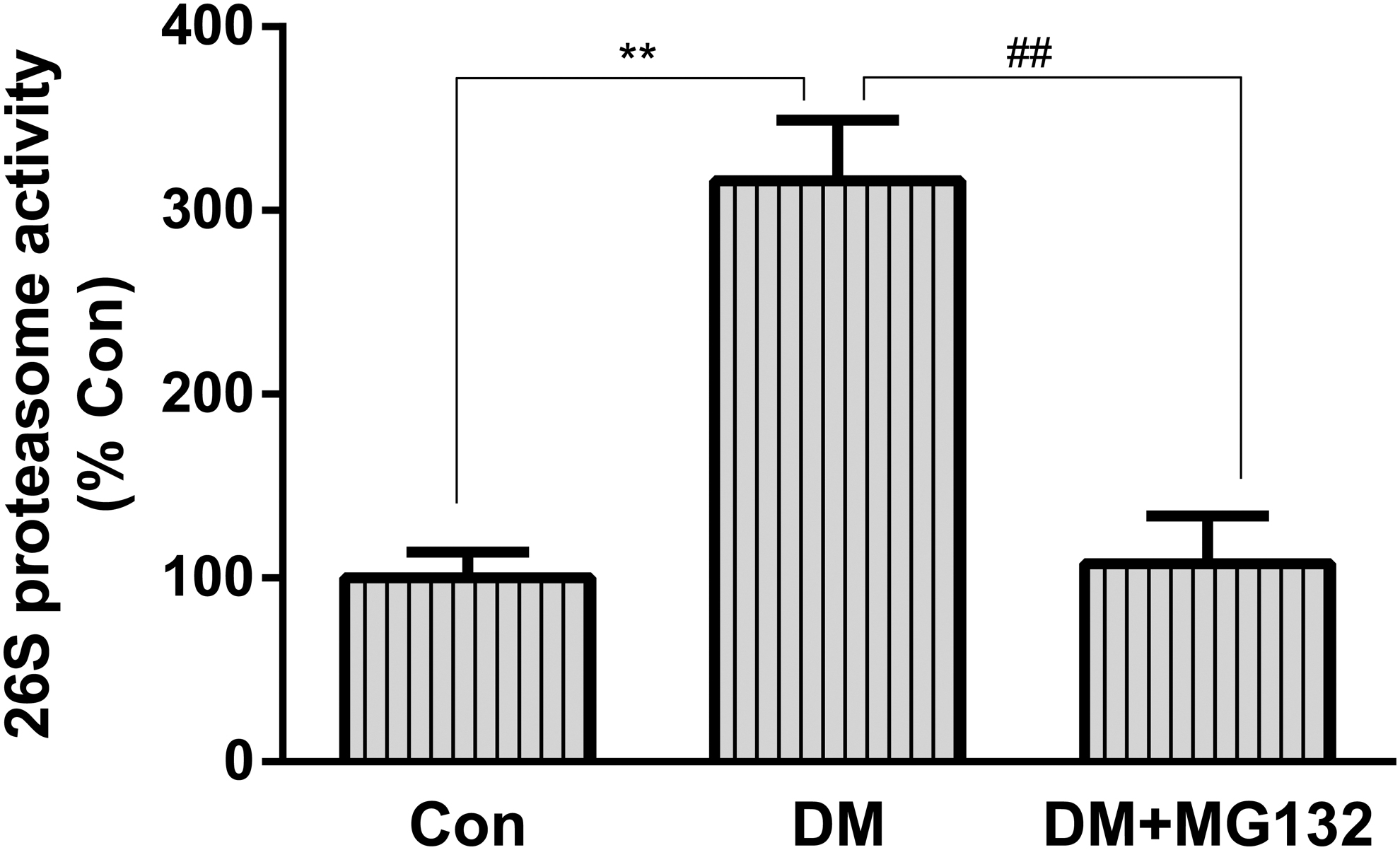
Effects of MG132 on 26S proteasome activity in rat retinas. 26S proteasome activity assay was performed with fluorescent proteasome substrates Suc-LLVY-7-amido-4-methylcoumarin in retinal tissue homogenates. Fluorescence was measured at emission 460 nm/excitation 380 nm. Data are mean ± SD from six rats per group, and the experiments were repeated independently at least three times with similar results. **P < 0.01 versus control; ##P < 0.01 versus diabetes.
MG132 unregulated Nrf2 and downstream antioxidant protein expression while reducing Keap1 protein level in diabetic rats
Western blot showed that, compared with the control rats, the diabetic rat group showed significantly increased expression of Keap1 protein in retinal tissues, plus decreased both of Nrf2 and Nrf2-target antioxidant proteins, including NQO1 and HO-1, whereas MG132 significantly inhibited these changes. Furthermore, we showed that MG132 did not increase Nrf2mRNA levels in diabetic rats by RT-PCR. (Fig. 4).
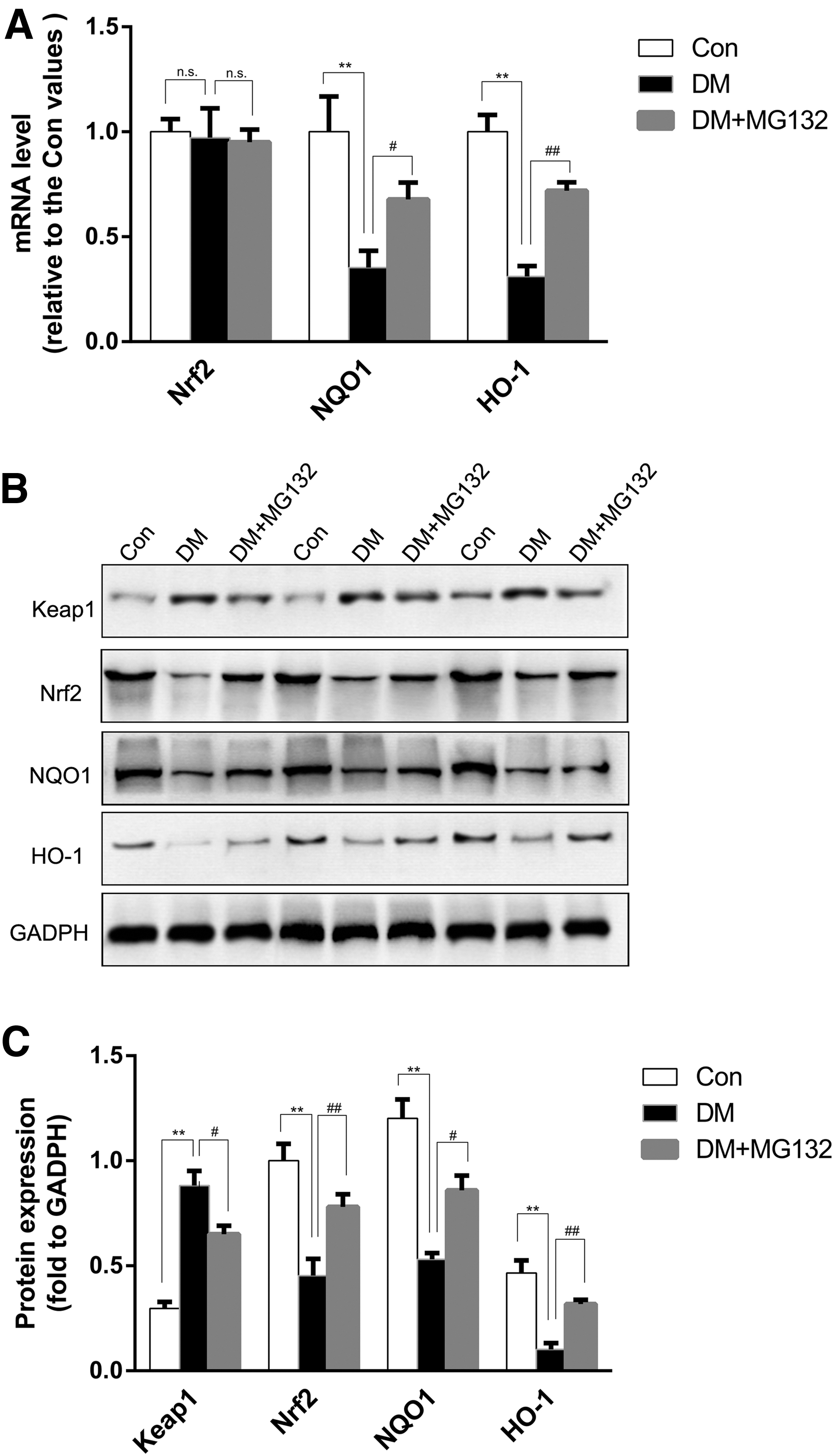
Effects of MG132 on Keap1, Nrf2, NQO1, and HO-1 expression in rat retinas.
MG132 reverts HG increased 26S proteasome activity in retinal capillary cells
In addition, we examined the effect of MG132 on 26S proteasome activity in HRECs. Total 26S proteasome activity in HRECs was assayed with specific fluorescent proteasome substrates under our experimental conditions. As depicted in Fig. 5, exposure of HRECs to HG but not NG significantly increased 26S proteasome activity by threefold; this elevation was significantly reduced in cells exposed to high glucose in the presence of MG132.
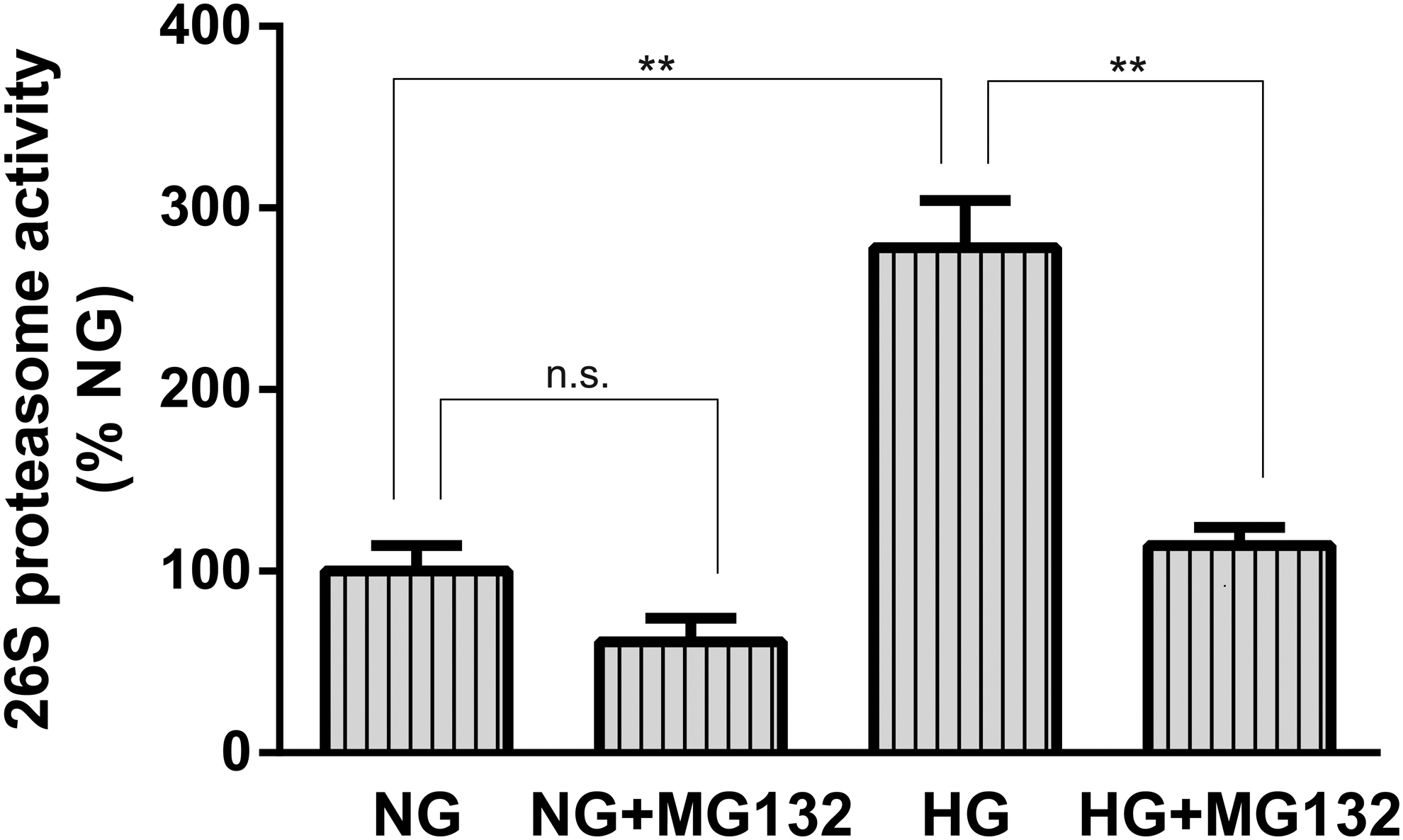
Effects of MG132 on 26S proteasome activity in HRECs. 26S proteasome activity assay was performed with fluorescent proteasome substrates Suc-LLVY-7-amido-4-methylcoumarin in confluent HRECs, which were cultured in NG or HG with or without MG132 preincubation. Fluorescence was measured at emission 460 nm/excitation 380 nm. Data are mean ± SD from at least three independent experiments with similar results. **P < 0.01 versus HG. HG, high D-glucose; HRECs, human retinal endothelial cells.
MG132 unregulated Nrf2 and downstream antioxidant protein expression in retinal capillary cells under high glucose
We examined the expression levels of Keap1, Nrf2, and downstream antioxidant protein, including NQO1 and HO-1 under our experimental conditions. As shown in Fig. 6A–C, compared with normal glucose, high glucose induced a significant decrease of Nrf2 protein expression, which remained increased in cells exposed to high glucose in the presence of MG132; and similar changes occurred for the protein levels of NQO1 and HO-1 (Fig. 6A, B). Furthermore, we showed that MG132 did not increase Nrf2 mRNA levels in high glucose conditions by RT-PCR.
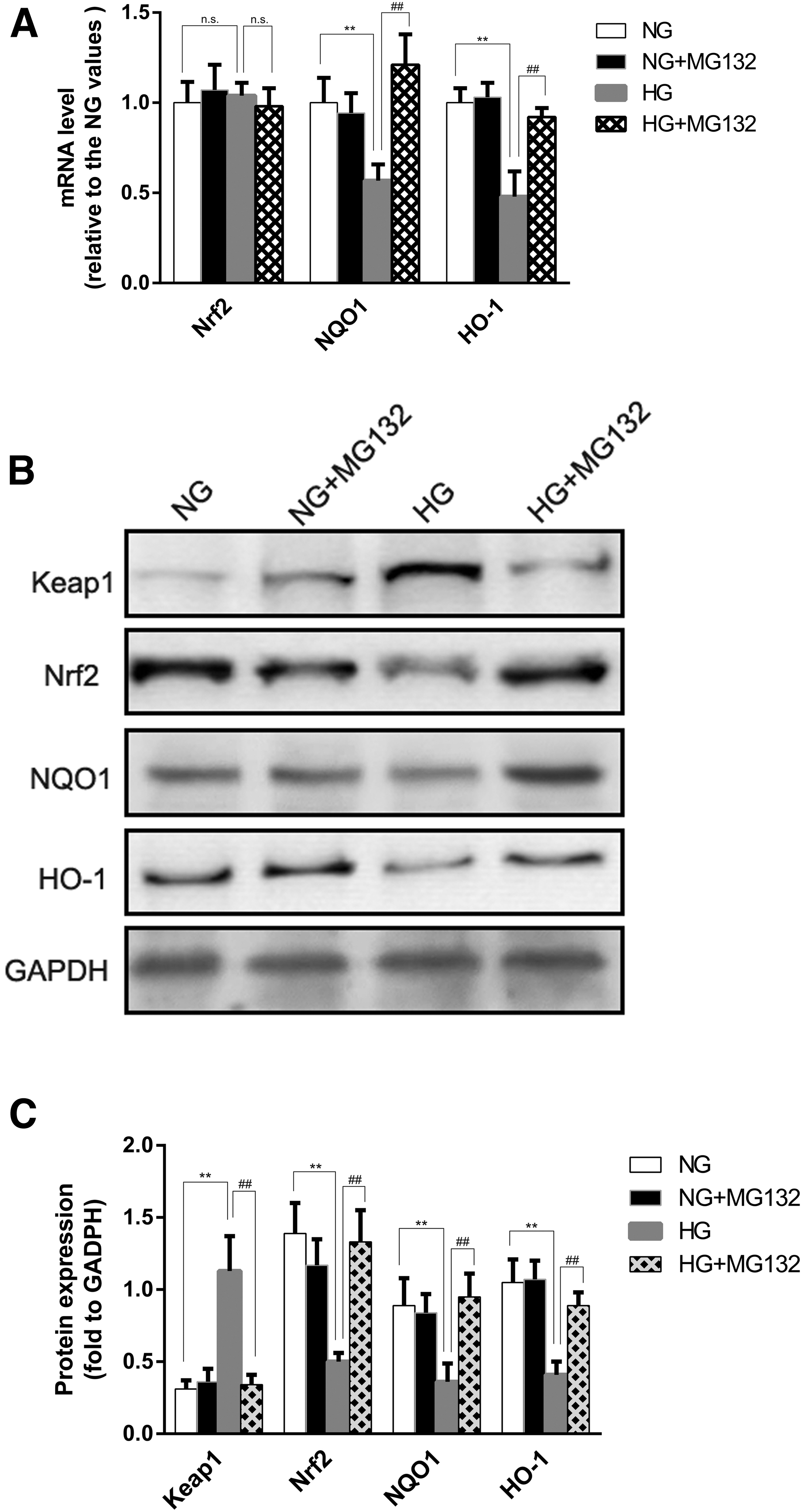
Effects of MG132 on Keap1, Nrf2, NQO1, and HO-1 expression in HRECs.
MG132 reverts hyperglycemia-induced retinal capillary cell ROS production through targeting Nrf2
The fluorescent probe CM-H2DCFDAwas used to investigate effects of MG132 on ROS production in retinal capillary cells under our experimental conditions. The results of this study indicated that HG significantly increased the ROS production compared with the normal control group (P < 0.05). The addition of MG132 significantly decreased the ROS production compared with HG-treated cells (P < 0.05), thereby MG132 could protect against HG-induced oxidative stress in retinal capillary cells (Fig. 7E, F).

The role of Nrf2 in MG132 treatment of HG-induced ROS production in HRECs.
To investigate the mechanism by which MG132 reduces ROS production, we used siRNA to modulate Nrf2. We first performed western blot analysis to confirm that transfecting NRF2 siRNA efficiently decreased Nrf2 protein levels in retinal capillary cells, and Nrf2 siRNA2 provided the highest silencing efficiency (Fig. 7A, B). We also transfected HRECs with the PcDNA3.1(+)-Nrf2 plasmid to overexpress Nrf2. Then, we examined the expression levels of Nrf2; as expected, the results indicated that this method successfully induced Nrf2 overexpression (Fig. 7C, D).
In addition, we also evaluated whether Nrf2 regulates MG132-mediated hyperglycemia-induced ROS production, as shown in Fig. 7E and F. The addition of Nrf2 siRNA significantly enhanced ROS production compared with NG-treated cells (P < 0.01), conversely, Nrf2 overexpression significantly reduced HG-induced ROS production (P < 0.01). Moreover, Nrf2 siRNA also dramatically increased ROS production compared with HG combined with MG132-treated cells (P < 0.05).
These data indicate that MG132 modulates hyperglycemia-induced retinal capillary cell ROS production through targeting Nrf2.
MG132 reverts hyperglycemia-induced retinal capillary cells apoptosis through targeting Nrf2
Flow cytometry (Fig. 8A, B) was used to investigate effects of MG132 on apoptosis in retinal capillary cells under our experimental conditions. The results showed a significant increase in the number of apoptotic retinal capillary cells in cultures exposed to HG (37.70% ± 8.60%) compared with cultures exposed to NG (3.34% ± 1.02%, P < 0.01). The increase in the number of apoptotic cells in the hyperglycemic group was partly reversed in the presence of MG132 (12.92% ± 3.51%, P < 0.01 compared with HG).
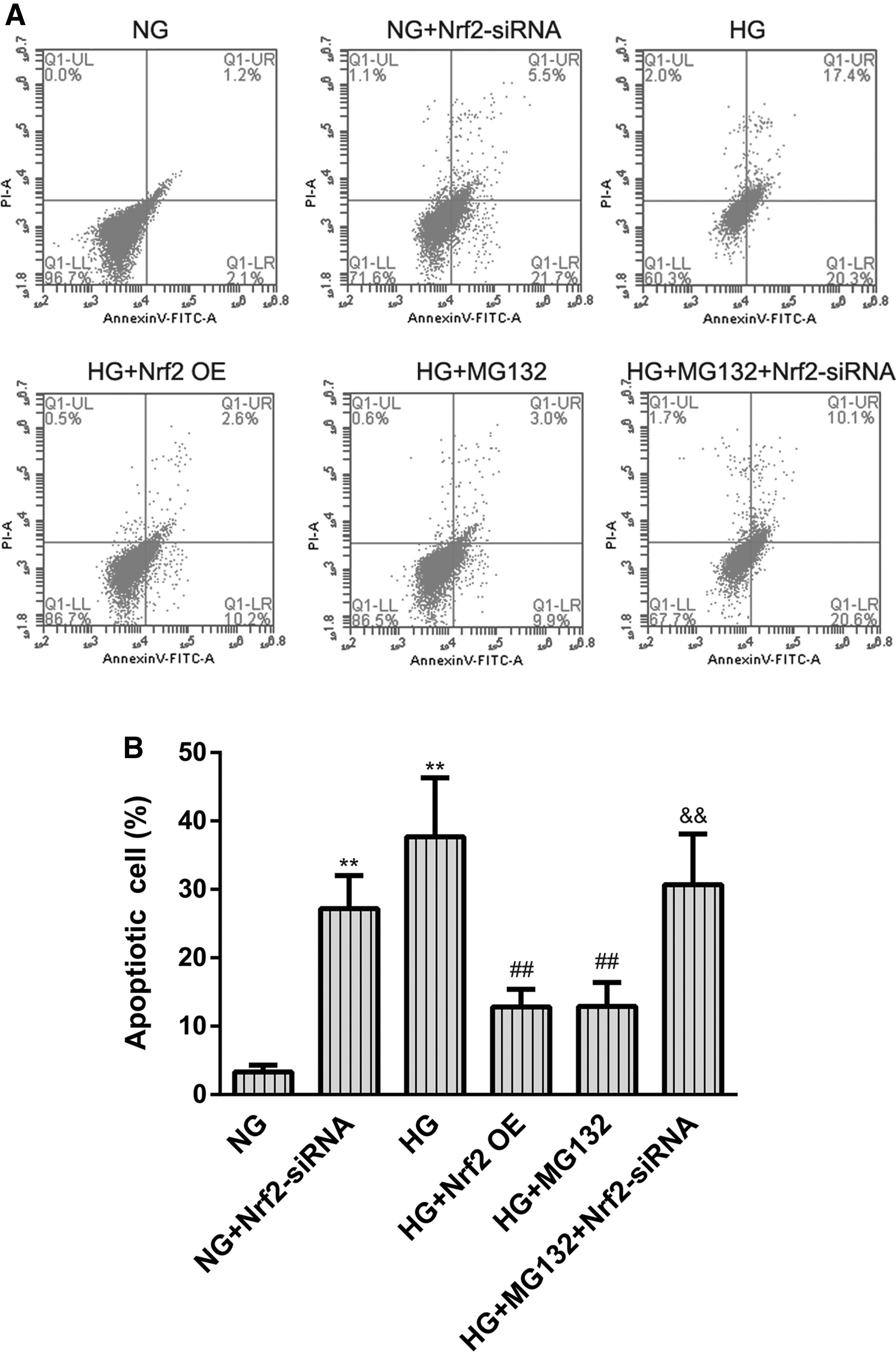
The role of Nrf2 in MG132 treatment of HG-induced HREC apoptosis.
To determine whether MG132 reverts hyperglycemia-induced retinal capillary cell apoptosis through targeting Nrf2. We also evaluated Nrf2 knockdown or overexpress, respectively, regulates MG132-mediated hyperglycemia-induced retinal capillary cell apoptosis, as shown in Fig. 8. The addition of Nrf2 siRNA significantly increased the percentage of apoptosis of retinal capillary cells compared with NG-treated cells (27.21% ± 4.83%, P < 0.01 compared with NG); conversely, Nrf2 overexpression significantly reduced HG-induced retinal capillary cell apoptosis (12.80% ± 2.60%, P < 0.01 compared with HG). Moreover, Nrf2 siRNA also dramatically increased the percentage of apoptosis of retinal capillary cells compared with HG combined with MG132-treated cells (30.70% ± 7.40%, P < 0.01 compared with HG+ MG132).
Discussion
In the current study, we provide evidence that administration of MG132, a proteasomal inhibitor with vasoprotective properties, to a rat model of diabetes and cultured HRECs, can effectively suppress the degradation of Nrf2 through suppression of 26S proteasomal activity. We also demonstrate that this action upregulates the levels of phase II antioxidant enzymes, such as HO-1 and NQO1.These changes were correlated with the inhibition of oxidative stress, and accompanied by a reduced vascular permeability of blood vessels in the diabetic retina.
DR is the most common complication of diabetes and is a leading cause of blindness among working-age adults. 21 Retinal vascular lesions are a major cause of vision loss in DR; occurring at early stages and reflecting a breakdown of the BRB. 2 There is currently no effective treatment with which to prevent the development of the retinal vascular lesions that are induced by diabetes. The identification of an effective treatment for this complication of DR would be an important advance. In this report, we focused on MG132, a nonspecific proteasomal inhibitor, is able to efficiently degrade intracellular proteins with high selectivity, which affects cell cycle regulation, apoptosis, antigen presentation, inflammatory reactions, and gene transcription.22–24 Many studies have shown that MG132 has therapeutic effects on diabetic nephropathy and diabetic cardiomyopathy,6–8,25,26 but the potential beneficial effects of MG132 on BRB are still controversial.
In this study, to evaluate the impact of MG132 in vivo, we applied it to the well-characterized STZ rat model with diabetes. In these studies, we found that MG132 alleviates the BRB leakage observed in STZ-treated rats. The inhibition of vascular lesions by MG132 was independent of a change in blood glucose, as this measure (and body weight) was comparable in STZ rats that were administered MG132. Instead, our results indicate that MG132 reduces the production of mitochondrial ROS in response to hyperglycemia, which is a key initiating step in the pathogenesis of diabetes, including DR. While the mechanism by which MG132 acts on intracellular ROS production is unclear. It has been demonstrated that, the ubiquitin proteasome pathway plays a pivotal role in the cellular response to oxidative stress.27,28 As a key component in the UPS, the 26S proteasome functions to recognize, unfold, and ultimately remove oxidatively damaged proteins, and is of critical importance to maintain normal cell homeostasis and viability.24,29–31 Our studies indicate that, MG132 inhibits the elevated 26S proteasome activity that is caused by diabetes, which were similar to those previously reported.32,33 Therefore, these reports raise the possibility that a mild reduction of 26S proteasome activity by MG132 can serve as a cellular defense mechanism to protect against oxidative stress during DR.
Then, we investigated the mechanism by which MG132 could regulate oxidative stress as a 26S proteasome inhibitor. We hypothesized that MG132 could disrupt basal ubiquitin-dependent degradation of Nrf2 by the 26S proteasome, leading to nuclear Nrf2 accumulation and activate Nrf2-targeted antioxidant gene induction. Nrf2 is considered to be one of the most important transcription factors associated with oxidative stress and can modify the basal and inducible expression of several antioxidant genes,11,34_ENREF_30. Under physiological conditions, Nrf2 locates in the cytoplasm and combines with its inhibitor kelch-like ECH-associated protein 1 (Keap1), which could mediate a rapid ubiquitination and subsequent degradation of Nrf2 by the proteasome35–37 _ENREF_33. While under conditions of severe stress, such as oxidative stress or diabetes, Nrf2 is free from Keap1 and translocates into the nucleus to bind to AREs in the genes encoding antioxidant enzymes, such as NQO1, glutathione S-transferase, HO-1, and γ-glutamylcysteine synthetase, increasing their expression to play a role of detoxification and antioxidation.38–42
We propose that MG132 may activate Nrf2 through decreasing the degradation of Nrf2 protein, and the activation of the following Nrf2-ARE pathway as a probable mechanism underlying the protective effect of proteasome inhibitors, warrants further investigation. It has been demonstrated that the inhibitory effect of MG132 on 26S proteasome activity can induce the accumulation of Nrf2, because Nrf2 is an ubiquitin–proteasome substrate.43,44 In this study, we found that MG132 upregulated intracellular Nrf2 protein level and activated the Nrf2-ARE pathway. Furthermore, we showed that MG132 did not increase Nrf2 mRNA levels. These findings are consistent with the fact that Nrf2 protein level increased by MG132 seems to be the result from a post-transcriptional mechanism, rather than of an increase in Nrf2 transcription. Furthermore, we demonstrated that the activation of Nrf2 and its downstream targets NQO1 and HO-1 could improve metabolic disorder and alleviate retinal damage in STZ-induced diabetic rats and revert hyperglycemia-induced retinal capillary cell apoptosis. This study conclusively indicates the requirement of Nrf2 for MG132-induced retinal protection against diabetes. 44
In summary, the present data demonstrate that MG132 was observed to reduce overactivated 26S proteasome and diabetes-induced oxidative stress through Nrf2/ARE pathway, which prolongs the impact of early vascular dysfunction on the progression of retinal endothelial injury. This study provides insight into the application of MG132 as an effective treatment for the early stages of progression in DR.
Footnotes
Authors' Contributions
Conceptualization: J.L. Data curation: P.W., S.F., and J.L. Formal analysis: S.F. and J.Y. Funding acquisition: J.Y. and J.L. Investigation: X.H. and L.L. Methodology: S.F. and J.L. Project administration: X.H., L.L., and J.L. Resources: J.L. Supervision: J.L. Validation: P.W. and J.Y. Visualization: P.W, J.Y., and J.Y. Writing—original draft preparation: P.W. and X.H. Writing—review and editing: S.F. and JL.
Author Disclosure Statement
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the article; or in the decision to publish the results.
Funding Information
This work was supported by the Health Development Planning Commission Science Foundation of Zhejiang Province (2020364455 and 2022KY1434).