
Abstract
Purpose:
Although it is now understood that most antiglaucoma surgeries fail because of scarring of the filtering tract, the underlying mechanism remains to be elucidated. The present study investigated the mechanism by which the interleukin (IL)-22/IL-22 receptor alpha 1 (IL-22RA1) signaling pathway regulates scar formation in glaucoma patients.
Method:
A total of 31 glaucoma patients who underwent trabeculectomy surgery with uncontrollable intraocular pressure because of scarring and 19 strabismus patients as the control patient group were included in the present study. ELISA was performed to measure the content of IL-22 in peripheral blood. Serum from patients was used to incubate human Tenon’s capsule fibroblasts (HTFs) cells and IL-22 antibody rescued the effect of IL-22 on the biological functions. qPCR and Western blot were performed to determine IL-22RA1 mRNA and protein expression levels. Flow cytometry was performed to assess the cell cycle distribution and the Cell Counting Kit-8 assay was used to analyze cell proliferation.
Results:
The ELISA assay revealed that the serum IL-22 level of glaucoma patients was significantly higher than the healthy group (29.80 ± 5.1 ng/µL vs. 5.21 ± 0.9 ng/µL). After incubation with patient serum, the proliferation and activation of human Tenon fibroblasts (HTFs) were promoted. IL-22 mediated the biological function of HTFs via directly binding IL-22RA1. Moreover, transfection of the siR-IL-22RA1 or IL-22RA1 gene resulted in significant antifibrosis or profibrosis in HTFs. When a signal transducer and activator of transcription (STAT) 3 inhibitor (BAY) was introduced to the IL-22RA1 overexpression group, IL-22-induced proliferation was reduced in HTFs. Additionally, glaucoma patients had increased levels of IL-22 expression following surgery.
Conclusions:
The IL-22/IL-22RA1/STAT3 signaling pathway promoted fibroblast cell proliferation and alpha-smooth muscle actin, potentially regulating fibrosis in glaucoma filtration tracts. Our results provide hitherto undocumented insights into the pathophysiology of postoperative scarring.
Introduction
It is well-established that glaucoma causes blindness accounting for 3.8% of all blind people between the age of 40 and 80 years in the world. 1 The number of glaucoma patients over 40 has exceeded 9.4 million, of which 5.2 million are monocular blindness and 1.7 million are blindness in both eyes, and the number is expected to reach 111.9 million in 2040. 2 Glaucoma is the leading cause of irreversible eye disease that can cause blindness and disability, with a significant impact on the quality of life for patients. To date, intraocular pressure control remains the mainstay of treatment, and surgery is often considered when medication and laser therapy fail to lower ocular pressure. 3
The current surgical treatments for glaucoma encompass trabeculectomy, aqueous humor drainage implantation, and minimally invasive glaucoma surgery (MIGS). In recent years, many types of MIGSs have emerged. However, procedures for internal drainage of aqueous humor to Schlemm’s canal cannot reduce intraocular pressure below the level of episcleral venous pressure. 4 Besides, external drainage procedures require patients to face stent pigment release, pigment obstruction, and wound scarring formation. 5 From a long-term point of view, trabeculectomy is still a recognized surgical method for lowering intraocular pressure, providing patients with reliable and durable effective intraocular pressure. However, postoperative scarring of the filtering tract, resulting in uncontrolled intraocular pressure and optic nerve damage, is still a vital cause of blindness in glaucoma patients.
Transforming growth factor-β (TGF-β), the main inflammatory factor in the filtration tract, stimulates the proliferation of human Tenon’s capsule fibroblasts (HTFs) and triggers their transition into myofibroblasts. Moreover, HTFs synthesize and secrete a large amount of extracellular matrix (ECM), leading to the development of scar tissue at the wound site. 6 During this process, a sustained immune response and the release of inappropriate immune mediators play an important role.
The expression of immune factors is increased postoperatively, such as interleukin (IL)-1β, IL-6, IL-8, IL-17, IL-22, etc. 7 IL-22 belongs to the IL-10 cytokine family and is among the cytokines that respond immediately to tissue damage. 8 It is mainly secreted by CD4+ T cells, Th cells, and Th22 cells and is the only known immune factor secreted by immune cells, mainly acting on nonimmune cells. 9 As a necessary factor for the repair of nonimmune cells (epithelial cells, mesenchymal cells, fibroblasts), IL-22 affects the wound healing process. It plays an important biological function after binding to the IL-22 receptor complex, a heterodimeric transmembrane receptor composed of interleukin 22 receptor alpha 1 (IL-22RA1) and interleukin 10 receptor 2 (IL-10R2) composition. IL-22 can reportedly stimulate cells expressing IL-22RA1 by increasing the phosphorylation level of Janus protein tyrosine kinase (JAK) and signal transducer and activator of transcription (STAT) 3. This signaling pathway is involved in various cellular processes such as cell proliferation, differentiation, apoptosis, and profibrotic effects. 10 Previous studies have shown that the expression of IL-22RA1 in the fascia tissue of the filtration tract in patients with glaucoma after surgery is increased. 11 However, the mechanism underlying the effect of IL-22 on filtration tract scarring remains unclear. Therefore, to study filtration tract scarring formation and find new therapeutic targets, it is important to understand the characteristics of IL-22 expression and its binding to receptors and analyze the molecular mechanism of downstream signaling pathways.
Methods
Patient and tissue collection
Serum and fascia tissue were collected from 31 glaucoma patients (GP group) and 19 strabismus patients as control patient group (CP group) from November 2021 to March 2022 in Changsha Aier Eye Hospital. The criteria for selecting patients were: GP group included glaucoma patients initially diagnosed with mediate primary open-angle glaucoma that was already treated with trabeculectomy surgery, and owing to scar formation, the intraocular pressure was uncontrolled; CP group included strabismus patients without other ocular diseases; all patients with no immune diseases, for example, ulcerative colitis, atopic dermatitis, and rheumatoid arthritis, and no recent history of acute infection in the past month. Peripheral blood (5 mL) of all the patients was collected from median cubital veins. By centrifuging at 600g and 4°C for 5 min, serum was separated for ELISA test and incubating cells. The fascia tissue samples were snap-frozen in liquid nitrogen and then stored at −80°C for further use. All procedures conducted in this study were performed in compliance with rules described in the Declaration of Helsinki. All patients provided their written informed consent prior to surgery and tissue collection. The study protocol was approved by the Human Ethics Committee of Changsha Aier Eye Hospital (Changsha, China) and the ethics number is KYPJ012.
Measurement of serum cytokine level
The serum level of IL-22 in the patients was measured by ELISA kits (Quantikine ELISA kits, R&D Systems; Minneapolis, MN, USA). The absorbance of each well was measured with a microplate reader (Multiskan FC, Thermo Fisher Scientific, Waltham, MA, USA) set to 450 nm, with the wavelength correction set to 540 nm.
Cells culture
HTFs were isolated from individuals undergoing strabismus surgery who had no history of conjunctival disease or use of topical ocular medication as previously described. 11 The HTFs were cultured in Dulbecco’s modified Eagle medium supplemented with 10% (v/v) fetal bovine serum (Hyclone Laboratories, Logan, UT, USA) and 1% streptomycin–penicillin (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) in a humidified atmosphere at 37°C with 5% CO2. HTFs cultured in above medium were termed as complete medium group (CM group). Passages 4–6 were used for further experiments. Prior to each experiment, the cells were allowed to reach a subconfluent status (∼80% confluence), after which, they were cultured in serum-free medium for 24 h.
To investigate the effect of patient serum on HTFs, serum from GP group or CP group was mixed with medium at a ratio of 1:10 to treat HTFs cells. Each experiment was performed more than three times on cells of different passages (n > 3). For IL-22 experiments, cells were exposed to different dose-course or time-course of human IL-22 (Peprotech, Rocky Hill, NJ, USA). In order to study the rescue effecting, cells were treated with human IL-22 antibody (1:800, ab133545, Abcam, Cambridge, UK).
Cell transfection
The nucleotide sequences that encoded for IL-22RA1 were amplified via PCR performed with the following primers: F, 5′-GGGGTACCATGAGGACGCTGCTGACCATCTTGA-3′; R, 5′-CCGCTCGAGTCAGGACTCCCACTGCACAGTCAGG-3′. Those sequences were then subcloned into the KpnI and XhoI sites of a pcDNA 3.0 vector. An empty pcDNA 3.0 vector was used as a control. siRNA-NC, siR-IL-22RA1 were obtained from GenePharma (Suzhou, China). HTFs were seeded into the wells of six-well plates and cultured to ∼80% confluence. Transfections were performed using Lipofectamine 3000 (Thermo Fisher Scientific, Waltham, MA, USA) as described in the manufacturer’s instructions.
Flow cytometry
According to the manufacturer’s manual, flow cytometry performed cell cycle analysis. First, preparation cell with DNA Reagent kit (BD Biosciences, San Jose, CA, USA). In brief, the harvested cells were incubated in sequence with solutions. Then, the cells were analyzed using a flow cytometer and ModFit software (v3.2; Verity Software House, Inc.). Each experiment was repeated three times.
Cell viability assay
Cell viability was measured by a Cell Counting Kit-8 (CCK-8) (Lianke Bio, Hangzhou, Zhejiang, China) according to manufacturer’s instructions. In brief, cells were seeded at a density of 1 × 104 cells/well in a 96-well plate triplicated. After the cells with specified treatment, a CCK-8 working reagent was gently added to each well and then placed in a 37°C incubator for 2 h. Finally, the absorbance of each well at 450 nm was measured with a microplate reader (Multiskan FC, Thermo Fisher Scientific, Waltham, MA, USA).
RNA extraction and PCR analysis
RNA was extracted from HTFs cells using Trizol reagent (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). PowerUP SYBR Green Master Mix kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to detect mRNA expression of IL-22 (F 5′-CAACAGGCTAAGCACATGTCATATT-3′; R 5′-TCTCTCCACTCTCTCCAAGCTTT-3′), IL-22RA1 (F 5′-CCTGATGTGACCTGTATCTCCAA-3′, R 5′-GGTCAGGcCGAAGAACTCATATT-3′), α-smooth muscle actin (α-SMA) (F 5′-GAGACCACCTACAACAGCATCAT-3′, R 5′-GCCGATCCACACCGAGTATTT-3′), and GAPDH (F 5′-GGAGTCCACTGGCGTCTTCA-3′, R 5′-GTCATGAGTCCTTCCACGATACC-3′) according to the manufacturer’s protocol. Resulting data were analyzed using the comparative cycle threshold (Ct) method. The target gene cycle thresholds were adjusted relative to a calibrator (normalized Ct value obtained from control groups) and expressed as 2−ΔΔCt (Applied Biosystems User Bulletin no. 2: Rev B “Relative Quantitation of Gene Expression”).
Protein preparation and Western blot analysis
Total protein was lysed by cold RIPA buffer. Then, a BCA assay kit (Thermo Fisher Scientific, Waltham, MA, USA) measured the protein concentration of the supernatant. For Western blot experiments, IL-22 antibody (1:800, AF782, R&D Systems; Minneapolis, MN, USA), IL-22RA1 antibody (1:800, ab5984, Abcam, Cambridge, UK), α-SMA antibody (1:1000, ab28052, Abcam, Cambridge, UK), STAT3 antibody (1:800, ab68153, Abcam, Cambridge, UK), and p-STAT3 antibody (1:800, ab267373, Abcam, Cambridge, UK) were used to detect certain protein, respectively, whereas β-actin antibody (1:5000, ab8227, Abcam, Cambridge, UK) was used as a 1:5,000 dilution to determine β-actin protein abundance. Next the membrane was incubated with HRP-conjugated goat antirabbit or goat antimouse secondary antibodies. ECL Ultra Western HRP kit (Thermo Fisher Scientific, Waltham, MA, USA) was used to detect areas of luminescence, and the relative-staining intensity of each protein band was quantitated using ImageJ software (Ver. 6.0, Media Cybernetics, Inc., Rockville, MD, USA). A ratio of IL-22, IL-22RA1, STAT3, p-STAT3, and α-SMA protein intensity over β-actin protein intensity was used to quantify protein expression.
Tissue histological observation and immunohistochemistry staining
Tissues fixed in 10% formalin were embedded in paraffin and sectioned. Eight-micrometer-thick sections were cut and stained. Briefly, IL-22RA1 or α-SMA was incubated with tissue sections at a 1:200 dilution overnight. The tissue slides were subsequently incubated with secondary antiserum (Alexa Fluor 647 goat antirabbit IgG; Alexa Fluor 567 goat antimouse IgG Molecular F Probes, Eugene, OR, USA) at 1:400 dilution.
Immunofluorescent staining of fibroblasts cells was performed on microcoverslips in 12-well tissue culture plates. Cells were serum-starved for 24 h and then treated with patients’ serum medium or 20 ng/mL recombinant IL-22 for 24 h. Cells were washed and fixed in 4% formaldehyde, then stained with IL-22RA1 or α-SMA following incubation with required secondary antibodies. Their nuclei were stained with DAPI. Image acquisition was performed using a Zeiss confocal microscope or an upright Zeiss microscope with Axiovision software.
Ethical statement
For all articles reporting data from studies involving human participants or animals, formal review and approval, or formal review and waiver, by an appropriate institutional review board or ethics committee is required and should be described in this subsection. For those investigators who do not have formal ethics review committees, the principles outlined in the Declaration of Helsinki should be followed.
Statistical analysis
All experiments were performed in triplicate and the data were analyzed using SPSS Software (Version 23.0, IBM Corp, Armonk, NY, USA). Results were presented as the mean ± SD. Differences between groups were analyzed using the t-test or ANOVA followed by the Tukey test. For all statistical tests, a P value <0.05 was statistically significant.
Results
IL-22 expression is elevated in peripheral blood from patients with postsurgical scarring
The serum IL-22 levels were significantly elevated in glaucoma patients in the postsurgical scarring (GP) group compared with the CP group (29.80 ± 5.1 ng/µL vs. 5.21 ± 0.9 ng/µL) (Fig. 1A). To determine whether IL-22 might play a role in filtering tract scarring, we analyzed the mRNA and protein expression of IL-22. As shown in Fig. 1C, IL-22 protein expression was significantly higher in tissues from the patient group than in the healthy group (P < 0.05). Consistent findings were observed for mRNA expression (Fig. 1B).
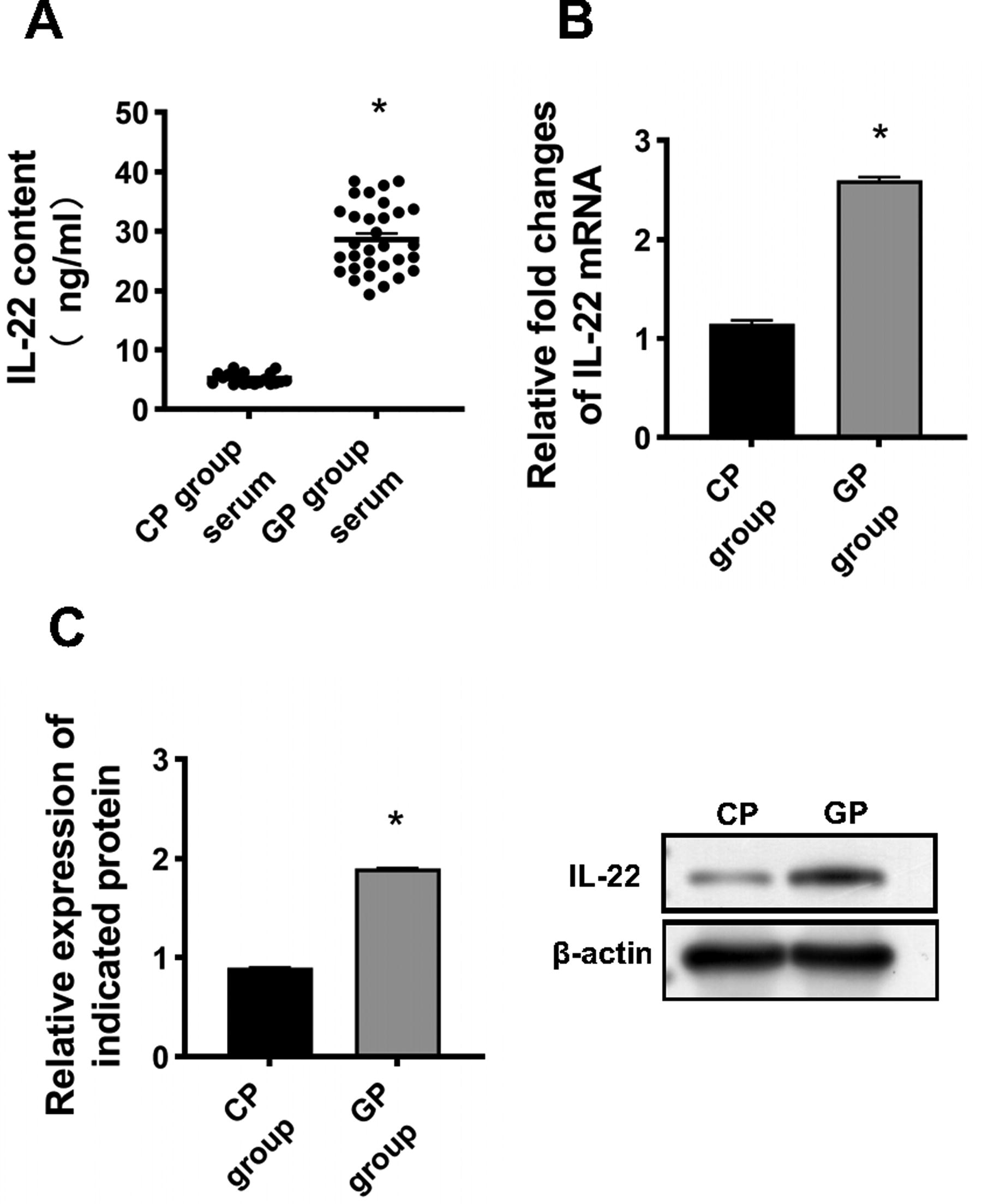
IL-22 was elevated in patients with postsurgical scar.
Expression of IL-22RA1 is elevated in postsurgical scar tissue
The specificity and sensitivity of IL-22 are dependent on IL-22RA1 expression and localization. It has been established that IL-22RA1 is widely distributed, especially in Tenon’s capsule fibroblasts. To assess the underlying mechanisms, we detected IL-22RA1 expression by PCR and Western blotting and localized IL-22RA1 protein by immunohistochemical techniques in postsurgical scar tissue. As shown in Fig. 2A, IL-22RA1 mRNA expression was higher in scar tissue than in healthy tissue. Western blot consistently confirmed IL-22RA1 protein expression in scar tissue lysates from antiglaucoma surgery patients and healthy patients (Fig. 2B). The localization of IL-22RA1 in tissue was detected using confocal microscopy, which showed that it was localized to the plasma membrane (Fig. 2C). α-SMA is a marker of HTFs activation. As shown in Fig. 2, α-SMA was elevated in postsurgical scar tissue at the mRNA and protein levels.
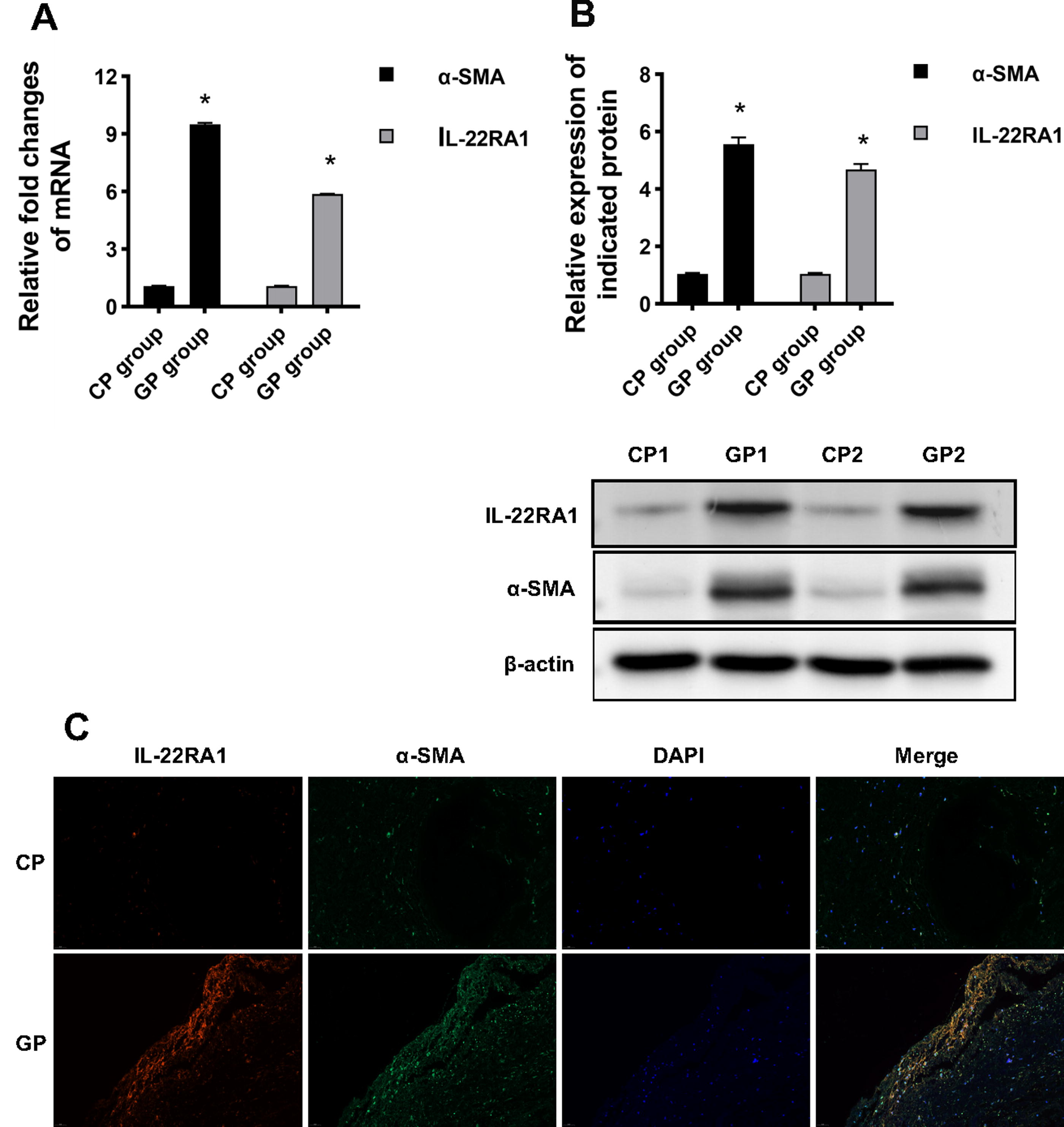
Expression of IL-22RA1 and α-SMA was increased in patients with postsurgical scar.
Glaucoma patient serum stimulates HTFs proliferation and activation
After incubation with healthy or glaucoma patient serum for 24 h, the levels of IL-22RA1 mRNA in HTFs were determined by RT-qPCR. The expression of IL-22RA1 mRNA in HTFs treated with serum from the GP group was significantly higher than in untreated HTFs (CM) (P < 0.05), and there was no significant difference between the control group and the healthy serum group (CP group) (Fig. 3A). As shown in Fig. 3B, the proliferation of HTFs at 0, 12, 24, 48, and 72 h after serum treatment was significantly higher than that of untreated cells (P < 0.05), and there was no significant difference between the control group and the healthy serum group (P < 0.05).

Incubation with patient serum induced the expression of IL-22RA1 in HTFs cells.
The proportion of G1 phase cells of HTFs treated with patient serum was significantly lower than the control group (P < 0.05; Fig. 3C), and there was no significant difference in the proportion of G1 cells between the control group and the healthy serum group. Moreover, the proportion of S-phase cells in HTFs treated with serum from glaucoma patients was higher than in the control group, and there was no significant difference between the control and healthy serum groups.
In addition, it has been established that patient serum plays an important role in HTFs activation. The mRNA and protein expression levels of IL-22RA1 and α-SMA were significantly higher than in the control group (P < 0.05; Fig. 3A, D), whereas protein expression was comparable between the control group and the healthy serum group.
IL-22 mediates the biological function of HTFs
To further investigate whether IL-22 regulates the proliferation and activation of HTFs, the IL-22 antibody was used to conduct rescue experiments. After adding the IL-22 antibody, the proliferative capacity of HTF cells was reduced in patients’ serum compared with the healthy serum group, which was like that in control group (Fig. 4A). Furthermore, as shown in Fig. 4B, the addition of IL-22 antibody to the serum group inhibited the cell cycle progression from the G1 to the S phase, resulting in a pattern similar to that of the control group. Additionally, the IL-22 antibody group showed significantly lower levels of IL-22RA1 protein expression compared with the patient serum group (P < 0.05, Fig. 4C) and levels were similar to those found in the control group. In addition, the protein expression of α-SMA in the IL-22 antibody group was significantly lower than in the patient serum-treated group (P < 0.05; Fig. 4C). In summary, these findings indicate that the administration of patient serum increased the expression of IL-22RA1 in HTFs, whereas the use of IL-22 antibody reversed the effects and restored IL-22’s biological functions, allowing direct HTF proliferation and activation.

IL-22 antibody-protected HTFs cells.
IL-22 exerts its function by binding to IL-22RA1
To assess whether IL-22 transmits intracellular signals through IL-22RA1, IL-22RA1 was knocked down or overexpressed in HTFs, and then subsequently stimulated with IL-22. Figure 5A shows the transfection efficiency of siR-IL-22RA1 and IL-22RA1. The expressions of IL-22RA1 and α-SMA in the siR-IL-22RA1 group were significantly lower than in the siR-NC group (P < 0.05). In addition, the expression of IL-22RA1 and α-SMA in the IL-22RA1 overexpression group was significantly higher than in the NC group (P < 0.05; Fig. 5B). These results indicated that the transfection experiments were successful. Compared with siR-NC and NC groups, IL-22-induced proliferation was inhibited in the siR-IL-22RA1 group but increased in the IL-22RA1 overexpression group. Compared with the siR-NC group and the NC group, the proportion of S-phase cells in the siR-IL-22RA1 group decreased, whereas the proportion of S-phase cells in the IL-22RA1 overexpression group increased (P < 0.05; Fig. 5C). These results indicated that IL-22 exerts its biological function through IL-22RA1.
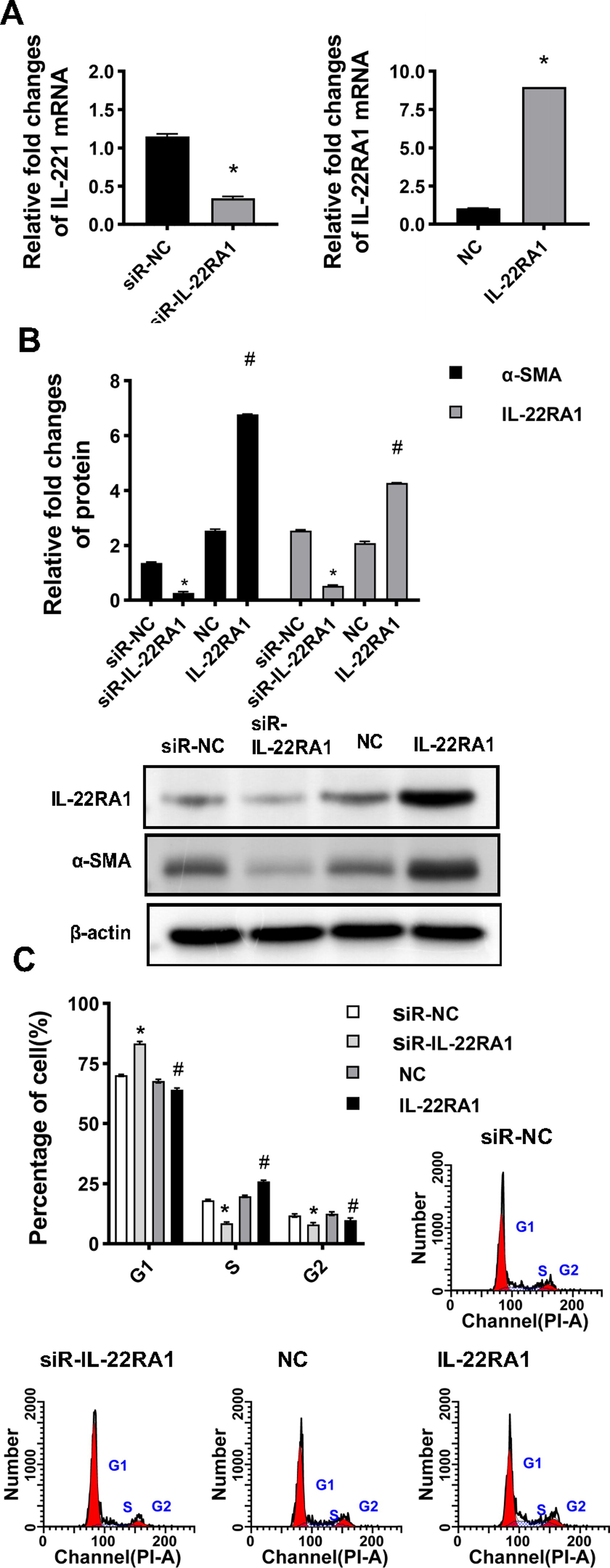
Silencing of IL-22RA1-protected HTFs cells.
IL-22/IL-22RA1 regulates the proliferation and activation of HTFs via the STAT3 signaling pathway
To further investigate how IL-22RA1 regulates the functions of fibroblasts through IL-22, this study investigated the effect of STAT3 signaling pathway inhibitor BAY (2 µM) on HTFs. The supplementation of BAY to the IL-22RA1 overexpression group reduced the IL-22-induced proliferation of HTFs (P < 0.05; Fig. 6A). Moreover, BAY limited the effect of IL-22 on α-SMA expression (P < 0.05; Fig. 6B). Flow cytometry results showed that cells treated with BAY reduced the effect of IL-22 on the proportion of cells transitioning from G1 to S phase (P < 0.05; Fig. 6C). Overall, these results suggest that IL-22 may regulate the proliferation and expression of α-SMA proteins in HTFs via the STAT3 signaling pathway.
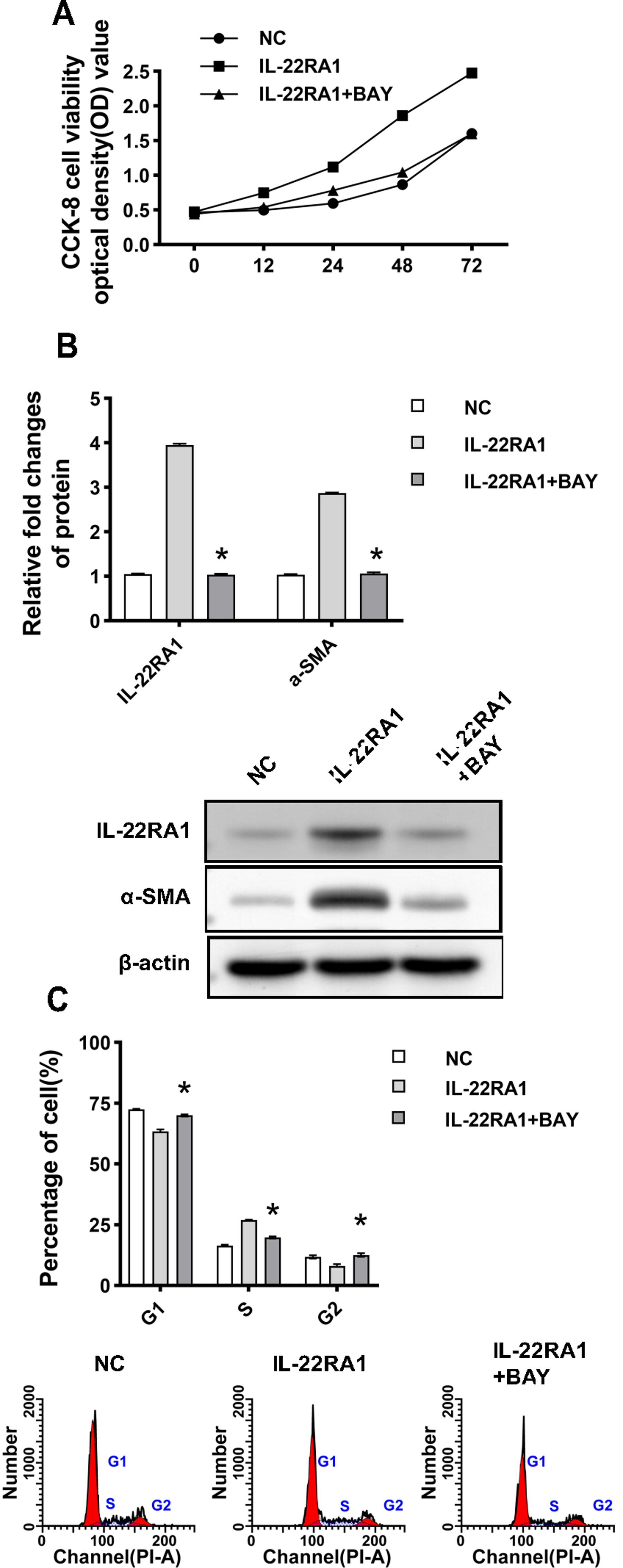
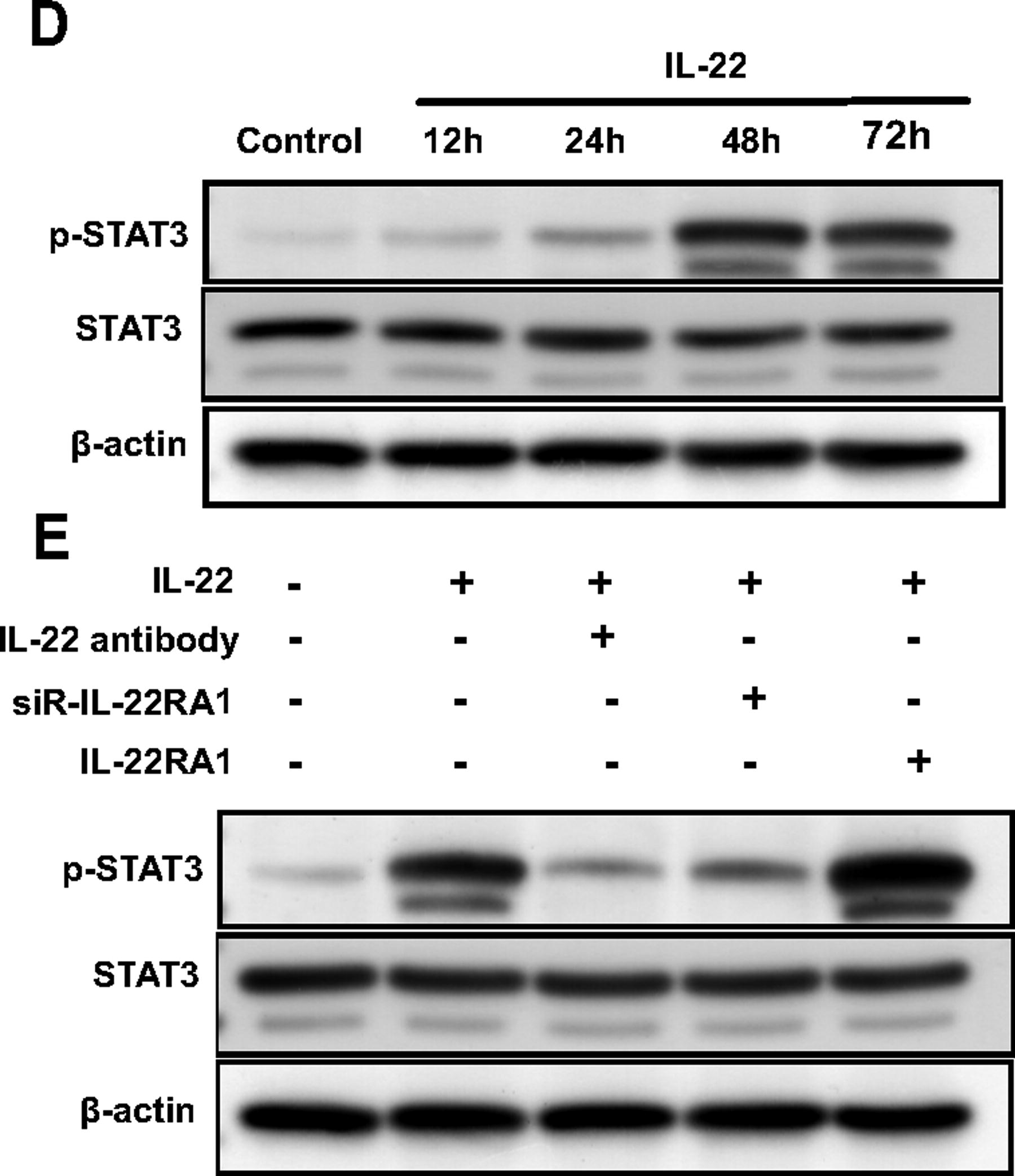
(Continued).
Furthermore, IL-22 induced STAT3 phosphorylation in HTFs. A time-course study showed that IL-22-induced STAT3 phosphorylation gradually increased but began to decrease at 72 h (Fig. 6D). To further assess the role of IL-22RA1 in IL-22 signaling, we investigated the effect of IL-22-induced STAT3 phosphorylation using anti-IL-22 antibody, siR-IL-22RA1, and IL-22RA1. As shown in Fig. 6E, treatment with anti-IL-22 antibody and siR-IL-22RA1 resulted in 56% and 83% inhibition of IL-22-induced STAT3 phosphorylation in HTFs, respectively. Moreover, IL-22RA1 transfection increased STAT3 phosphorylation.
Discussion
After glaucoma surgery, cells around the wound can detect various extracellular environmental signals, including chemical and mechanical signals. Mechanical signals that cells can sense from their surrounding environment include physical forces such as tension, compression, and shear. By the changes, the dynamic balance of hydrophobic and lipid–lipid interactions of the lipid bilayer membranes, mechanosensitive Ca2+ permeating channels Piezo1, 12 and transient receptor potential vanilloid 4 13 were activated, which regulated macrophages and fibroblasts accumulation. Chemical stimulations mainly come from TGF-β, MMPs, and IL-1β. Fibroblasts have been activated and overproducing proteins of the ECM. Besides promoting scar formation, fibroblasts can crosstalk with inflammatory macrophages. 14 Therefore, the interaction between fibroblasts and macrophages may be an important early event in the recruitment of monocytes and may facilitate a cytokine network that maintains the activation of tissue inflammation.
In various tissues, IL-22 stimulates inflammatory responses, wound healing, and tissue regeneration. By activating the STAT3 pathway while promoting macrophage M2 polarization, IL-22 affects inflammatory and fiber proliferation process. 15
It is widely thought that HTFs from glaucoma patients express high levels of IL-22RA1, and this expression increases with scarring. In fibroblasts, IL-22 binds to IL-22RA1, activates downstream STAT3 phosphorylation, and triggers the expression of fibronectin and collagen. In psoriatic dermal fibroblasts, IL-22 binds to IL-22RA1 and stimulates the activation of the JAK/STAT pathway, resulting in increased production of ECM, fibronectin, and collagen. 16 Moreover, both IL-20 and IL-22 are members of the IL-10 family. It has been shown that in trabecular cells of glaucoma patients, IL-20 binds to the receptor IL-20 receptor A and participates in ECM, remodeling through the JAK/STAT signaling pathway, leading to increased intraocular pressure. 17
Our study found that STAT3 inhibitors inhibited the effect of the IL-22/IL-22RA1 signaling pathway on the proliferation of HTFs and the production of α-SMA. HTFs induced STAT3 phosphorylation in response to IL-22, directly correlating with the function of IL-22RA1. Furthermore, both overexpression and blockade experiments demonstrated that IL-22RA1 was required for STAT3 phosphorylation. These results suggest that IL-22/IL-22RA1 may promote fibroblast proliferation and activation through the STAT3 signaling pathway during postoperative filtering tract fibrosis. The exact mechanism is not completely understood and requires further investigation.
Wound healing is an important process in the formation of functional filtering blebs after glaucoma surgery. Fibroblasts transdifferentiate into myofibroblasts, causing wound contraction and formation of posterior parallel fiber bundles. This change affects tissue regeneration. 18 Even under optimal conditions, healing often results in fibrotic scarring. 19 Therefore, understanding how to shift wound repair from scarring to regenerative outcomes remains a challenge for researchers in the field. In this regard, progress has been hampered by the lack of suitable animal models of scar-free wound healing under natural conditions. Finding effective strategies to reduce scarring and achieve superior regenerative wound healing are next steps in research. The role of IL-22 in glaucoma has been largely underinvestigated. Overall, our study provides novel insights into understanding antifibrosis mechanisms after surgery. Serum IL-22 levels represent a potential marker to guide the postoperative management of this patient population.
Footnotes
Acknowledgment
The authors wish to acknowledge the excellent facility of Aier Eye Institute.
Authors’ Contributions
All authors contributed equally to this work. X.D. and Y.Z. designed the experiments. Y.Z., X.Z., and X.Z. performed the experiments. X.Z., and B.C. performed the statistical analysis. Y.Z. and X.D. wrote the article. All authors have read and approved the final version of the article.
Disclaimer
Opinions, interpretations, and recommendations are not necessarily endorsed by Baihua Chen from the Department of Ophthalmology in The Second Xiangya Hospital.
Author Disclosure Statement
All the authors of this article state that no potential conflicts, either personal or financial, exist.
Funding Information
This work was funded by the National Natural Science Foundation of China (Grant No. 81970801 to X.D.), the Natural Science Foundation of Hunan Province, China (Grant Nos. 2024JJ9021 to Y.Z. and 2023JJ70014 to X.D.), Hunan Engineering Research Center for Glaucoma with Artificial Intelligence in Diagnosis and Application of New Materials (Grant No. 2023TP2225 to X.D.), the Science and Technology Foundation of Aier Eye Hospital Group, China (Grant No. AR2206D5 to X.D. and Aier Glaucoma Institute), and the Science and Technology Foundation of Aier Eye Hospital Group, China (Grant Nos. AR2106D1 and AR2206D3 to Y.Z. and Aier Glaucoma Research Institute). Changsha Municipal Natural Science Foundation (No. kq2208495 to X.D.).