
Abstract
Purpose:
Glaucoma is a leading cause of irreversible blindness. Glaucomatous intraocular pressure (IOP) triggers deleterious effects, including gliosis, optic nerve (ON) axonal retraction, neurotrophic factor deprivation, inflammation, and other pathological events, leading to retinal ganglion cell (RGC) loss. Trophic factor impairment enhances RGC apoptosis susceptibility. Neuritin 1 (NRN1), a neurotrophic protein downstream of various neurotrophins, exhibited RGC protection and regeneration in axotomy models. We evaluated human recombinant NRN1's impact on human RGCs cultured in pressurized conditions within the ex vivo translaminar autonomous system to simulate glaucoma pathogenesis.
Methods:
Human glaucomatous and non-glaucomatous donor eyes were obtained from eye banks according to the Declaration of Helsinki. Initially, we evaluated NRN1and RGC marker expression in glaucoma and non-glaucomatous retina to determine the NRN1 level and its association with RGC loss. Further, we evaluated NRN1’s therapeutic potential by treating pressurized human eyes at normal and high IOP for seven days. Retina, ON, and conditioned medium were analyzed for RGC survival (THY1, RBPMS), gliosis (GFAP), apoptosis (CASP3, CASP7), and extracellular matrix deposition (COLIV, FN) by qRT-PCR and western blotting. Paraphenylenediamine staining assessed ON axonal degeneration, whereas ex vivo electroretinogram assessed retinal activity.
Results:
Glaucomatous retinas exhibited significant reductions in both NRN1 (*p = 0.007, n = 5) and RGC marker expression (*p = 0.04, n = 5). NRN1 treatment reduced gliosis, extracellular matrix deposition, ON degeneration, and increased retinal activity in pressure-perfused eyes.
Conclusions:
Our study confirms that NRN1 enhances human RGC survival and improves retinal function in degenerative conditions, substantiating it as a promising candidate for rescuing human RGCs from degeneration.
Introduction
Glaucoma, a progressive optic neuropathy, globally affects around 3.54% of individuals aged 40–80 years and 1.86% in the United States aged 40 years or older. 1 Elevated intraocular pressure (IOP) is the primary modifiable risk factor. 2 Despite IOP lowering, ongoing vision loss persists due to retinal ganglion cell (RGC) degeneration. Elevated IOP causes gliosis, RGC axonal transport deficits, and neurotrophic factor (NTF) impairment, 3 leading to RGC soma degeneration and RGC loss. 4 Therefore, lowering IOP is insufficient for preserving RGCs, highlighting the need for targeted therapy to safeguard RGC somas and axons.
An obstruction in NTF supply exacerbates RGC degeneration. For instance, major NTFs such as brain-derived neurotrophic factor (BDNF), glial cell line-derived neurotrophic factor (GDNF), and ciliary neurotrophic factor (CNTF) are downregulated in glaucoma.5–7 Animal studies evaluated multiple NTFs in glaucoma; recombinant BDNF preserved visual function in rodents, 8 whereas AAV2-mediated BDNF gene delivery protected RGCs long term by upregulating TrkB receptors. 9 The growth factor, CNTF, an extensively studied NTF, 10 has often showed variable outcomes such as inducing axonal outgrowth in neonatal RGCs 11 and not in mature RGCs. 12 Conversely, AAV2-mediated CNTF delivery was shown to promote RGC regeneration.13,14 The GDNF sustained delivery via microspheres enhanced long-term RGC survival. 15 However, identifying a factor that encompasses neuroprotective, neuroregenerative, and chemotactic properties is crucial for rescuing mature RGCs from glaucomatous degeneration and ensuring targeted regeneration.
Neuritin 1 (NRN1), a small glycosylphosphatidylinositol anchored protein, was first identified in the rat visual cortex. 16 It enhances neuronal activity and is potentiated by NTFs like BDNF, NGF, and NT-3, known for promoting neurite outgrowth and neuronal arborization.17,18 Our previous studies demonstrated the regenerative potential of AAV2-hNRN1 on axotomized mouse RGCs following optic nerve (ON) crush. 19 Intravitreal administration of AAV2-Nrn1 reduced RGC apoptosis and induced ON regeneration in an ON crush rat model. 20 Downregulation of NRN1 has been linked to increased RGC damage, inner retinal degeneration, and suppressed Akt and ERK phosphorylation. 21 The NRN1’s chemotactic capability was also validated in both neuronal cell lines and rat cortical interneurons from embryonic brain slice. 22 Although these studies provide valuable insight into the neuroprotective, chemotactic, and regenerative efficacy of NRN1, it is critical to evaluate NRN1’s protective effect on human RGCs within a pressurized ocular microenvironment as observed in glaucoma. This could be achieved using our ex vivo perfused pressurized translaminar autonomous system (TAS), which can simulate pressure-induced pathogenesis.23,24 Thus, by using the TAS model with human donor eyes at normal and high IOP, we aimed to test the therapeutic potential of NRN1. Our study demonstrated the beneficial effects of hNRN1 by reducing RGC apoptosis due to pressure-induced degeneration.
Materials and Methods
Human donor tissue collection and demographics
The methods adhere to Indiana University School of Medicine’s approved guidelines for Not Human Subject Research (NHSR) and to the Association for Research in Vision and Ophthalmology (ARVO) guidelines for handling de-identified human specimens. The ocular tissues were collected within 24–48 h postmortem from Lions World Vision Institute (Tampa) and Lions Gift of Sight (Minnesota) eye banks and processed according to the Declaration of Helsinki. Only postmortem eyes harvested with nonreactive serology and negative COVID-19 status were used. Donors with ocular infection, other ocular pathology, or progressive central nervous system diseases were excluded. Experiments were performed as four independent repeats.
Perfusion culture of human posterior eye cup
Human posterior segment culture experiments using donor eyes were performed in accordance with the Indiana University—Institutional Biosafety Committee approval (Proposal # IBC 1318). The posterior cups were perfused within the TAS model as described previously. 24 Briefly, eyes were washed with phosphate-buffered saline (PBS) and cross-sectionally dissected into anterior and posterior segments. A 1-diopter power soft contact lens (Acuvue) was gently applied over the retina to prevent retinal detachment. The posterior cup was placed on the IOP chamber of the TAS, with the ON facing upward and tightly sealed with epoxy resin O-ring and screws. The intracranial pressure (ICP) chamber was positioned above the ON and sealed. Inflow and outflow tubing were connected to corresponding portals of IOP and ICP chambers. Inflow syringes were filled with neurobasal media prepared as previously published.19,23,25 Briefly, the medium was prepared using penicillin (100U/mL, Cytiva), streptomycin (100 µg/mL, Cytiva), GlutaMAX (2 mM, Thermo Fisher Scientific), pyruvate (1 mM, Thermo Fisher Scientific), insulin (5 µg/mL, Gibco), transferrin (100 µg/mL, Gibco), sodium selenite (40 ng/mL, Gibco), progesterone (60 ng/mL, MilliporeSigma), putrescine (16 µg/mL, MilliporeSigma), thyroxine (100 ng/mL, MilliporeSigma), tri-iodothyronine (40 ng/mL, MilliporeSigma), forskolin (5 µM/mL, MilliporeSigma), and 1% human serum (Gibco) with basal neurobasal medium (Gibco) containing 10X B27 supplements (Gibco). In all experiments, the ocular sinister (OS, left eye) eye received hNRN1 treatment, whereas the paired eye, oculus dexter (OD, right eye) served as control. In the treatment group, hNRN1 was dissolved in complete neurobasal medium at a concentration of 200 ng/mL, a concentration known for RGC protection. 19 All eyes were perfused either at normal IOP or high IOP for seven days, guided by the observed pattern of 65% of RGC loss in axotomized eyes occurring within a 7-day timeframe. 26 The entire system was cultured in a CO2 incubator. Inflow syringes were connected to an automated pump with an adjusted flow rate of 0.3–0.5 µL/min for normal IOP (∼17.9 mmHg) and 1–2 µL/min for high IOP (∼28.2 mmHg). The outflow syringes were connected to manually calibrated hydrostatic pressure transducers linked to a multichannel bridge amplifier. Conditioned medium was collected every 48 h, and pressure data recorded every 24 h and averaged using LabChart software (ADInstruments). After seven days perfusion, tissues were harvested for downstream analysis.
Immunostaining of the donor retina
Glaucomatous and non-glaucomatous retinas underwent immunofluorescence staining for RBPMS and NRN1. Retinal flat-mounts were fixed in 4% formaldehyde for 15 min, washed with PBS, permeabilized with 0.5% Triton X-100 for 20 min, and blocked with SuperBlockTM T20 (Thermo Fisher Scientific) for 30 min. Primary antibodies (RNA binding protein, mRNA processing factor (RBPMS), Novus, Cat. No. NBP2-03905, 1:300, Neuritin1, Abcam, Cat. No. Ab64186, 1:500) were diluted with blocking buffer and incubated overnight at 4°C. Following PBS wash, retinas were incubated with secondary antibodies (goat anti-mouse IgG, Alexa Fluor 568, 1:300, goat anti-rabbit IgG, Alexa Fluor 488, 1:300, Invitrogen) for 2 h at room temperature. Retinas were mounted with ProLongTM Gold Antifade Reagent with DAPI (Thermo Fisher Scientific) and images captured at 10X magnification.
Quantitative gene expression analysis
RNA extraction was performed using the MACHEREY-NAGEL kit protocol. Retinal tissues (2 mm) were collected from glaucomatous (n = 5) and non-glaucomatous donors (n = 5), as well as from OD and OS eyes of pre- and post-perfusion (n = 4). The RNA (200 ng) was reverse transcribed to cDNA using iScriptTM kit (Bio-Rad Laboratories). The NRN1, RBPMS, and THY1 expression in glaucomatous and non-glaucomatous retinas were assessed by SYBR Green PCR using PowerUp™ SYBR™ Green Master Mix (Applied Biosystems), 200 nM of forward and reverse primers, and cDNA. The primers used were NRN1 Forward: 5′-TCCCCCGCGTTCTCTAAACT-3′, NRN1 Reverse: 5′-GCCCAGCTTGAGCAAACAGT-3′, RBPMS Forward: 5′-AAACAGCCTGTAGGTTTTGTCA-3′, RBPMS Reverse: 5′-GGAATTTCAGGATCGAAGCGG-3′, THY1 Forward: 5′-CTCCTGCTAACAGTCTTGC-3′, THY1 Reverse: 5′-CCAAAGAGCACGTGCTTCT-3′, and GAPDH Forward: 5′-ACAGTCAGCCGCATCTTC-3′, and GAPDH Reverse: 5′-ACGACCAAATCCGTTGACTC-3′. For assessing retina suitability for perfusion culture, pre-perfusion eyes cDNA were assessed for inflammatory (TLR4, GFAP, GSS, and AIF1), apoptotic (BAX, CASP3, and CASP7), and retinal markers (RBPMS, ISL-1, THY1, NEFH, RHO, and PRKCA). To assess hNRN1 effects, post-perfusion retinal cDNA was evaluated for apoptotic (CASP3 and CASP7), astrogliotic (GFAP), and RGC markers (RBPMS and THY1). TaqMan array qRT-PCR was used for assessing pre- and post-perfusion retinal gene expression, with reaction mixture comprised of TaqMan fast advanced master mix (Thermo Fisher Scientific, Cat. No. 4444558), cDNA, and primers in a pre-designed array plate. The qRT-PCR was performed (Applied Biosystems QuantStudio3 system) and fold changes (FC) were calculated using 2^-ΔΔct with GAPDH as the housekeeping control.
Western blotting for protein analysis
Conditioned medium proteins were concentrated using StrataClean Resin (Agilent) by adding 10 µL of resin to 1 mL of medium. Protein pellets were collected and concentrations determined by Bicinchoninic Acid Kit assay (MilliporeSigma). The SDS-PAGE was conducted with 20 µg of protein at constant voltage and proteins transferred to PVDF membranes using an Invitrogen wet transfer system. After PBS washing and 1 h blocking with SuperBlockTM T20 (Thermo Fisher Scientific), blots were incubated overnight at 4°C with primary antibodies against fibronectin (Millipore, Cat. No. AB 1945, 1:1000) and collagen IV (Novus, Cat. No. NB120-6586, 1:1000). After PBS wash, HRP-conjugated anti-rabbit IgG (Santa Cruz, Cat. No. sc-2004, 1:1000) was added for 1 h at room temperature. Blots were visualized using chemiluminescence, with Ponceau staining as a normalizing control.
Paraphenylenediamine (PPD) staining
The ON, 1 cm in length from the base of the ON head were dissected from pressure-perfused eyes and fixed in a 2% paraformaldehyde/2.5% glutaraldehyde mixture in 0.1M sodium cacodylate buffer. Sectioning and imaging of the ON were performed using an electron microscope facility at the University of Wisconsin, Madison. Before sectioning, tissues were rinsed with 0.1M sodium cacodylate and incubated overnight with 2% osmium. Tissues were washed in 0.1M sodium cacodylate, dehydrated, and embedded in BEEM (pre-shaped polyethylene moulds) capsules (Electron Microscopy Services). Tissue cross-sections, 0.75–1 µm thick, were taken from the distal portion and placed on a drop of water. They were heated to aid adherence to the slide. The sections were incubated with 1% PPD in isopropanol: methanol (1:10) for 10–30 min, followed by isopropanol washing. Axons were visualized under a phase-contrast microscope at 20X magnification. Ten cross-sections of the ON were taken from each perfused eye for the axon count. The axons were counted at superior, inferior, nasal, and temporal regions in each section. One ON axon in the normal IOP group was excluded owing to noncountable ON sectioning, limiting our sample size to three. Double masked manual counts of normal and degenerated axons were done for each sample.
Ex vivo electroretinogram (ERG) for measuring retinal function
The ex vivo ERG was performed as previously published. 23 Briefly, central retinal sections (2 mm diameter) were obtained from perfused eyes in neurobasal medium and dark-adapted for 15 min. Halved sections were positioned on an OcuScience ex vivo sample holder, ensuring the photoreceptor layer was upward. Impedance (≤20KOhms), temperature (37°C), and offset voltage (≤10 mV) were adjusted. Samples were perfused with Ames’ Medium at 50 mL/h, and full-field flash ERG recording was performed using a handheld multi-species ERG unit by stimulating the retina with sequential intensities of flashlights. The b/a ratio and b-wave implicit time were evaluated for assessing inner to outer retinal activity at 1000 mcd flash intensity. One pair of eyes in each set was excluded from analysis owing to the absence of a flash response.
Experimental design and statistical analysis
GraphPad Prism was used for comprehensive result analysis. NRN1 and RGC markers expression in glaucomatous and non-glaucomatous retinas were assessed by two-way ANOVA. Pre-perfusion gene expression significance and changes in IOP over 24 h for seven days within each group were evaluated using two-way ANOVA. Extracellular matrix (ECM) marker expression and ERG data were assessed via Student’s t-tests. Post-perfusion retinas underwent two-way ANOVA for CASP3, CASP7, and GFAP, and Student’s t-tests for RBPMS and THY1. Axon counts were analyzed via two-way ANOVA. A significance threshold of p < 0.05 was applied to all experiments. Data are presented as mean ± SEM.
Results
Demographics of study subjects
Detailed demographics of glaucomatous and non-glaucomatous donors are provided in Supplementary Tables S1 and S2. All donors were Caucasians. No significant age differences were found between glaucomatous and non-glaucomatous donors in the NRN1 gene expression study (age: glaucoma, 77.4 ± 3.32; non-glaucoma, 82 ± 2.34; n = 5; p = 0.51; nonsignificant) (Supplementary Table S1). Similarly, no significant age differences were observed between donors used in perfusion studies (age: normal IOP, 85.25 ± 5.54; high IOP, 85 ± 3.89; n = 4; p = 0.57; nonsignificant) (Supplementary Table S2). Medical history assessment revealed no ocular history associated with any non-glaucomatous donor. Among glaucomatous donors, three received IOP lowering drops, whereas others had no documented ocular drug history. For perfusion studies, an equal number of male and female non-glaucomatous donors were used.
Decreased expression of NRN1 and RGCs in glaucomatous retinas
To examine NRN1’s correlation with RGC damage, we assessed NRN1, RBPMS, and THY-1 expression in glaucomatous retinas versus non-glaucomatous retinas (Fig. 1A). Gene expression identified significant reduction of NRN1 in glaucoma retinas as compared with non-glaucomatous retinas. (FC: non-glaucoma, 1.8 ± 0.8; glaucoma, 0.33 ± 0.15; n = 5; **p = 0.007) (Fig. 1B). Similarly, significant decrease in RGC marker RBPMS (FC: non-glaucoma, 1.23 ± 0.55; glaucoma, 0.12 ± 0.05; n = 5; *p = 0.04) together with decreased trend of THY1 (FC: non-glaucoma, 1.14 ± 0.5; glaucoma, 0.25 ± 0.11; n = 5; p = 0.14; nonsignificant) (Fig. 1B) were noted in glaucomatous retinas. To investigate NRN1’s source within the retina and its association with RGCs, we conducted immunostaining on retinal flatmounts using antibodies against NRN1 and RBPMS (an RGC marker). This showed RGCs as the primary source of retinal NRN1, evidenced by the colocalization with RBPMS. However, consistent with our gene expression data, NRN1 protein level was notably lower in glaucomatous RGCs than non-glaucomatous RGCs. Remarkably, substantial RBPMS positive cells in glaucomatous retinas lacked detectable NRN1 staining (Fig. 1C).
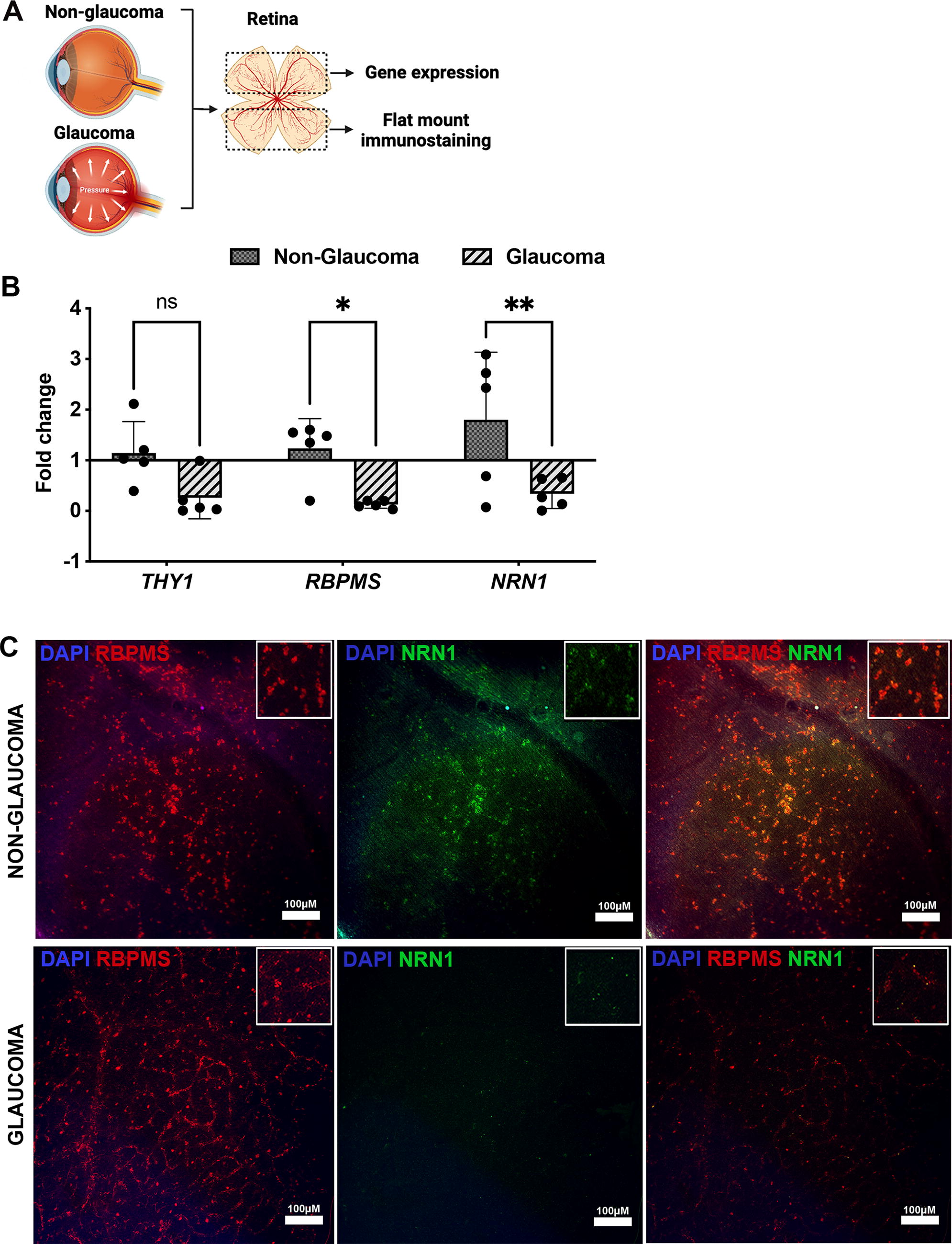
Expression of retinal ganglion cell markers and neuritin 1 in postmortem non-glaucoma and glaucoma retinas.
The TAS model successfully maintained normal and high IOP perfusion
To evaluate NRN1’s neuroprotective efficacy, non-glaucomatous eyes were perfused with or without recombinant hNRN1 in the TAS model under normal and high IOP conditions (Fig. 2A). Paired control eyes allowed normalization of basal apoptosis levels due to axotomy and existing degeneration. We evaluated the retinal health upon receiving eyes using gene expression via TaqMan arrays. This revealed measurable levels of inflammatory (TLR4, GFAP, GSS, and AIF1), apoptotic (BAX, CASP3, and CASP7), and retinal genes (RBPMS, ISL-1, THY1, NEFH, RHO, and PRKCA) at pre-perfusion (Supplementary Fig. S1) with no significant difference in any tested genes between the OD and OS eyes. After confirming pre-perfusion retinal viability, eyes were perfused in TAS model either at normal or high IOP based on our experimental paradigm. During perfusion, OS eyes received 200 ng/mL of hNRN1 in complete neurobasal media, whereas OD eyes served as the vehicle group and received perfusion media without hNRN1. The TAS effectively maintained the donor eyes for 7 days under both normal IOP (vehicle: 17.9 mm Hg ± 3.9 mm Hg, hNRN1: 14.5 mm Hg ± 1.3 mm Hg, n = 4, p > 0.05, nonsignificant) (Fig. 2B) and high IOP conditions (vehicle: 28.2 mm Hg ± 5.36 mm Hg, hNRN1: 30.1 mm Hg ± 3.35 mm Hg, n = 4, p > 0.05, nonsignificant) (Fig. 2C). No significant pressure differences were noted between paired donor eyes within each group under normal or high IOP (Fig. 2B and C).
Maintenance of perfusion pressures with donor eyes in an ex vivo TAS model.
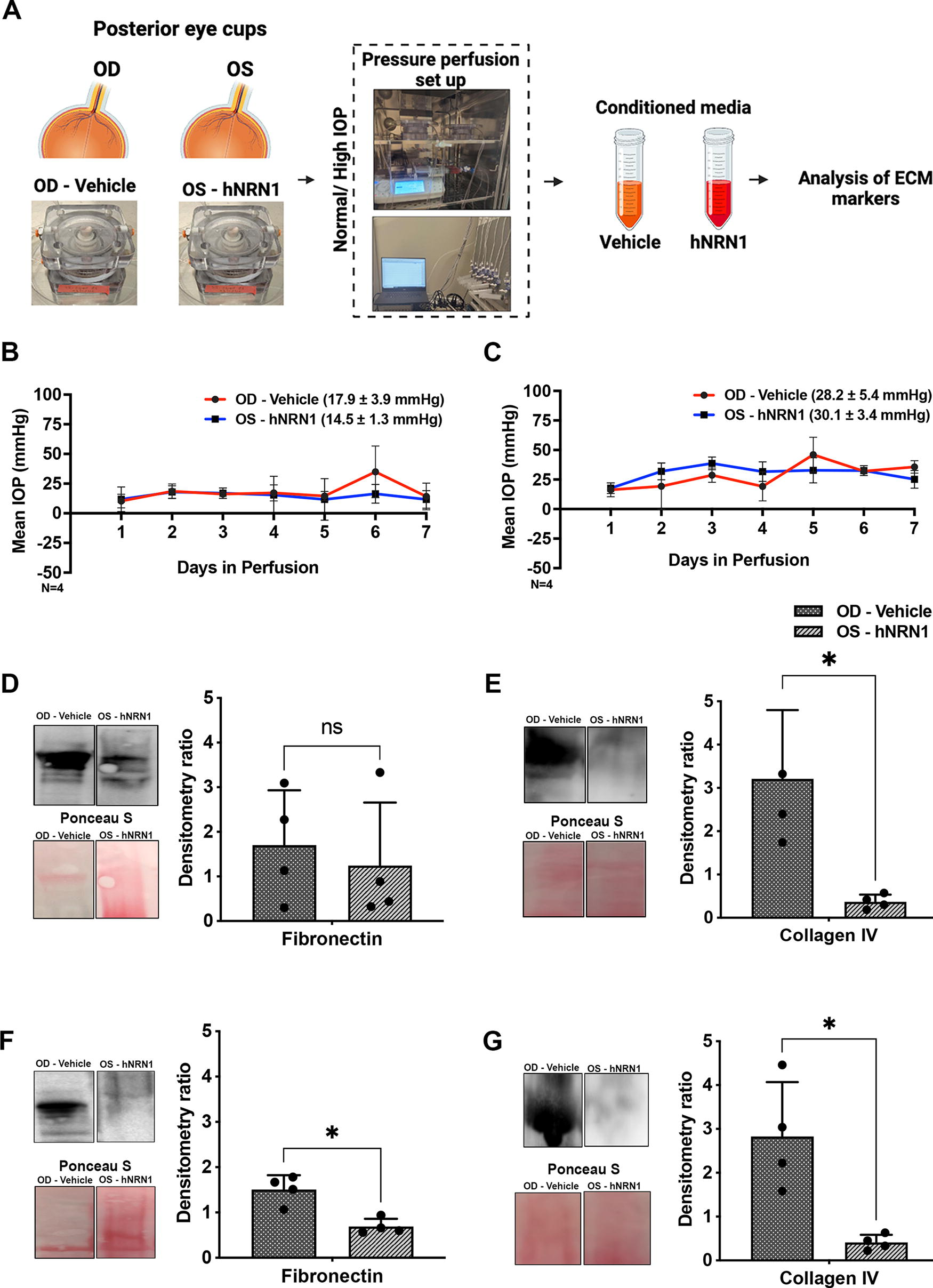
NRN1 treatment reduced the extensive deposition of ECM proteins
Western blotting using conditioned media detected downregulation of fibronectin (n = 4, p = 0.74, nonsignificant) (Fig. 2D) and significant downregulation of collagen IV (n = 4, *p = 0.04) (Fig. 2E) in hNRN1-treated retinas under normal IOP. Likewise, significant downregulation of fibronectin n = 4, *p = 0.04) (Fig. 2F) and collagen IV (n = 4, *p = 0.04) (Fig. 2G) was observed in hNRN1-treated eyes at high IOP than in vehicle-treated eyes. Detailed intensity ratios for each marker are provided in Supplementary Tables S3A and S3B.
NRN1 treatment decreased gliosis and apoptosis
Retinal gene expression analysis of GFAP, CASP3, CASP7, RBPMS, and THY1 was performed in pressure perfused eyes (Fig. 3A). Under normal IOP, hNRN1-treated eyes exhibited decreased GFAP expression (n = 4, *p = 0.01) with no significant difference in apoptotic markers CASP3 (n = 4, p = 0.9, nonsignificant) and CASP7 (n = 4, p = 0.9, nonsignificant) compared with vehicle-treated eyes (Fig. 3B). However, a trend toward increased expression of RGC markers RBPMS (n = 4, p = 0.32, nonsignificant) and THY1 (n = 4, p = 0.64, nonsignificant) in hNRN1-treated eyes under normal IOP conditions were noted (Fig. 3C, 3D). In contrast, under high IOP, hNRN1-treated eyes exhibited significant decrease in GFAP expression (n = 4, ***p = 0.0005) and in the apoptotic markers CASP3 (n = 4, *p = 0.01) and CASP7 (n = 4, **p = 0.004) compared with vehicle-treated eyes (Fig. 3E). Furthermore, a significant increase in RBPMS expression (n = 4, *p = 0.01) (Fig. 3F) and a trend toward increased THY1 expression (n = 4, p = 0.64, nonsignificant) in hNRN1-treated eyes under high IOP were noted (Fig. 3G). Detailed FC and absolute values for each tested gene are provided in Supplementary Tables S4A-D.
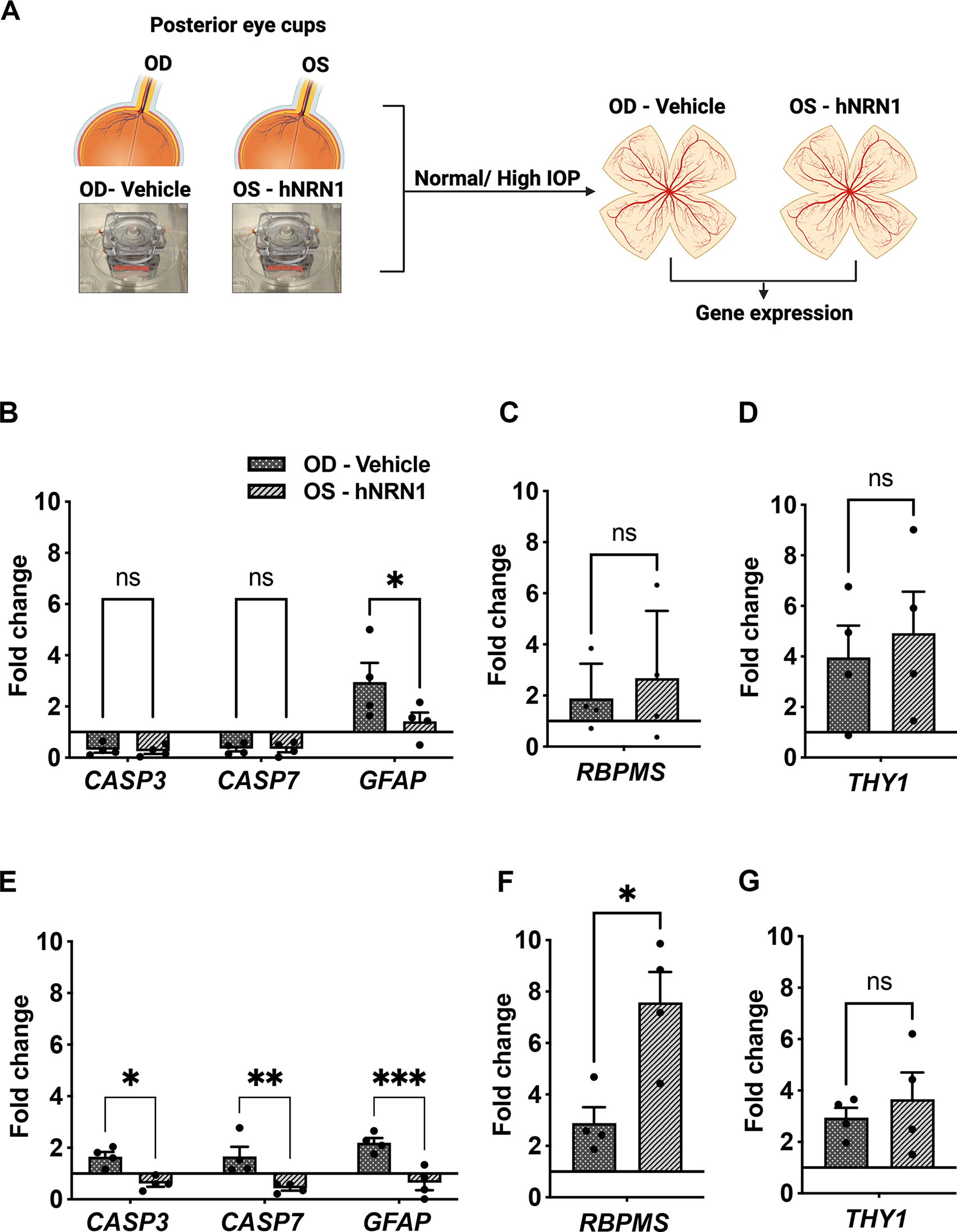
Quantitative gene expression of hNRN1-treated eyes in comparison with vehicle-treated eyes.
NRN1 treatment reduced axonal degeneration
To evaluate the effects of hNRN1 on RGC axons, 1 cm-long ON segments were dissected from the ON head after perfusion. The distal ON segments underwent PPD staining to assess axon health (Fig. 4A) and the percentage of normal and degenerated axons in each condition were calculated. In normal IOP group, a comparable number of normal and degenerated axons were seen between vehicle (Fig. 4B) and hNRN1 treatment (Fig. 4C). In Figure 4B, C, E, and F, asterisks indicate healthy axons and arrowheads indicate degenerated axons. The axon counting further confirmed it (Fig. 4D) (normal axons: vehicle, 64.12 ± 3.7%; hNRN1, 60.6 ± 5.7%; n = 3; p > 0.05; nonsignificant; degenerated axons: vehicle, 35.8 ± 3.7%; hNRN1, 39.3 ± 5.7%; n = 3; p > 0.05; nonsignificant). Furthermore, axon degeneration was notably exacerbated in high IOP eyes, confirming high IOP accelerates axon degeneration compared with normal IOP (Fig. 4E). However, hNRN1 treatment at high IOP reduced the number of degenerated axons (Fig. 4F). The axon counts showed a significant increase in normal (vehicle: 62.7 ± 0.86%, hNRN1: 66.4 ± 0.43%, n = 4, **p < 0.004) and decrease in degenerated axons (vehicle: 37.2 ± 0.87%, hNRN1: 33.5 ± 0.43%, n = 4, **p < 0.004) in hNRN1-treated eyes compared with vehicle-treated eyes at high IOP. These collective findings provide evidence for the potential of hNRN1 in preserving RGC axonal integrity especially at high IOP.

Paraphenylenediamine staining revealed reduced axonal degeneration upon hNRN1 treatment.
NRN1 treatment enhances retinal activity
Ex vivo ERG evaluated retinal function in pressurized eyes (Fig. 5A). We measured scotopic b/a ratio and b-wave implicit time, as a measure of inner to outer retinal responses. The hNRN1 administration under normal IOP showed no significant change in b/a ratio (vehicle: 6.2 ± 5.2 µV, hNRN1: 5.82 ± 3.1 µV, Fig. 5B, n = 3, p = 0.94, nonsignificant) but slight decrease in b-wave implicit time (vehicle: 21.4 ± 0.89 mS, hNRN1: 20.7 ± 0.43 mS, Fig. 5C, n = 3, p = 0.17, nonsignificant). However, hNRN1-treated eyes at high IOP exhibited a significant increase in b/a ratio (vehicle: 11.7 ± 6.23 µV, hNRN1: 16.16 ± 6.28 µV, Fig. 5D, n = 3, ***p = 0.001), with a shortened implicit time for b-waves (vehicle: 20.9 ± 0.92 mS, hNRN1: 19.9 ± 0.05 mS, Fig. 5E, n = 3, p = 0.34, nonsignificant), suggesting retinal functional restoration with hNRN1 at high IOP.
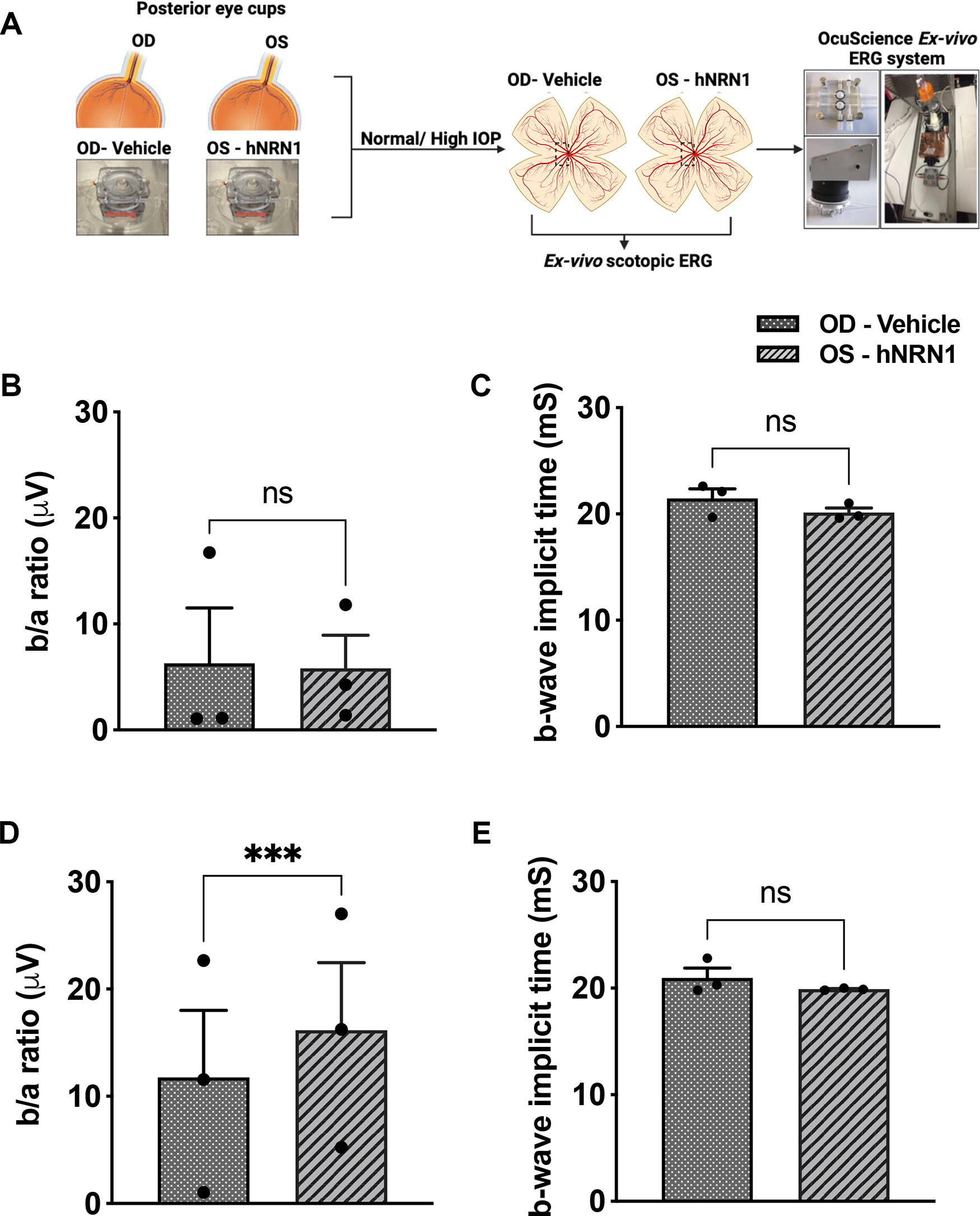
Measurement of retinal activity in hNRN1-treated eyes using ex vivo ERG.
Discussion
Elevated IOP-induced optic neuropathy is a major global cause of vision loss. Even with well-controlled IOP, individuals may suffer significant vision loss due to progressive RGC degeneration. 27 Hence, identifying a molecular target to protect and regenerate RGCs is crucial in tackling glaucoma. Prior studies showed protective effects of NTFs like BDNF and CNTF on RGCs. However, their practical limitations including BDNF’s affinity for ipRGCs 28 and CNTF’s context-dependent effectiveness, particularly in ON injury models. 29 In contrast, NRN1 stands out as a promising alternative. The NRN1, a downstream effector molecule in the neurotrophin family, has nine binding sites for transcription factors responsive to neural activity.17,30 It offers neuronal protection by activating mitogen-activated protein kinase or phosphatidyl insositol-3 kinase pathways. 31 With significant neuroprotective and neuroregenerative attributes of NRN1 as evidenced at a dosage of 200 ng in both the in vivo and in vitro studies of glaucoma,19,21 exploring the therapeutic potential of NRN1 in human models of glaucoma holds significant promise. Activating NRN1 could precisely protect RGCs by triggering multiple downstream pathways of neuroprotection, 21 regeneration, 32 and chemotaxis. 22
Despite prior in vitro and in vivo evidence of NRN1’s therapeutic potential,19–21 its efficacy in human RGCs or glaucoma remains unexplored. Prior research demonstrated NRN1 mRNA expression in purified adult rat RGCs, 33 and our current study confirmed NRN1 expression in human RGCs. Neurotrophin deprivation and axonal damage are hallmarks of glaucoma. 34 Given that NRN1 is a downstream effector of multiple NTFs,17,30 it is highly plausible that reductions in NTF levels, such as BDNF, NT-3, or NGF could lead to decreased activation of the NRN1 mRNA and subsequently contribute to the accelerated RGC loss. This could explain the observed reduction in NRN1 levels and RGC numbers in the glaucomatous retinas (Fig. 1). One limitation of our study is the lack of detailed glaucoma history. The donor eyes were categorized as glaucoma and non-glaucoma through optic cup evaluations and medical and family investigations conducted by the eye bank. However, eye banks’ electronic medical records do not contain information regarding glaucoma types or the last IOP record. Consequently, we were unable to specify the glaucoma types used in this study. These constraints have been taken into careful consideration throughout our analysis and results interpretations. However, evaluating the association of NRN1 with various types of glaucoma is essential and will be a future extension of this study.
To assess hNRN1’s effect on RGCs in a human glaucoma-like microenvironment, we used an established ex vivo glaucoma model, the TAS.23,24 This system allowed us to perfuse human posterior eye cups and serve as a human preclinical pressurized glaucoma model. Remodeling of ECM in the lamina cribrosa is well-documented in glaucoma 35 and strongly associated with astrogliosis and RGC death.36,37 Gliosis enhances ECM remodeling by increasing both metalloproteinases activity38,39 and neuroinflammatory cytokines secretions in an altered retinal microenvironment.40,41 Targeting gliosis and ECM remodeling will have a beneficial impact on RGC survival. Notably, NRN1 has been documented to mitigate astrogliosis and ameliorate cognitive dysfunctions in diabetic individuals. 42 Our gene expression data align with these findings, as we observed a significant decrease in gliosis following hNRN1 treatment in eyes subjected to both normal and high IOP (Fig. 3B and E). However, hNRN1 protective effects were higher in high IOP eyes than in normal IOP eyes. This could be attributed to two potential reasons. Firstly, normal IOP, irrespective of hNRN1 treatment, caused minor homeostatic disruptions, mainly activating astrocytes. Thus, hNRN1 treatment could only reverse the astrogliosis. The studies, including ECM evaluation, apoptotic gene expression, axon counts, and retinal activity, have substantiated this conclusion. Secondly, it is plausible that NRN1 is activated primarily in challenging conditions like high IOP, which needs to be investigated further.
Gliosis enhances ON degeneration in glaucoma43,44 with elevated IOP being a major risk factor associate with damage. 45 Prior studies demonstrated that NRN1 enhances axonal transport and promotes ON regeneration in axotomized rodent ONs.19,20 We used PPD staining to evaluate the distal portion of ON after perfusion. The percentages of normal and degenerated axons were comparable in normal IOP eyes regardless of hNRN1 treatment, indicating minimal homeostatic disruption and a lesser need for hNRN1’s therapeutic effect. However, in high IOP, hNRN1 provided greater axonal protection evidenced by significant reduction in degenerated axons with hNRN1 treatment (Fig. 4). Thus, our findings align closely with prior studies where NRN1 was shown to reduce neuronal death with increased synaptic connectivity, 46 revealing reduced axonal degeneration in the high IOP eyes treated with hNRN1.
To evaluate hNRN1’s effect on retinal activity, we performed ex vivo ERG and assessed retinal activity by calculating the b/a ratios and b-wave implicit time. Previous reports have identified that b/a ratio and b-wave implicit time are sensitive analyses for evaluating treatment outcomes in retinal pathologies associated with inner retinal dysfunctions.47–49 Interestingly, eyes at normal IOP, with or without hNRN1 depicted a slight decrease in b/a ratios with reduced b-wave implicit time; suggesting that hNRN1 does not enhance retinal activity under normal conditions. Under high IOP, the ERG showed significant increase in b/a responses with less implicit time, suggesting hNRN1 treatment restored inner retinal function under high IOP, which strongly correlates with our ON degeneration data.
In conclusion, our study strongly supports the potential therapeutic efficacy of hNRN1 in glaucoma. Nonetheless, there are several facets of this research that require further validation. One approach could involve extending hNRN1 treatment duration or dosing under normal IOP to explore its effects in conditions like normal tension glaucoma. This could lead to more significant functional improvements under normal IOP, offering promising neurotherapeutic options for individuals presenting with normal IOP but significant RGC loss. In addition, testing hNRN1 as a cotherapy with established IOP-lowering treatments holds promise for enhancing protective effects in individuals with glaucoma.
Footnotes
Acknowledgements
The authors thank the UW Madison Electron Microscope Facility and Randall J. Massey for PPD staining, processing of the optic nerve head sections and electron microscopy. The authors also thank the donors and families of human donors for providing their tissues for research. The authors acknowledge Biorender for generating the schematic images presented in the article.
Authors’ Contributions
S.S.H. performed design, investigation, data acquisition, validation, and writing original draft of the article. S.S.H. together with N.E.B. and R.C.M. was responsible for formal analysis of the data and reviewing the article. T.P.S. was responsible for the conceptualization, project design, funding acquisition, project administration, supervision, reviewing, and approving the final version of the article.
Availability of Data and Materials
All the data generated and analyzed during this study are included in this published article and its supplementary information files.
Author Disclosure Statement
The corresponding author declares the conflicts of interest on the TAS model, which received U.S. Patent US20190327958A1.
Funding Information
This work was supported by a grant received from
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.