
Abstract
Abstract
Background:
The lymphatic system plays a critical role in the body's fluid and protein homeostasis, immune regulation, and dietary fat absorption. One of the major pathologies of the lymphatic system is primary lymphedema, which occurs in approximately 0.6% of live births and is caused by missing or impaired lymphatic vessels. Although there is a great need for medical intervention into diseases of the lymphatic system, very little is known about its development or how it maintains integrity over time. Recent studies have suggested that biophysical components, such as local extracellular fluid flow, may be important factors during initiation of lymphangiogenesis. We hypothesize that interstitial fluid flow functions as an important morphoregulator during developmental lymphangiogenesis.
Methods and Results:
In the present study we use pharmacological agents and a mutant fish line to modulate interstitial flow. Our data confirm that a sufficient increase or decrease in interstitial flow can profoundly affect lymphatic patterning and may result in a lymphedema-like phenotype. Proper interstitial flow appears to be necessary during LEC migration for proper lymphatic development.
Conclusions:
These results support the contention that interstitial flow is an important morphoregulator of developmental lymphangiogenesis.
Introduction
Our current understanding of the events characterizing developmental lymphangiogenesis comes largely from murine studies (reviewed in Refs. 8 and 9). This body of research has shown that lymphatic development initiates at embryonic day (E) 9–9.5 and is marked by cells in the anterior region of the cardinal vein expressing LYVE-1 (lymphatic vessel hyaluronan receptor 1).10,11 These LYVE-1-expressing cells are the lymphatic endothelial cell (LEC) progenitors that receive signals guiding their migration from a venous origin to a site where the primitive lymphatic sac forms.8,11–13 The primitive lymphatics transition to a phase of vascular coalescence, maturation, and differentiation into the lymphatic capillaries and collecting vessels. 9 A number of genetic factors are thought to play regulatory roles in developmental lymphangiogenesis (reviewed in Ref. 8); however, in vivo studies exploring important flow-modulated regulation resulting from changes in the local microenvironment remain unexplored. Intravital fluid flow (interstitial and intravascular) is known to impose substantial mechanical stresses (e.g., shear, stretch, transmural pressure) on adjacent and underlying cells, as well as facilitate convective transport of heat and a myriad of biomolecules. These biofluid flows have been well established as key morphoregulators in a number of developmental processes including symmetry determination,14,15 cardiogenesis,16–18 blood vessel formation,19,20 glomerulogenesis, 21 brain development, 22 and lung development.23,24 In addition, aberrant flows have been shown to be important factors in pathogenesis of a variety of diseases of the cardiovascular,25–27 nervous,28–30 and renal31–33 systems as well.
Interstitial flow is fluid moving through the three-dimensional extracellular matrix comprised of fibroblasts, tumor cells, tissue immune cells, and adipocytes. 34 These intravital flows are an important component of normal tissue function and homeostasis, serving as the conduit for fluid and biomolecule delivery to and from metabolizing tissues. Interstitial flow is present in nearly all tissues and its multidirectional motions around the cell–matrix interface provide an array of cues impacting tissue morphogenesis and function, cell migration and differentiation, and matrix remodeling. 35 Data currently available on the lymphangiogenetic process suggest that interstitial fluid flow may stimulate lymphatic channel formation, expression of primary lymphangiogenic genes, and vessel formation by LECs.35,36 It has been posited that functional lymphatic growth may be dependent upon preexisting interstitial flow.37–39 Unfortunately, most of these studies have been performed in vitro and, therefore, suffer from the inability to reproduce the complexity of biological flows and the physiochemical interconnectivity of adjacent tissues seen within a developing embryo.34,40,41
The flow of any fluid is the result of a pressure drop along its net path of travel. The driving force for interstitial flow is the pressure difference between the blood vasculature and the initial lymphatics. 1 Plasma may exit blood vessels via paracellular pathways if blood pressure sufficiently exceeds local colloid osmotic pressure. In the fields of fluid dynamics and hydrology, Darcy's law is used to describe the flow of a fluid through a porous medium. It is predicated on the proportional relationship between the instantaneous rate of fluid discharge, fluid viscosity, and the distance over which a measurable pressure drop occurs. Darcy's law has been used in biological applications to describe intravital fluid velocity moving through a 3D extracellular matrix as the pressure difference (Pblood – Plymphatic) divided by the intervessel distance and the interstitial hydraulic conductivity (which varies depending on the density and composition of each tissue). 34 If we assume that tissue resistance is constant, Darcy's law predicts an increase in bulk interstitial flow by raising blood pressure.
The role of interstitial flow on secondary lymphangiogenesis has been addressed with mouse tail assays; 39 however, this method cannot be used to study the role of developmental lymphangiogenesis due to murine in utero development. In order to determine whether changes in the dynamic, intravital flow milieu can affect the developing lymphatic vasculature, a model system that is optically accessible during lymphatic development and amendable to pharmacological and genetic manipulation is needed.
The zebrafish (Danio rerio) has recently been identified as a model for studying developmental lymphangiogenesis due to its optical clarity, genetic plasticity, small size, and rapid development (the primary lymphatic vessel, the thoracic duct [TD], forms by 4 days post-fertilization [dpf]).42–44 Pharmacological manipulation of blood pressure is readily accomplished by adding agents directly into the embryonic medium, resulting in measureable changes in heart rate, vessel diameter, stroke volume, and blood flow. 45
Our study utilizes the embryonic and larval zebrafish to determine whether interstitial fluid flow plays a morphogenic role during developmental lymphangiogenesis. Taking advantage of the strengths of the fish model, we manipulated interstitial flow by modulating blood pressure both pharmacologically and genetically to determine its effect on the extent and timing of lymphatic development. We determined that both elevated and depressed interstitial flow resulted in significant dysmorphogenesis of the lymphatic vasculature and that these events appear to take place during a specific developmental window (2–3 dpf). As a result, we conclude that the biophysical forces produced by flow within the interstitium are unlikely to be exerting a shear stress effect on the LECs, but rather, the flow is likely important in establishing a biochemical gradient of some factor influencing the migration of LECs (e.g., VEGF-C). Finally, we propose a developmental molecular model that attempts to explain the observed dysmorphogenesis in cases of both excessive and insufficient interstitial flow.
Materials and Methods
Zebrafish lines and maintenance
In the UC-CCHMC zebrafish colony, embryos were obtained from breeding adult zebrafish. Adult fish were maintained with 14:10 light:dark cycles at 28.5°C and fed a mixed diet of Aquatox flake food (Aquatic Ecosystems, Inc.) and live Artemia. Care of young embryos, as well as raising, maintaining, and spawning of adult zebrafish, was performed as previously described. 46 Water chemistry was maintained at conductivity 490–530 μS, temperature 26.5–28.5°C, pH 7.1–7.4, and dissolved oxygen of 5.0–7.5 mg L−1. All experimental manipulations and animal husbandry were performed in accordance with both CCHMC and UC IACUC guidelines. The Tg(kdr-l:HsHRAS-mCherry)s916xfli1:GFP (from here on referred to as kdr-l:RasCherryxfli1:GFP) double transgenics were used in these experiments to optically determine if the lymphatic vasculature formation was altered with various flow modulation treatments. The kdr-l:RasCherryxfli1:GFP line express green fluorescence in only the lymphatic vessels, while blood vessels are marked with both red and green fluorescence 47 (with the exception of the GI vasculature), 42 allowing easy optical analysis of TD formation.42,47,48
Confocal microscopy
Embryos were mounted on their sides in glass-bottom Petri dishes into 1.2% agarose containing 125–150 mg L−1 MS-222. 18 Imaging of the embryonic vasculature was performed using a Ziess LSM 510 scanning confocal scanner and associated Axiovert 100M inverted microscope. Sample illumination was provided by 488 nm and 543 nm solid-state lasers as appropriate for the GFP (488 nm), YFP (513 nm), and RasCherry (587 nm) fluorophores. The light path was configured with a HFT 488/543 excitation filter, a bandpass 500–550 nm IR filter and a longpass 560 nm filter to establish the individual detection channels. A Plan-Apochromat 20X/0.6 objective (zoom 0.7) was used throughout. Z-stacks of each embryo were taken using an approximately 4 micron Z-plane intervals to create three-dimensional mid-sagittal renderings of each fish in the images shown, unless otherwise noted. Maximum intensity projection (MIP) images were prepared in Zen 2009 Light Edition (Carl Zeiss Microimaging GmbH, Germany).
Angiography
3dpf AB Wild Type embryos were injected with Alexa Fluor 568 labeled 10,000 molecular weight dextran (Invitrogen). Preliminary studies revealed that this size of dextran is small enough to pass through the walls of embryonic arteries and into the interstitium, but it is not filtered through the kidney. Embedded injected embryos had an overlay of embryo medium containing the same concentration of agent corresponding to their treatment group. Microinjections were performed using a PLI-100 Pico Injector (Harvard Apparatus, Hollister, MA) with borosilicate glass pipettes drawn to a tip diameter of 2–3 μm using a P-2000 horizontal laser puller (Sutter Instruments, Novato CA). Multi-bolus injections were made into the common cardinal vein or posterior cardinal vein using injection pressures of 2–5 psi. Injected dextran was allowed to circulate approximately 10 min before the first confocal image was taken. The rate of dextran passage from the blood vasculature into the interstitium was optically quantified by monitoring the loss of pixel intensity associated with dextran in a segment of the dorsal aorta (DA) over time. Maximum pixel intensity was calculated using Volocity image analysis software (PerkinElmer, Waltham, MA) for this analysis. The intensity difference between time points was divided by the initial intensity, to standardize and determine the percentage decrease over the experimental time frame. This approach allowed us to compare data between individual fish despite variations in initial fluorescence intensity due to subtle changes in dextran injection volume.
Blood pressure modulation
Reduction of interstitial flow was accomplished by decreasing blood pressure through exposure to 8 mM 2,3-butanedione monoxime (2,3-BDM) (Sigma-Aldrich, St. Louis, MO), an inhibitor of myofibrillar ATPase activity. 49 2,3-BDM has been shown to be effective in reducing embryonic zebrafish heart rate in a dose-dependent fashion, resulting in diminished blood flow. 50 A 2,3-BDM stock was prepared at 0.1 g ml−1 in methanol. This stock was diluted to 8 mM 2,3-BDM, such that experimental treatments contained no more than 0.8% methanol. Interstitial flow was pharmacologically increased through introduction of epinephrine (Sigma) (a β-adrenergic agonist that increases blood pressure) into the embryo medium. It has been shown that developing zebrafish have adrb1, a beta adrenergic ortholog, expressed in both the heart and brain, 51 which is important for epinephrine signaling herein. A 50 mg ml−1 stock was prepared in hydrochloric acid and diluted to 150 μM such that experimental treatments contained no more than 0.005% hydrochloric acid. As an additional way to increase interstitial flow, phenylephrine (Sigma) was used. Although phenylephrine is traditionally a specific alpha-adrenergic agent, it has been shown in zebrafish to also bind the beta2a-adrenergic receptor, 52 but is still more specific than epinephrine, which should bind to all of the adrenergic receptors. Phenylephrine was dissolved in water to the concentration of 1 mM. Embryos were treated with 1 mM phenylephrine from 2–4 dpf.
Results
Alteration of interstitial fluid flow
The hypothesis that interstitial flow serves as an important morphoregulator of developmental lymphangiogenesis was tested by altering zebrafish blood pressure as a means of changing interstitial flow. This approach was based on the predictions of Darcy's law which suggest that increased blood pressure should provide the necessary driving force for increasing bulk interstitial flow by forcing blood plasma, through paracellular pathways, into the interstitium. Applying this logic, epinephrine or 2,3-BDM were used to increase or decrease interstitial flow, respectively (as described in Materials and Methods).
To determine if these pharmacological agents were altering flow as would be predicted, AB wild-type zebrafish embryos (3 dpf) were exposed to either 150 μM epinephrine, 8 mM 2,3-BDM or control (embryo water with vehicle) for 1 h. Angiography using 10,000 MW dextran as the contrasting agent was performed and confocal images acquired at 10 min post-injection (time 1) and then again 35 min later (time 2) ) (Fig. 1A, single plane image shown).
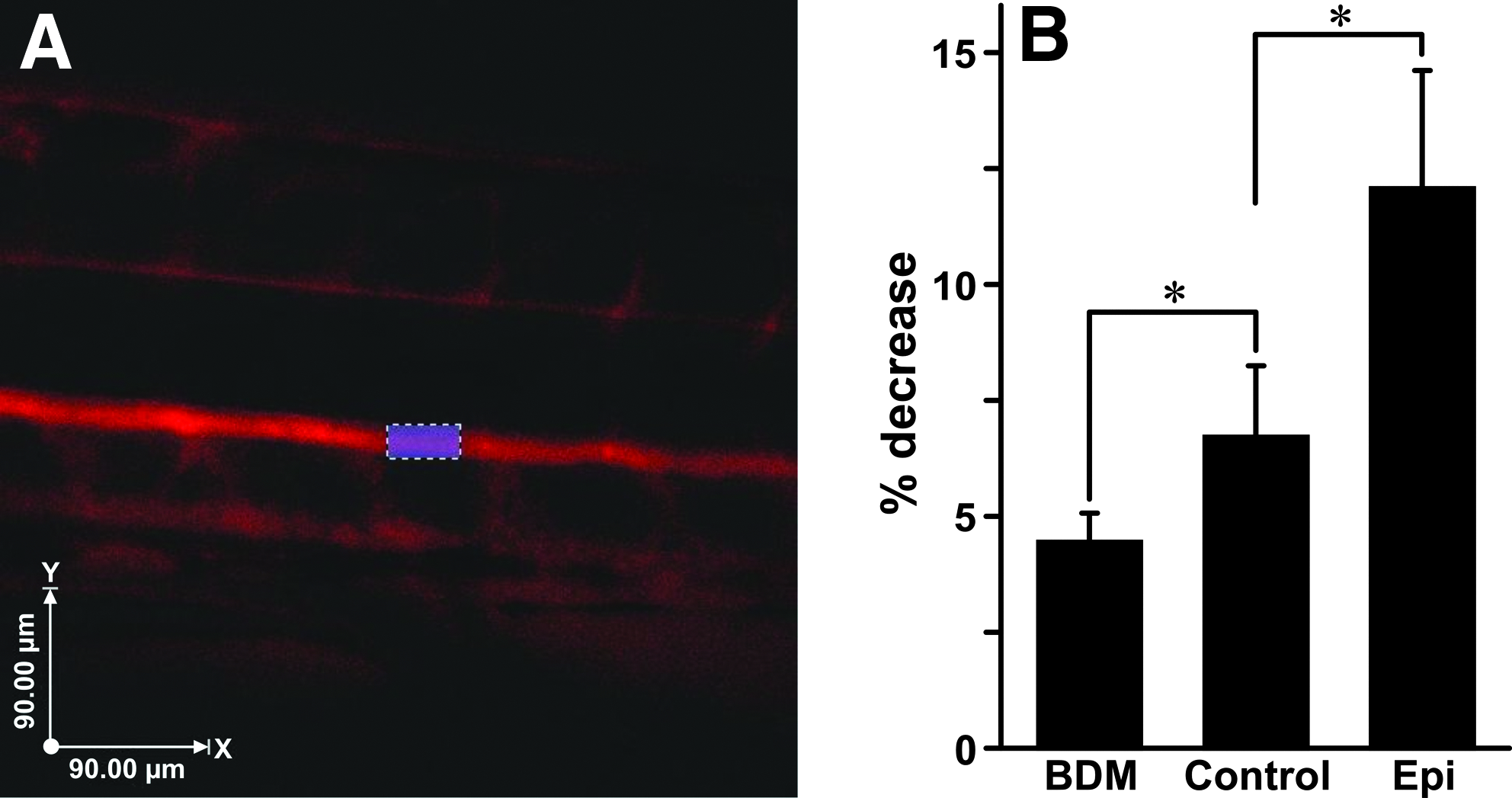
Dextran angiography injections show the predicted drug induced changes in flow patterns.
It was expected that the 2,3-BDM treatment would have the smallest percentage decrease in dextran intensity and that the epinephrine-treated embryos would have the greatest percentage decrease, as the blood pressure should be the highest in these embryos and, therefore, the greatest percentage of dextran should be expelled over the experimental time frame. Embryos treated with 2,3-BDM showed a 4.4% decrease in aortic fluorescence intensity over the 35 min experimental window due to transport into the interstitium. Embryos with elevated blood pressure due to epinephrine exposure demonstrated more rapid movement of dextran out of the blood vasculature (12.06% decrease). Control nontreated embryos that exhibited normal developmental flow showed a more moderate decrease of 6.71% (Fig. 1B). These data support our expectation that experimentally induced changes in blood pressure within zebrafish embryos result in corresponding changes in interstitial fluid flow.
Pharmacological manipulation of interstitial flow
After confirming that changes in blood pressure resulted in commensurate changes in interstitial flow, we proceeded to test whether those changes altered the extent and timing of lymph vessel formation. Zebrafish LECs begin their migration from the parachordal lymphangioblast (PL) string to form the primary lymphatic vessel, the TD, at approximately 2 dpf.44,48 Over the next 48 h, these initial “islands” of progenitor cells coalesce to form a complete vessel by 4 dpf. Therefore, we treated the kdr-l:RasCherryxfli1:GFP double transgenic line for 48 h during this critical migratory event (2–4 dpf) with either 150 μm of epinephrine or 8 mM 2,3-BDM to modulate interstitial flow as before. Our data indicate that any significant alteration (increase or decrease) in interstitial flow resulted in complete loss of TD formation (Fig. 2). Our observation that either an increase or decrease in interstitial flow leads to a lymphatic dysmorphogenesis suggests that our experimental flow profiles are disrupting a biochemical gradient necessary for cell migration rather than providing a direct biomechanical stimulus. This hypothesis is supported by the expectation that a flow-induced stress (e.g., shear) applied to the system would result in an altered phenotype in the case of either an increase or decrease in the magnitude of the stressor, but not both.

Alterations (increase or decrease) in interstitial flow cause a loss of normal TD development. All embryos are from the kdr-l:RasCherryxfli1:GFP line.
We contend that the observed dysmorphogenesis of lymphatic development following alteration of interstitial flow results from either a disruption in migration of LECs from the PL string or their assembly into a functional vessel. To determine which process aberrant interstitial flow was affecting, we altered flow during the two major steps in lymphatic development, LEC migration (2–3 dpf) or coalescence (3–4 dpf). We performed identical pharmacological experiments (utilizing epinephrine and 2,3-BDM) in the kdr-l:RasCherryxfli1:GFP double transgenic line, but limited the exposure of our drug treatments to a 24 h developmental window in which migration or lymphatic vessel coalescence was occurring. We observed a loss of the TD in the 2–3 dpf treatment group closely phenocopying what we observed in the groups treated from 2–4 dpf (Figs. 3B and 3C). The group treated from 3–4 dpf, however, had normal TD formation (Figs. 3D and 3E). These data strongly support the contention that interstitial flow is an important morphoregulator for the proper migration of LECs.

Altering (increasing or decreasing) interstitial flow during LEC migration causes loss of normal TD lymphatic development. All embryos are from the kdr-l:RasCherryxfli1:GFP line.
Additional manipulation of interstitial flow
While our experiments demonstrate that pharmacologically-induced changes in interstitial flow, via altered blood pressure, may result in failed developmental lymphangiogenesis, they assume that neither epinephrine nor 2,3-BDM exert other effects that may impinge upon vascular development. To assuage these concerns, we retested the hypothesis of interstitial flow being a morphogen for lymphatic development in a flow-compromised genetic mutant, silent heart (sih). The sih mutation affects the gene tnnt2, which encodes the thin-filament contractile protein cardiac troponin T. 53 These mutant embryos are identifiable via the visual phenotype of a noncontractile heart, pericardial edema, and complete lack of blood flow. We crossed the sih line with a fli1:GFP transgenic line, which express GFP in all blood and lymphatic vasculature for direct visualization of lymphatic vascular phenotype. Analysis of lymphangiogenesis in these mutants at 4 dpf revealed that sih-/- mutants have a substantial loss of TD development while maintaining a normal looking fluorescent blood vasculature (Fig. 4). These results are concordant with the data from our 2,3-BDM-treated embryos which expressed TD loss under conditions of reduced interstitial flow.
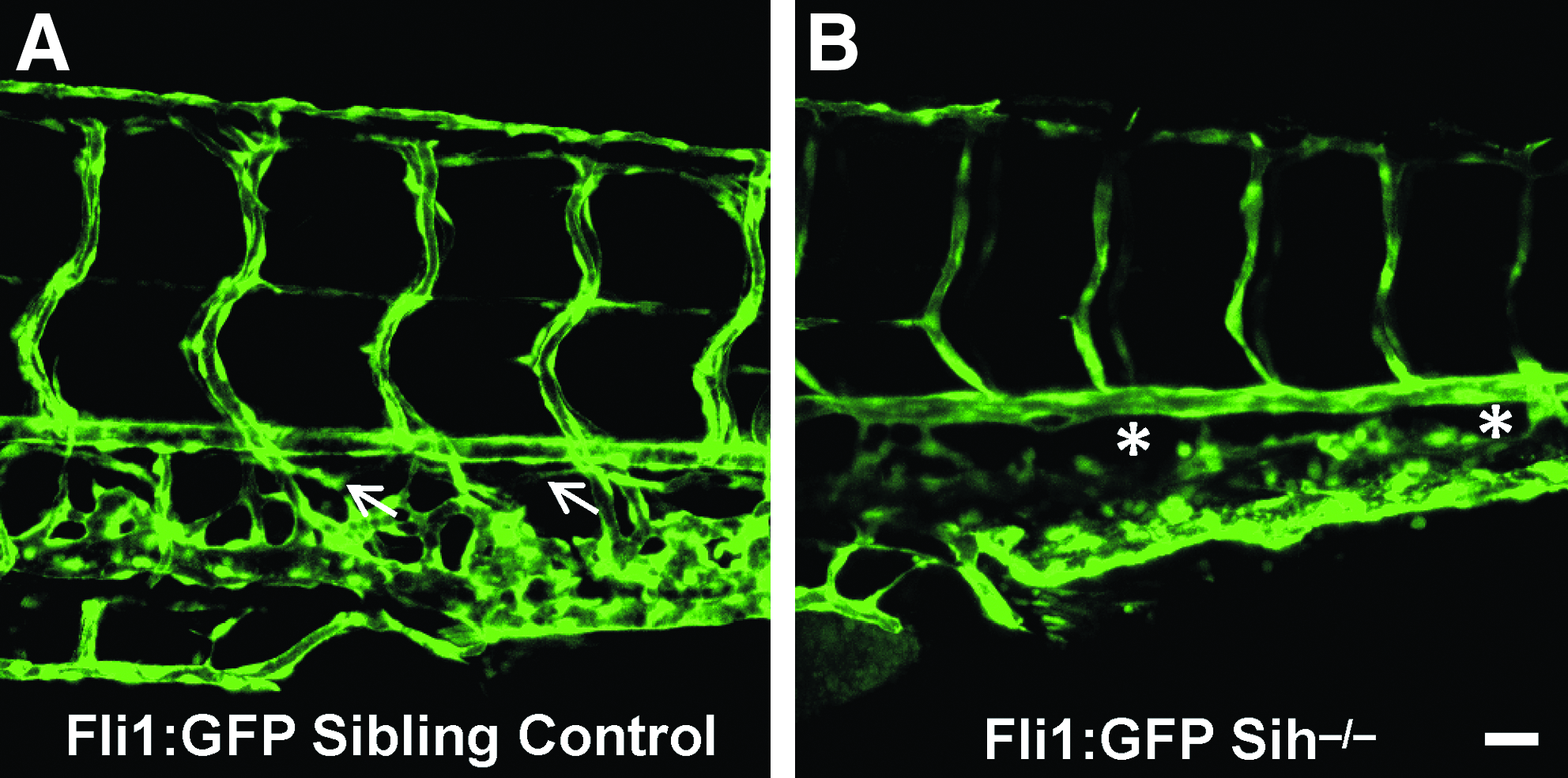
Lymphatic analysis of the genetic flow mutant, sih-/-, shows loss of normal TD lymphatic formation.
We are aware of no existing genetic mutant with an elevated flow to compare with our epinephrine-treated groups, so we used phenylephrine, an alpha-adrenergic agonist as an additional way to increase blood pressure and, therefore, interstitial flow. Phenylephrine has been used previously in zebrafish.52,54 We also did additional testing to ensure that phenylephrine was constricting the DA (the major artery of the zebrafish). The diameter of the DA 10 min after the addition of 1 mM phenylephrine was decreased by an average of 4.67% compared to the pre-treatment diameter. This analysis shows that phenylephrine causes a constriction of the DA, which Darcy's law implies will cause a corresponding rise in blood pressure and interstitial flow. We next tested the effect of phenylephrine on TD formation from 2–4 dpf. We found that at 4 dpf the embryos with this treatment had substantial loss of TD formation (Fig. 5.). This data provide further support for our hypothesis that normal interstitial flow is an important morphoregulator of developmental lymphangiogenesis, as both of these additional ways of decreasing and increasing interstitial flow yielded the same result of obstructing normal developmental lymphangiogenesis.

Lymphatic analysis of the embryos treated with the phenylephrine to increase flow show loss of normal TD lymphatic formation. Embryos are from the kdr-l:RasCherryxfli1:GFP line.
Discussion and Conclusions
The genetic and biophysical regulation of developmental lymphangiogenesis remains elusive despite a renewed focus on lymphatic vascular biology. 55 Although recent studies have begun to shed light on potential developmental mechanisms that lead to lymphedema, a frustrating disease that diminishes quality of life, 56 further research in this area is imperative for substantial progress to be made towards medical cures as the current treatments are only palliative. 57 One area where progress is being made is in the investigation of potential morphoregulators of lymphatic development. One such factor, interstitial fluid flow, is particularly promising in that it continues to be implicated in the regulation of a number of critical developmental processes, including developmental lymphangiogenesis.35,36 Despite the seemingly sluggish flow rates observed, in vitro studies have demonstrated that endothelial cells are capable of sensing and responding to minute changes in flow magnitude and direction,38,58 and response to these stimuli may be greatly enhanced by extracellular matrix components. 34 This is likely due to the fact that while flow only slightly biases the distributions of a secreted matrix-binding morphogen (a molecule that gives cells directional and positional information), flow also affects the proteases that liberate the morphogens. Thus, the flow effects are additive as additional flow-related bias occurs after the morphogen has been liberated. 34
The use of powerful new vertebrate models, such as the zebrafish, greatly enhances our ability to characterize and manipulate the complex fluid flow environment found within living systems. Not surprisingly, use of the zebrafish in studies of developmental lymphangiogenesis is becoming more widespread as researchers recognize the utility of this model system, specifically its small size, optical clarity, genetic tractability, and rapid development. In fact, recent in vivo studies have revealed that LECs begin to migrate from the PL string at 2 dpf 44 and follow the path of intersegmental arteries to first form the thoracic duct, 59 which is complete at approximately 4 dpf.
The goal of the present study was to test the hypothesis that interstitial flow is an important morphoregulator for developmental lymphangiogenesis. Using both pharmaceutical treatments and genetic mutation, we show that either increased or decreased interstitial flow leads to substantial loss of TD formation. These data suggest that normal interstitial flow is critical for proper developmental lymphangiogenesis, particularly during LEC migration (2–3 dpf). In addition, we demonstrate that lymphatic development is not affected by alterations in interstitial flow after LEC migration, during coalescence into the TD. Our results were confirmed in a mutant zebrafish line exhibiting impaired blood flow (sih-/-), where lack of normal TD formation is observed. The mechanism of action by which interstitial flow might elicit its effects include direct biophysical stimulation (e.g., shear stress, tissue stretch, or pressure) or convective transport of morphogens, resulting in a biochemical gradient. If biophysical stimulation were the underlying mechanism, the observed developmental defects in the lymphatic vasculature would have been expected with only increase or decrease in flow, but not both. Therefore, we suggest that interstitial flow may be acting to develop and maintain a biomolecular gradient which serves as a cue to direct LECs to their intended destination. It is widely accepted that pressure gradients and dynamic stresses exist in all living tissues. 34 In developing embryos, morphogen concentration gradients are frequently more essential in providing spatial information that guides cell differentiation and migration than is the total amount of morphogen present. 34
A potential factor that could serve as a migratory morphogenic cue is vascular endothelial growth factor C (VEGF-C), which has been implicated in the LEC migration phase of lymphangiogenesis.60,61 VEGF-C is synthesized as a precursor protein with long C- and N-terminal propeptides flanking the VEGF homology domain. 61 The VEGF-C protein is synthesized in one cell and then is exported from that cell to migrate and signal to its receptors on other cells. During this migration process, VEGF-C distribution is subject to interstitial flow. We speculate that during this time, normal interstitial flow is generating a concentration gradient of VEGF-C for support of proper LEC migration. Data from in vitro and regenerative models supports this contention,38,41 and these include computer simulations that model VEGF concentrations under both static and active flow conditions. 38 These data revealed that in the absence of either VEGF or convective fluid flow, little lymphatic development occurred, suggesting both components are needed for proper vessel generation.
At approximately 48 hpf, LECs begin to migrate from the PL string ventrally to form the TD. 44 48 At this stage in development, VEGF-C expression has been shown to be localized in the hypochord, dorsal aorta (DA), and ventral mesenchyme. 47 Blood pressure in the zebrafish is highest in the DA, also the area where VEGF-C is expressed during the time of embryonic LEC migration. When fluid pressure within the DA changes, attendant alterations in interstitial flow alter the establishment of a normal VEGF-C gradient, resulting in failed LEC migration and subsequent lymph vessel development. According to this model (Fig. 6), if interstitial flow is increased, the VEGF-C gradient should be artificially extended and thus be more dilute and not send the proper signaling gradient for the LECs to migrate ventrally (Fig. 6B). Alternatively, if pressure within the DA is dramatically reduced, our model predicts that insufficient VEGF-C signal would expand dorsally, resulting in system dysregulation, as not enough to VEGF-C signal would reach the PL string to cue the LECs to migrate ventrally (Fig. 6C). Either scenario would result in a lymphatic phenotype similar to what we observed in our drug and genetic experiments.
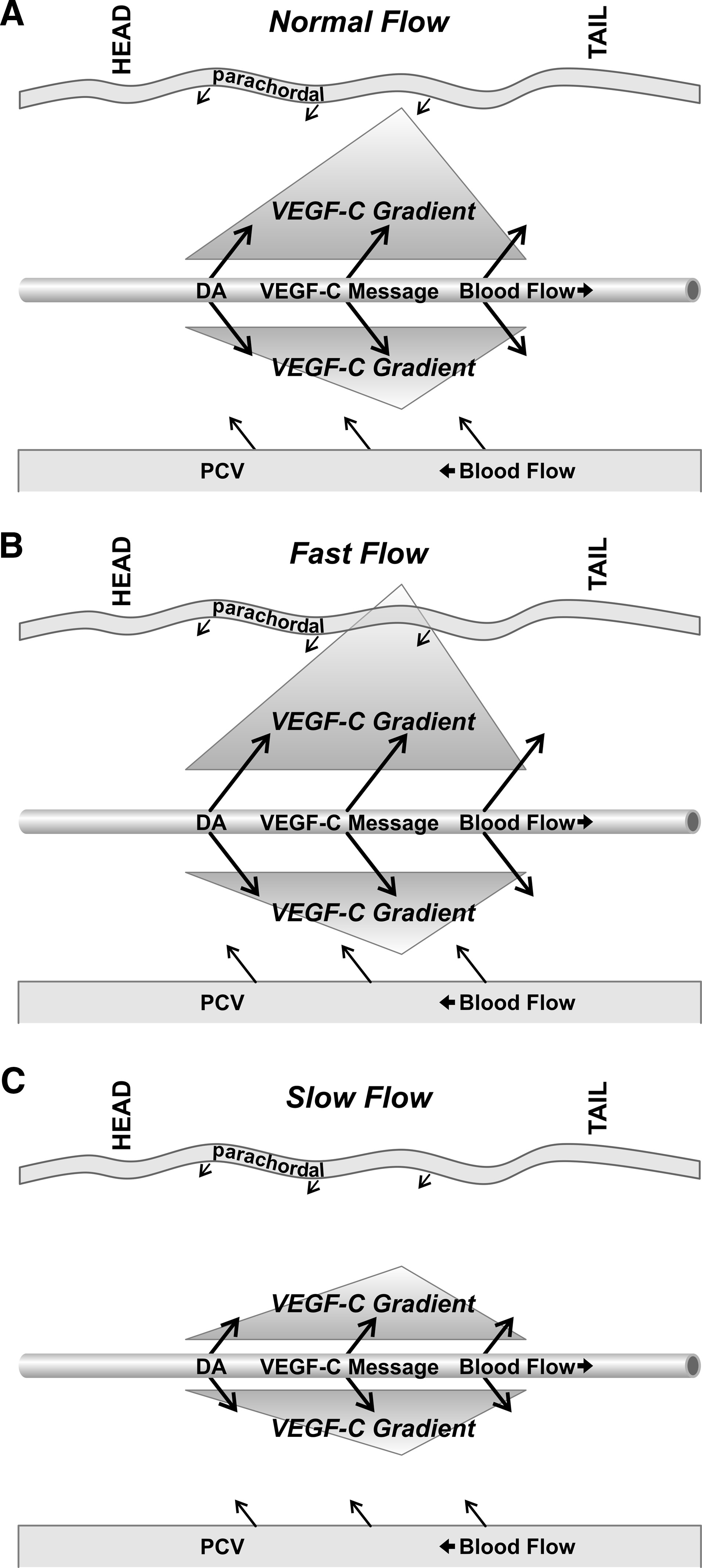
Model predicts that interstitial flow is responsible for setting up a gradient of VEGF-C to signal for proper migration of LECs, which originate in the PL string and migrate ventrally to form the TD.
In this study, we demonstrate that interstitial flow appears to be an important regulator of LEC migration. Citing expression profiles and functional data, we propose that interstitial flow is likely involved in setting up a VEGF-C gradient necessary for normal lymphatic development. Further, when interstitial flow is substantially altered, the redistribution of migratory cues may result in a skewed biomolecular gradient that fails to guide LECs properly, resulting in lymphatic tree ablation.
Footnotes
Acknowledgments
We would like to thank Mitul B. Desai for aid in embryo preparation. We would also like to thank Didier Stanier, Ph.D. of UCSF for generously providing the kdr-l:RasCherry x fli1:GFP fish. We thank Glenn Doerman for helping in figure preparation.
Author Disclosure Statement
Drs. Coffindaffer-Wilson and Hove and Mr. Craig have no conflicts of interest or financial ties to disclose.