
Abstract
Abstract
Background:
Secondary lymphedema, resulting from damage to lymphatic vessels, is a common sequela following surgical removal of lymph nodes for cancer. Current therapeutics for treating lymphedema are limited and further research on underlying causes is warranted. Published studies on molecular mechanisms of lymphedema primarily focus on lymphatic endothelial cells (LECs), which comprise the innermost lining of lymphatic capillaries and collecting vessels. However, traditional static culture of LECs may not adequately recapitulate the lymphedemous cell phenotype as transcriptomal comparison of human dermal LECs has shown significant differences in ex vivo and in vitro LEC gene expression. In this study, we designed a dynamic culture system, in which LECs were exposed to physiologic and excess mechanical strain to determine if native and lymphedemous phenotypes could be reproduced in vitro.
Methods and Results:
Purified human LECs were cultured in silicon dishes and subjected to 0% (control), 4%, and 8% mechanical strain for 72 hours. Our results indicate that control and stretched LECs maintained a mature phenotype. Extreme stretching at 8% strain significantly increased LEC proliferation and significantly increased Prox1 expression, suggesting a lymphedemous cell phenotype resulting with lymphangiogenesis.
Conclusion:
Mechanical strain reinforced a mature lymphatic phenotype and excess strain promoted lymphangiogenesis, while altering collagen deposition and cytokine secretion.
Introduction
T
Current research suggests that unlocking the causality between lymphatic vessel overload and lymphedema could give us a deeper understanding in the pathology of the disease. Studies suggest that histological changes in lymphatic vessels precede the onset of lymphedema. Okitsu et al. has shown in patients postlymphadenectomy that increased pressure inside lymphatic collecting vessels is preceded by extensive collagen deposition within vessel walls.6,7 Using histological sections from patients postlymphadenectomy, Mihara et al. demonstrated that lymphatic vessels became harder, lost elasticity, and potentially lost peristaltic capacity to transport lymph before the development of lymphedema. 7 This concept has also been studied in animals, in which Avraham et al. used a rat-tail model to demonstrate that tissue fibrosis is a key step in the development of lymphedema and prevents lymphatic regeneration. 8
However, existing evidence suggests that increased interstitial protein alone is not enough to induce fibrosis and inflammation in lymphedema. 9 Studies by Breslin and Kurtz have shown that lymphatic endothelial cells (LECs) change their morphology to better adapt to mechanical stress. 10 Therefore, in the present study, we sought to investigate the mechanical implications of lymphatic vessel overload and its relationship to the development of fibrosis and inflammation. We used uni-axial stretching to study LEC's response to mechanical stress. Physiologic lymphatic vessel activity was modeled using 4% stretching, and pathological activity was modeled with 8% stretch according to previous mechanobiological studies performed by Zhang and Wang. 11 LEC phenotype as well as the regulation of fibrotic and inflammatory genes were assessed.
Materials and Methods
Coating silicon dishes
Custom-made silicon dishes measuring 6 × 3 cm were each coated with 3 mL of 10 mg/mL of collagen solution (04902; Stemcell) for 2 hours at room temperature. The solution was aspirated and the dish was washed twice with PBS (70011-069; Invitrogen). The equipment and the technique were based on the protocol established by Zhang and Wang for tendon stem cells. 11 An additional 3 mL solution of 25 μg/mL fibronectin was then used to coat each dish for 2 hours at room temperature. The solution was aspirated and the dish was washed twice with PBS (Fig. 1).

The silicon dishes were 6 × 3 cm in dimension. The dish was coated with collagen and fibronectin to assist in cell attachment. The dish was covered with a glass lid anchored with a metal frame to prevent displacement during stretching. The dishes were pulled lengthwise 30 cycles per minute for 72 hours.
Culturing LECs
Adult human dermal LECs (C-12217; PromoCell) were then seeded at 3.0 × 105 cells per dish in 5 mL of VEGF-C-enriched media. The culture media consisted of endothelial basal medium-2 (cc-3156; Lonza) with 10 μg/mL hydrocortisone, 20% fetal bovine serum (16140-071; Gibco), 400 U/mL penicillin and streptomycin (17-602E; Lonza), 2 mmol
Cell conditioning
We modeled lymphatic activity with mechanical stretching in silicon dishes: 0% stretch served as control, 4% stretch represented physiological conditions, and 8% stretch represented lymphatic overload. The silicon dishes were stretched lengthwise at 0%, 4%, and 8% of the silicon dish at a rate of 30 cycles per minute for 72 hours. After 48 hours in standard culture, the medium was changed and four wells of cells were then cultured under each of the three conditions. The stretching apparatus was provided by the MechanoBiology Laboratory. The protein and RNA were harvested after 72 hours of continuous conditioning.
Reverse transcription-polymerase chain reaction (RT-PCR) assessment of inflammatory, fibrotic and lymphatic gene expression
Inflammatory markers IL1A, IL6, IFNG, and TNF were used. Fibrotic markers COL1A1 and COL3A1 were used. The LECs were characterized using LYVE1 and PROX1 (Taqman primers). All markers were normalized against GAPDH, a housekeeping gene.
Western blot was performed for collagen 1 and 3
Western blot for collagen 1 (34710; Abcam) and 3 (sc-28888; Santa Cruz) was used to confirm the presence of collagen and to provide further context for RT-PCR data on collagen production.
Results
Cell counts
At the end of 72 hours of conditioning, there were significantly more cells in the 8% stretch group than in the control group and the 4% stretch group (p < 0.05). The cell count was supported visually (Figs. 2 and 3).
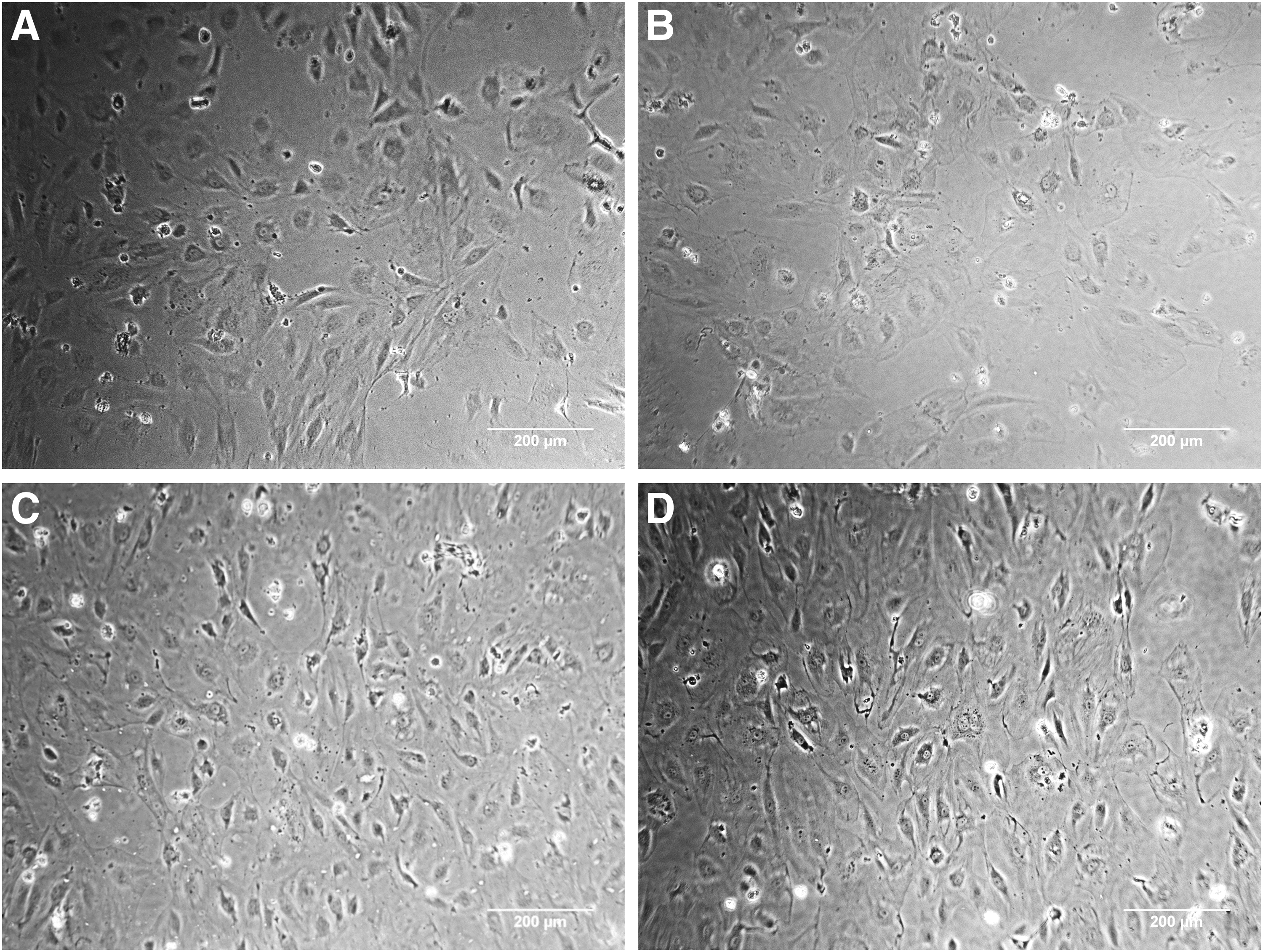
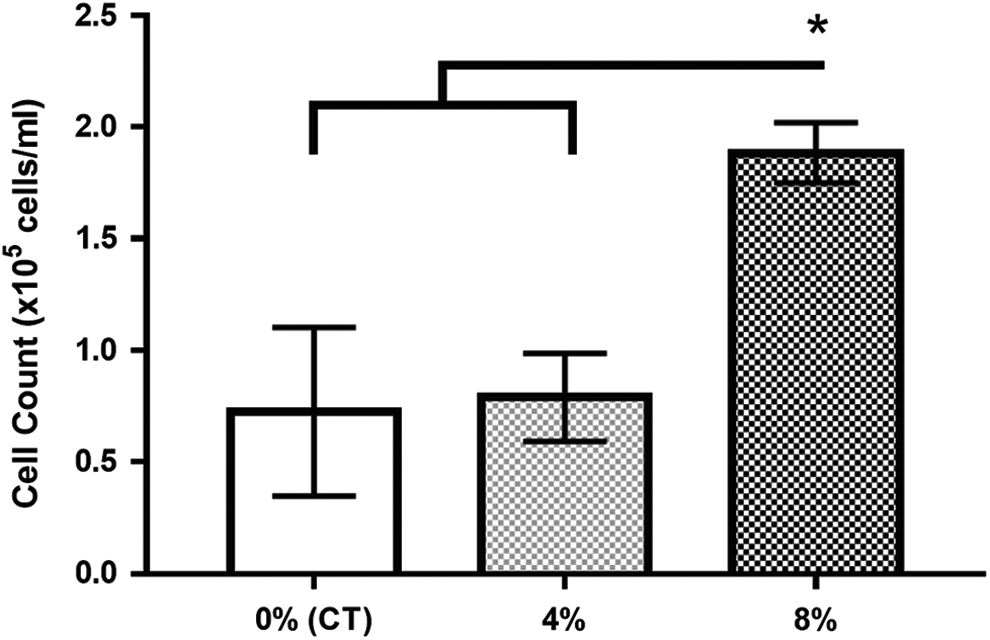
Cells conditioned at 8% stretch proliferated significantly more than those in the control group and the 4% stretch group. *p < 0.05.
LEC phenotype
Stretching appears to significantly influence gene expression and cellular phenotype in lymphatics. Although RT-PCR results indicate that the groups shared similar expression of the mature lymphatic marker, LYVE-1, there was significantly higher expression of Prox1 with 8% stretch than controls or 4% stretch, suggesting that lymphangiogenesis could be induced by intense stretching (p < 0.05) (Fig. 4).
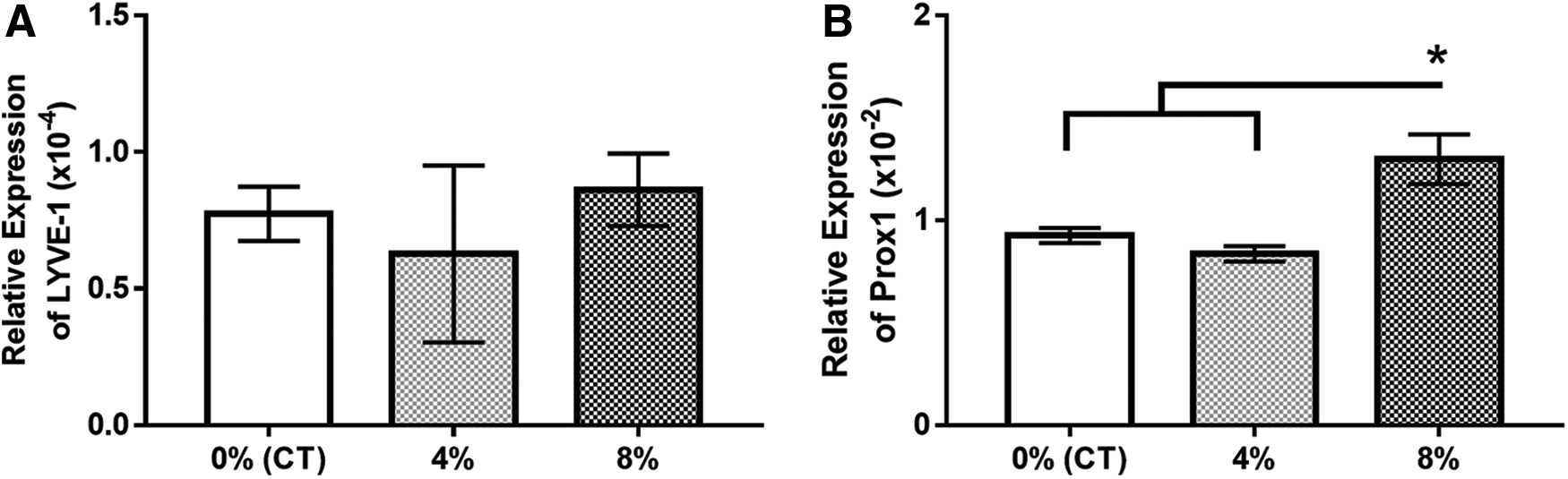
In qPCR data normalized against GAPDH:
Fibrosis and extracellular matrix deposition
There was no significant difference in COL1A1 between groups, though a similar trend of higher expression in control cells was observed with both RT-PCR and western blot analysis (Fig. 5). Relative expression of COL3A1 was significantly lower in control cells than in 4% and 8% stretching (p < 0.05).
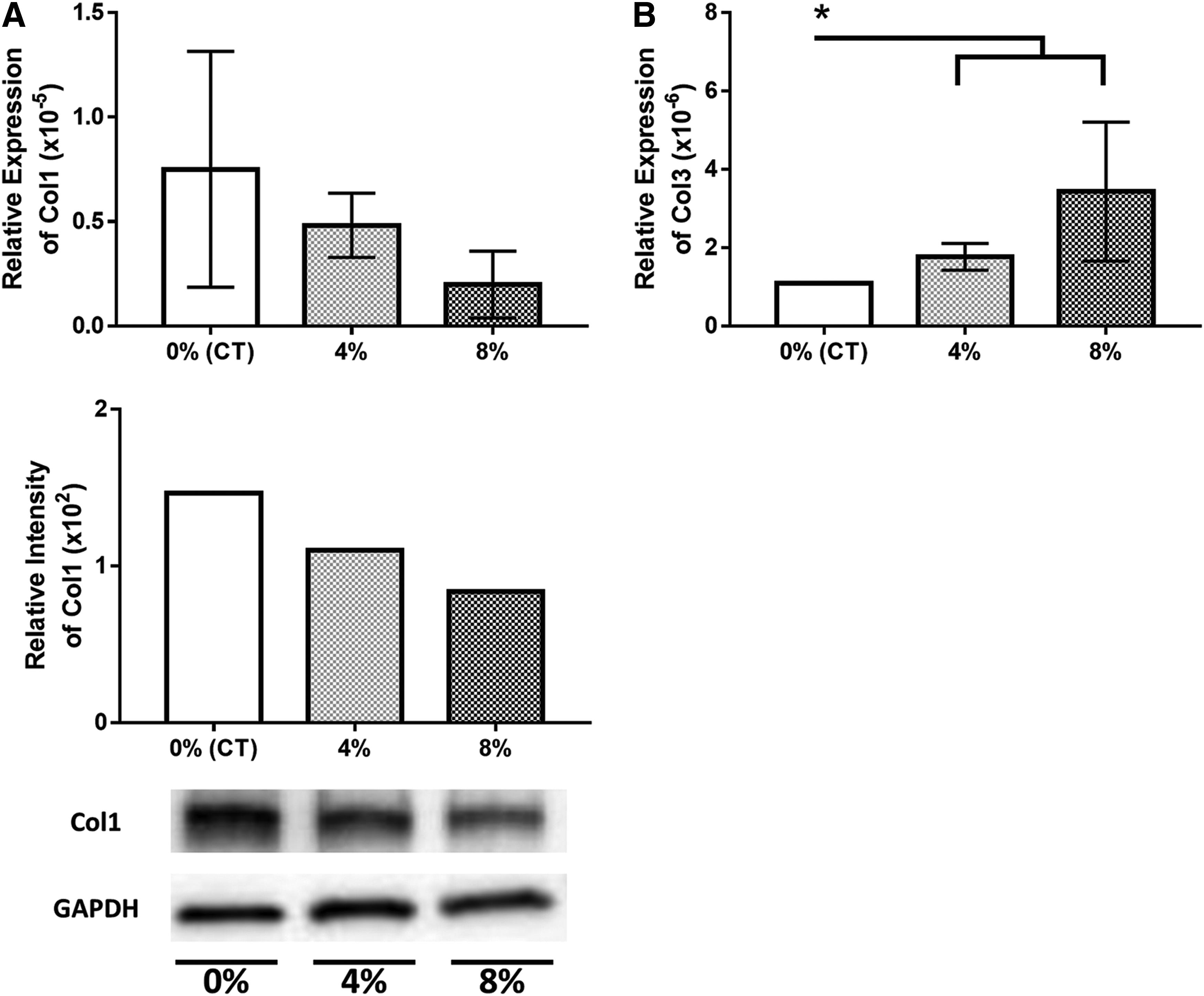
Inflammation
RT-PCR showed reduced expression of the inflammatory marker IL1A in the 8% stretch group compared with the 4% stretch group (p < 0.05), though expression was not significantly different from control cells. IL6 expression was also significantly lower in the 8% stretch group compared with both the control and 4% stretch group (p < 0.05). Results for IFNG and TNF expression showed very low expression of the inflammatory cytokines in all three groups with no significant differences between experimental groups (Fig. 6).
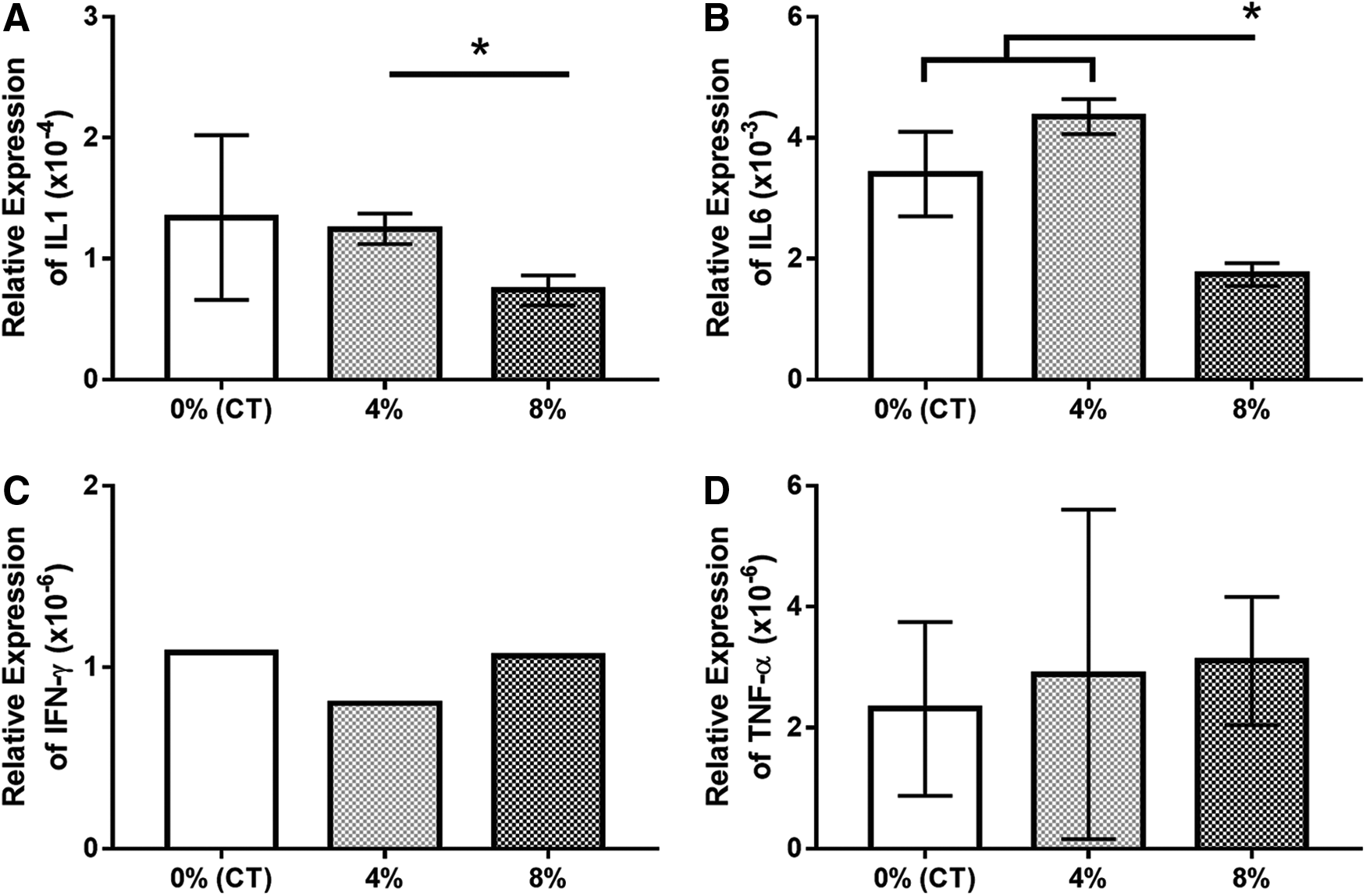
In qPCR data normalized against GAPDH:
Conclusion
Lymphatic capillaries are composed of a single layer of LECs attached to a discontinuous basement membrane by anchoring filaments that maintain vessel patency. Fluid movement from interstitium into the lymphatics is dependent on pressure gradients. Backflow is prevented by overlapping cell membrane junctions of VE-cadherin and PECAM-1 adhesion proteins. Once fluid has entered initial lymphatics, movement through vessels is determined by pressure gradients until entering collecting ducts, where intrinsic pumping mechanisms are required to move fluid against significant hydrostatic gradients. It has recently been observed that LECs are able to adapt their shape and membrane transport mechanisms under altered mechanical forces such as shear stress and increased luminal pressure to optimize fluid removal.12,13 For example, in response to decreased vessel afterload, LEC-derived nitric oxide and histamine cause a significant reduction in vessel contraction amplitude and frequency.14–16 During increased vessel afterload, LECs proliferation is induced through a mechano-induced colocalization of VEGFR3 and β1 Integrin, leading to lymphangiogenesis and increased interstitial fluid drainage.17,18
In our model, LECs appear to proliferate under stress
Inducing an 8% stretch resulted in significantly more cells than under control or 4% stretching conditions. Similar behavior has also been noted in some mesenchymal stem cell populations. 11 This cellular proliferation could contribute to the additional deposition of extracellular matrix present in lymphatic vessels of patients with lymphedema. 9 As such, we witnessed increased deposition of collagen 3, which is commonly found in blood vessels.
Stressed LECs also maintain mature phenotypes
All groups maintained similar levels of mature lymphatic cell markers LYVE-1. However, excessive lymphatic overload leads to lymphangiogenesis. Prox1 is a critical transcription factor for lymphangiogenesis and maintenance of lymphatic vessels. 19 Prox1 expression was significantly higher in the 8% stretching group than the control and 4% group. Prox1 expression in mature lymphatic vessels is independent of external stimuli, and Prox1-positive LECs are capable of migrating and forming lymphatic sacs and generating new lymphatic vessels.20,21
Stretching increased expression of collagen 3
Control groups had significantly lower COL3A1 expression than both stretched groups. The 8% stretch group also trended higher than 4% in COL3A1 expression. As mentioned previously, the increased deposition of COL3A1 could be attributed to lymphangiogenesis and cell proliferation. However, we did not expect to see a decrease in production of COL1A1 in both qPCR and western blots. This difference may be attributed to altered ratio of protein synthesis; therefore, it will be important to evaluate other extra cellular matrix protein deposition to obtain a more comprehensive picture of the fibrotic pathway.
Inflammatory response may require longer time points to assess
We originally anticipated that 8% stretch-conditioned cells would respond to stress through a variety of inflammatory cytokines such as IL1, IL6, IFNγ, and TNFα, however, this was not the case. The 8% stretch group had lower expression of IL1A and IL6 than the 4% stretch group. Overall, there was no trend in inflammatory response. The inflammatory response may require additional mechanical stimulation and a broader panel of markers to observe.
This study is not entirely without limitations. As a pilot study, sample size and conditioning duration was constrained, and a more robust, longer duration study will be necessary to further explore the regulation of stretched LECs. However, this study is the first to simulate lymphatic vessel overload conditions in LECs in vitro and highlights the importance of mechanical stimulus in maintaining cellular phenotype. This is a very important step for developing clinically relevant models to advance our understanding of the pathology of lymphedema and to pave the way for therapeutic testing.
Footnotes
Author Disclosure Statement
No competing financial interests exist.