
Abstract
Abstract
Background:
A problematic complication after radiation therapy is lymphedema. Development of lymphedema is associated with an increase in lymphatic paracellular permeability. The current study investigated the effects of radiation on intercellular junctions and paracellular permeability in cultured human dermal lymphatic endothelial cells (HDLECs).
Methods and Results:
Double immunofluorescence staining with vascular endothelial (VE)-cadherin and actin immediately after X-ray irradiation (5 or 20 Gy) was performed. Morphological changes induced by irradiation were assessed. Cell viability and paracellular permeability after irradiation were also evaluated. Broad junctions in which VE-cadherin was accumulated at cell–cell contacts and almost colocalized with actin were significantly decreased in a dose-dependent manner in confluent and sparse irradiated HDLECs. Irradiation shortened the width of VE-cadherin-positive areas at the cell–cell contacts. Actin filaments did not colocalize with VE-cadherin after 20 Gy irradiation. Although cell viability was not affected by irradiation, paracellular permeability significantly increased in a dose-dependent manner.
Conclusions:
A dose of 5 or 20 Gy irradiation in HDLECs does not affect cell viability, but changes VE-cadherin mediated intercellular junctions and actin structure, resulting in an increase of paracellular permeability. Further investigations on the regulatory proteins involved in radiation-induced changes, which were observed in the current study, may contribute to development of lymphedema therapy.
Introduction
O
One of the main factors in development of lymphedema may be an increase in lymphatic paracellular permeability. Intercellular junctions (tight junction, adherens junction, desmosome, and gap junction) play essential roles in the maintenance of vascular integrity and permeability.10–12 Vascular endothelial (VE)-cadherin is a transmembrane protein and major component of endothelial adherens junctions, which are responsible for intercellular mechanical adhesion. VE-cadherin associates with various intracellular proteins, such as zonula occludens-1 and β-catenin, and connects to the cytoskeletal protein actin.10–12 The connection between adherens junctions and the actin cytoskeleton mediated by VE-cadherin is thought to be crucial for regulating blood VE function. 13 The gap junction proteins, connexins (Cxs), are associated with lymphatic disorders such as lymphedema and chylothorax.14,15
Although some in vitro studies have focused on the effects of ionizing radiation on intercellular junctions in blood VE cells,1,16 few in vitro studies have analyzed radiation effects on lymphatic endothelial intercellular junctions. Therefore, the current study aimed to better understand the underlying etiology of radiation-induced lymphedema. The effects of ionizing radiation on intercellular junctions in cultured human lymphatic endothelial cells were investigated.
Materials and Methods
Cell culture
Human dermal lymphatic endothelial cells (HDLECs) were obtained from ScienCell Research Laboratories (Carlsbad, CA). HDLECs were routinely maintained on fibronectin-coated cell cultureware surfaces (Sekiyarika, Tokyo, Japan) in Endothelial Cell Medium (ScienCell Research Laboratories) supplemented with co-delivered mixed additives. This resulted in a final concentration of 1% endothelial cell growth supplement, 5% fetal bovine serum, and 1% of penicillin/streptomycin. Cells that were used in the experiments were between passages 2 and 10 counting from the stage of primary culture, for which the stable expression of investigated proteins had been confirmed.
Irradiation
Irradiation was performed using the MBR-1505R2 X-ray generator (Hitachi, Tokyo, Japan) with the following conditions: 150 kV, 4 mA, 1-mm aluminum filter, at room temperature. The details of radiation machine can be referred in our recent report. 17 Cells were irradiated with doses of 5 or 20 Gy (∼0.6 Gy/min). After irradiation, each experiment was immediately performed.
Reagents and antibodies
Goat anti-VE-cadherin antibody (C-19) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Rabbit anti-Cx43 antibody (ab11370) was purchased from Abcam (Cambridge, United Kingdom). Actin filaments were labeled with Alexa 568-Phalloidin (Invitrogen Molecular Probes, Karlsruhe, Germany). The nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI; Sigma-Aldrich, St. Louis, MO). Secondary antibody conjugated with Alexa 488 and 555 were also purchased from Invitrogen.
Immunofluorescence staining and microscopy
For immunofluorescence microscopy, cells were grown on micro cover glass that was coated with fibronectin stock solution (Sigma-Aldrich) in 35 × 10-mm-style polystyrene tissue culture dishes. Cells were fixed with 1% formaldehyde in phosphate-buffered saline (PBS) for 10 minutes at room temperature, treated with 0.2% Triton X-100 in PBS for 5 minutes, and washed with PBS. Cells were blocked with 1% bovine serum albumin in PBS overnight and then incubated with primary antibodies overnight. Cells were then rinsed thrice with PBS and incubated with appropriate secondary antibodies for 30 minutes. After rinsing with PBS, the specimens were embedded in FluorSave (Calbiochem, Darmstadt, Germany). Acquisition of images was performed with a BZ-X 700 (Keyence, Osaka, Japan).
Quantification of cell numbers and VE-cadherin-positive broad areas
We previously described the characteristic intercellular junctions in HDLECs. 18 In brief, the number of cells with a cell periphery, in which a broad distribution of VE-cadherin could be observed, were counted in the same images as those used to count the cell number (Fig. 1). Cell junctions with a “VE-cadherin-positive broad area” were designated as “broad junctions” in this study. The number of cell junctions without a VE-cadherin-positive broad area was also counted, and the number of total cell junctions was calculated. The cell number was counted in five randomly selected independent microscopic images (20 × ). The results were representative of three independent experiments.

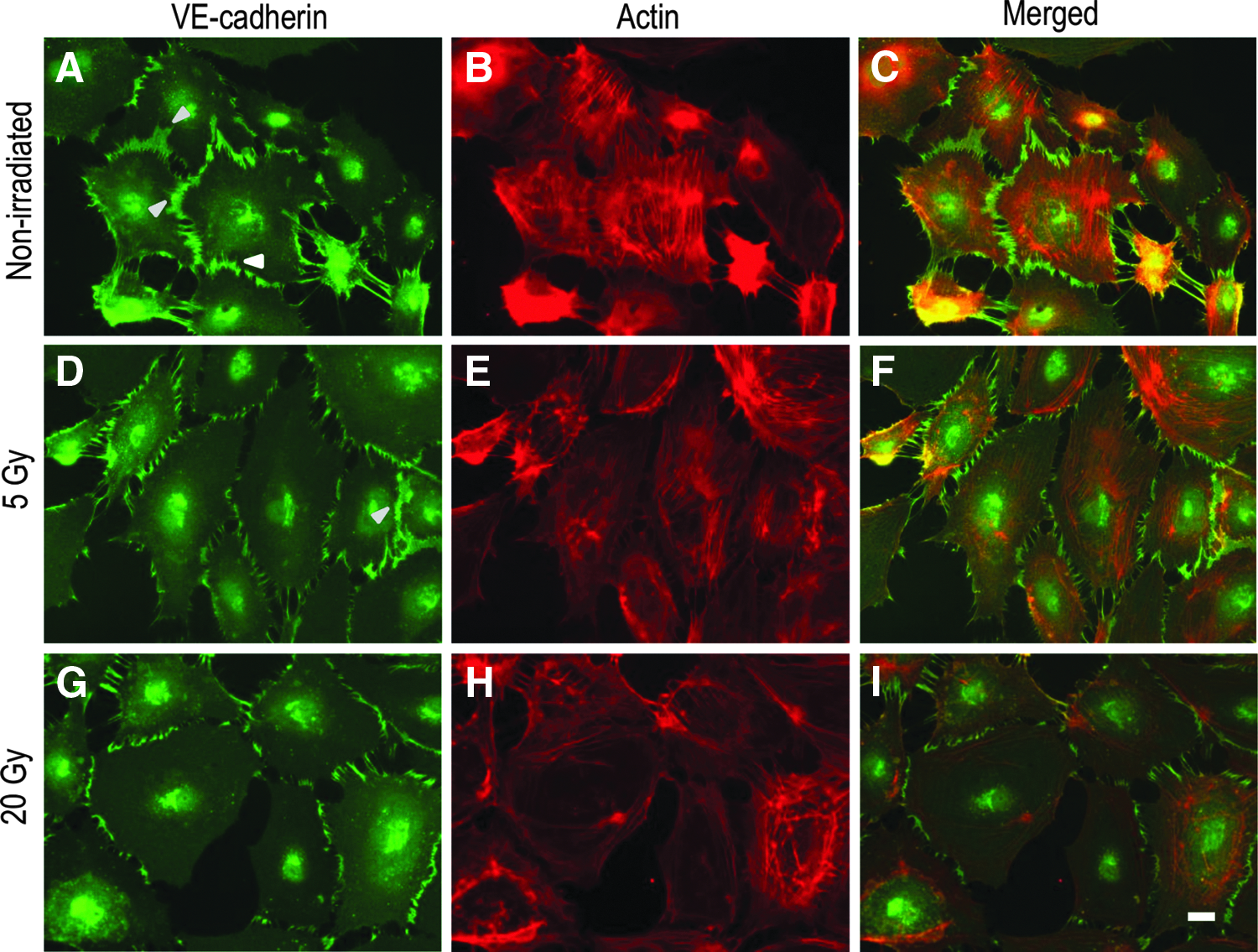
Double immunofluorescence staining for VE-cadherin (green) and actin (red) in confluent HDLECs. In nonirradiated cells, broad junctions are dominant, and VE-cadherin and actin are almost colocalized
Paracellular permeability assay
Paracellular permeability was evaluated with a 24-well in vitro Vascular Permeability Assay Kit (ECM644; Millipore, Billerica, MA) according to the manufacturer's protocol. Briefly, 1 × 105 cells were seeded into a collagen coated insert (1-μm pores within a polyethylene terephthalate membrane). Cells were incubated for 72 hours until a monolayer was formed. A volume of 150 μL of fluorescein isothiocyanate–dextran solution was added to each insert followed by incubation for 30 minutes at 37°C. Cells were irradiated in a dose of 5 or 20 Gy. Immediately after irradiation, 100 μL of the solutions from each well of the receiver tray were transferred to a 96-well plate. Absorbance at 485 and 535 nm was measured with the Fluoroskan Ascent FL (Thermo Fisher Scientific, Vantaa, Finland).
Cell viability assay
Cell viability was evaluated with the Cell Counting Kit-8 (Dojindo Laboratories, Kumamoto, Japan). This assay is based on the conversion of water-soluble tetrazolium salt, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt (WST-8) to a water-soluble formazan dye upon reduction by dehydrogenase in the presence of an electron mediator. This assay was performed according to the manufacturer's protocol. In brief, 5 × 104 cells were seeded in a 96-well plate. Forty-eight hours after incubation, cells were irradiated in a dose of 5 or 20 Gy. A volume of 10 μL of the Cell Counting Kit-8 solution was added to each well followed by incubation for 120 minutes. Absorbance at 450 nm was measured with the EnSpire Multimode plate reader (PerkinElmer, Waltham, MA).
Statistical analysis
All data are expressed as mean ± standard deviation of three to five independent experiments. R software (R Development Core Team, 2011) was used to analyze the data with one-way analysis of variance, and Tukey's multiple comparisons were used as a post hoc test. A p-value less than 0.05 was regarded as indicating statistical significance.
Results
The intercellular junctions in HDLECs can be divided into two types: continuous linear and discontinuous zig-zag junctions. 18 In some continuous junctions, VE-cadherin robustly accumulates not only at cell–cell contacts but also near the periphery of the cell. We have designated this characteristic pattern of VE-cadherin distribution in HDLECs as a VE-cadherin-positive broad area. 18 Continuous junctions are similar to stable and mature endothelial cell junctions, and a broad distribution of VE-cadherin at the periphery of cells may reflect lymphatic permeability.18,19
Consistently, the immunofluorescence signal for VE-cadherin was broadly detected near the periphery of the cell, as well as at cell–cell contacts in confluent nonirradiated HDLECs (Fig. 1A–C). Double immunofluorescence staining showed that irradiation with a dose of 5 Gy slightly decreased the VE-cadherin-positive broad area (Fig. 1D–F). A dose of 20 Gy irradiation appeared to decrease broad junctions (Fig. 1G–I). The immunofluorescence signals of VE-cadherin and actin were almost merged in confluent nonirradiated cells. In irradiated cells, the immunofluorescence signals of actin were strong near the periphery of the cell and did not merge with the immunofluorescence signals of VE-cadherin (Fig. 1F’, I’). The actin structures observed in the irradiated cells appeared to be similar to cortical actin belts, which show cortical enrichment of actin, arranged parallel to endothelial cell junctions. 20 Importantly, the characteristic actin structures in irradiated cells were not similar to stress fibers connecting to stress fibers of adjacent cells through discontinuous intercellular junctions induced by a major pro-inflammatory cytokine, tumor necrosis factor-α (TNF-α).18,21
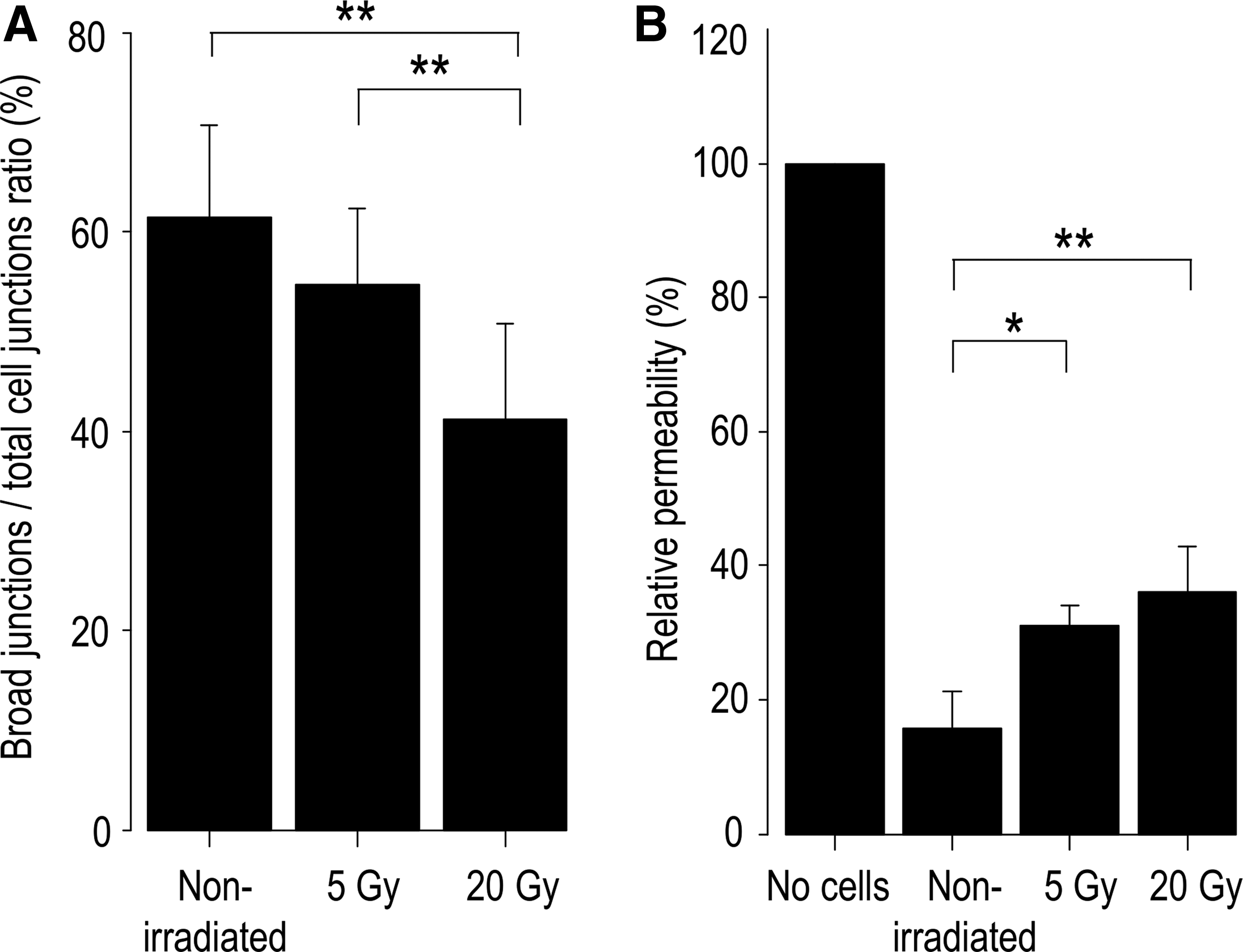
The same phenotypes induced by irradiation in confluent irradiated HDLECs were observed in the sparse culture (Fig. 2). In sparse nonirradiated HDLECs, broad linear junctions and discontinuous zig-zag junctions were intermingled (Fig. 2A–C). In sparse irradiated cells, the VE-cadherin-positive broad area appeared to decrease in a dose-dependent manner. Particularly, the irradiation shortened the width of the VE-cadherin-positive area at the cell–cell contacts. This change in the intercellular junctions, namely shortening of the VE-cadherin-positive broad area, was more evident in the 20 Gy-irradiated cells than in the 5 Gy-irradiated cells (Fig. 2D–I). Actin filaments were strongly accumulated parallel to cell junctions near the periphery of the cell and did not connect to VE-cadherin-positive intercellular junctions in the 20 Gy-irradiated cells (Fig. 2G–I). After counting the number of broad junctions and the total number of cell junctions, the percentage of broad junctions was significantly decreased in a dose-dependent manner (Fig. 3A), while the total cell number and cell viability were not affected by irradiation (Supplementary Fig. S1; Supplementary Data are available online at www.liebertpub.com/lrb).
Double immunofluorescence staining for VE-cadherin (green) and actin (red) in sparse HDLECs.
Dose-dependent decrease in broad junctions
To confirm the radiation-induced changes observed in immunofluorescence staining, a permeability assay was performed. Irradiation significantly increased paracellular permeability in a dose-dependent manner and reflected the morphological changes observed in immunofluorescence staining (Fig. 3B).
Finally, we evaluated the expression of Cx in irradiated HDLECs, because Cx deficiencies result in lymphatic disorders, including lymphedema and chylothorax. 15 Cx43, one of the major gap junction proteins in human VE cells, 22 colocalized with VE-cadherin in the broad junction in the nonirradiated and 5 Gy-irradiated cells (Fig. 4A–H). In the 20 Gy-irradiated cells, the immunofluorescence signals of Cx43 were decreased at the VE-cadherin positive discontinuous cell–cell contacts (Fig. 4I–L).
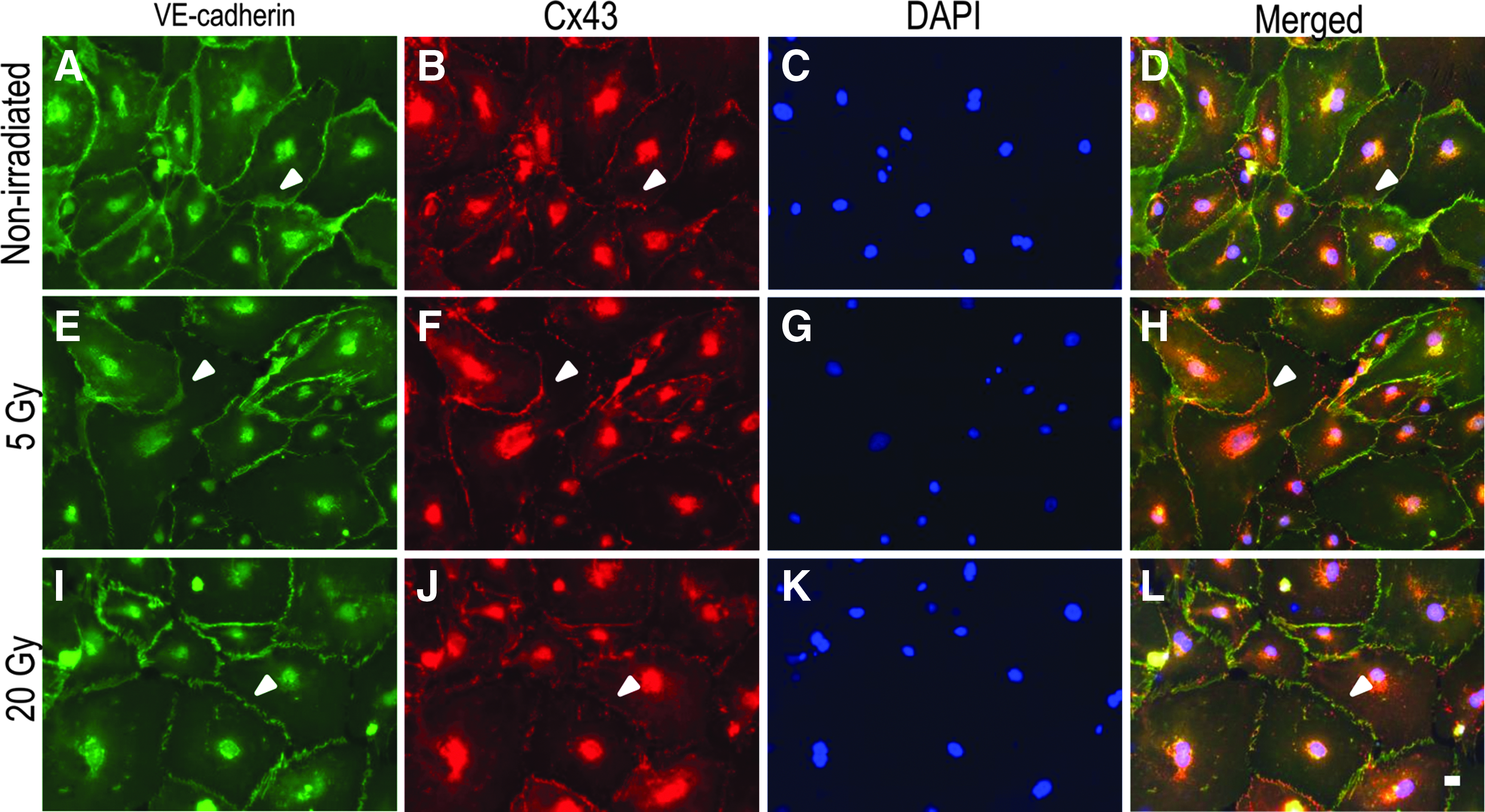
Triple immunofluorescence staining for VE-cadherin (green), Cx43 (red), and DAPI (blue) in confluent HDLECs.
Discussion
One of the problematic complications after radiation therapy for cancer is lymphedema. To develop therapy for lymphedema, the underlying etiology should be investigated. The current study showed that irradiation decreased the number of intercellular junctions in areas where VE-cadherin was broadly accumulated at cell–cell contacts and increased paracellular permeability in a dose-dependent manner.
The changes in intercellular junctions induced by irradiation were not similar to changes induced by the permeability-increasing agent TNF-α. 18 Our previous study shows that TNF-α stimulation alters continuous linear junctions to discontinuous zig-zag junctions in HDLECs. 18 In human umbilical vein endothelial cells (HUVECs), TNF-α stimulation also increases discontinuous adherens junctions and actin stress fibers. 21 Stress fibers are attached to VE-cadherin-mediated junctional complexes, and this formation is due to actin–myosin (force-producing motor protein) contractility in HUVECs.21,23 Stress fibers in blood VE cells regulate permeability to small solutes and may also help to withstand the mechanical stress generated by leukocyte transmigration under shear stress. 21 In contrast, radiation-induced changes of intercellular junctions in HDLECs involve shortening of the VE-cadherin-positive area at cell–cell contacts and disconnection between VE-cadherin junctions and actin filaments.
Endothelial cells in the vasculature have heterogeneous and organ-specific properties. 24 Injury to microvessels is more severe and qualitatively different from that in larger vessels. 25 In a previous study by Gabryś et al., 1 5–20 Gy irradiation caused a dose-dependent appearance of stress fibers. In addition, this dose altered continuous linear junctions to segmented and discontinuous junctions in dermal microvascular endothelial cells (DMECs). Actin remodeling and an increase in permeability in DMECs are regulated by a small G protein (RhoA). 1 The phenotype induced by irradiation in DMECs appears to be similar to that induced by TNF-α in HUVECs and HDLECs. Interestingly, irradiation does not induce actin remodeling and redistribution of VE-cadherin in HUVECs. 1 The difference in radiation-induced phenotype between DMECs and HUVECs is probably due to the response to radiation between large and small vessels, and signaling consequences of radiation in various endothelial cells are considered to be tissue dependent. 1
The lymphatic endothelium has site-specific, specialized intercellular junctions in each different lymphatic vascular component, such as capillaries, precollecting ducts, and collecting ducts. 26 HDLECs probably have properties of initial lymphatics (lymphatic capillaries) because they are derived from dermal lymphatic capillaries. 18 An in vivo previous study reported that lymphatic endothelial cells in mice in the small intestine and peri-tumor areas were highly resistant to radiation injury, while blood vessel endothelial cells were relatively sensitive to radiation. 27 In the mouse small intestine, 15 Gy of irradiation induces microvascular, but not lymphatic, endothelial apoptosis and does not alter VE growth factor-C and -D and VE growth factor receptor-3 expression. 27 Another in vivo study by Avraham et al. 28 reported that irradiation caused loss of dermal capillary lymphatics in dose- and time-dependent manners. The same authors' in vitro study using HDLECs showed that radiation induced apoptosis with a dose of 30 Gy, while 15 Gy irradiation did not induce apoptosis. 29 In the current study, the total number of cells and cell viability in HDLEC were not affected by 5 or 20 Gy irradiation. The discrepancy of the results between the current study and the study by Avraham et al. 28 is probably due to differences in irradiation doses and the time interval between experiments and irradiation.
Interestingly, Avraham et al. 28 additionally noted that HDLECs were sensitive to radiation in terms of cellular senescence. Senescence was induced 4 days after only 4 Gy irradiation in HDLECs, and the percentage of senescence increased in a dose-dependent manner (4, 8, or 12 Gy). 28 Cellular senescence is defined as an irreversible growth arrest occurring in cultured cells after many population doublings, even though nutrients, growth factors, and sufficient space are available for cells to divide. 29 In senescent HUVECs, expression of VE-cadherin is normal, but the distribution pattern of VE-cadherin at cell–cell contacts is different from that in nonsenescent cells. 29 Adherens junctions appear swollen and disorganized in senescent HUVECs. 29 Expression of claudin-5 (a major component of VE tight junctions) significantly decreases in senescent HUVECs, resulting in an increase of permeability compared with nonsenescent cells. 29 Notch signaling triggers early senescence in low-passage HUVECs, and the senescent phenotype is associated with an increase in monolayer permeability. This is evident with disrupted VE-cadherin and β-catenin levels and localization. 30 The senescent phenotype accompanied by endothelial barrier dysfunction in HUVECs is regulated by RhoA in Notch signaling. 30 Notch activation induces dissolution of the dense peripheral actin band, and the reticular cortical actin network is replaced by thick stress fibers that are arranged parallel to the spindle axis of cells. 30 The changes in distribution of VE-cadherin and actin structure induced by irradiation in HDLECs in the current study are not similar to the Notch signaling-induced senescent phenotype. Whether radiation-induced changes in HDLECs are due to cellular senescence still remains unclear. However, RhoA is probably not involved in radiation-induced changes in HDLEC because irradiation did not induce stress fiber formation in HDLECs in the current study.
Cxs (Cx37, Cx43, and Cx47) are expressed in the developing lymphatic vasculature and, when defective, are associated with lymphatic disorders in mice. 15 E16.5 embryos lacking Cx37 and Cx47 exhibit lymphedema, and lymphatic valves are completely absent in E18.5 mesentery samples lacking Cx43 but not Cx37. 15 In rat lung microvessels, suppression of Cx43 increases VE-cadherin expression, suggesting that reduction in Cx43 levels may modulate VE-cadherin levels. 31 In human, mutations in the Cx47 gene are known to increase the risk for secondary lymphedema following breast cancer treatment. 14 In epithelial cells, adherens junctions (cadherin–catenin complex) recruit plasma membrane protein such as Cx43 through microtubules. 32 The results in the current study probably indicate that the accumulation of Cx43 at cell–cell contacts depends on the formation of adherens junctions.
There are some limitations in this study. First, we only evaluated the acute phase following irradiation. In DMECs, radiation induces biphasic RhoA signaling. An initial burst in RhoA activity correlates with rapid cytoskeletal remodeling. A second peak of activity accounts for the sustained contractile and hyperpermeable cell phenotype. 1 Another limitation is that we could not identify which small G proteins were involved in radiation-induced changes in HDLECs. However, an increase in paracellular permeability following irradiation in HDLECs probably mimics one of the underlying mechanisms of lymphedema after radiation therapy. Therefore, further investigations on small G proteins involved in radiation-induced changes may contribute to development of lymphedema therapy.
Footnotes
Acknowledgments
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.