
Abstract
The CCAAT/enhancer-binding protein (C/EBP)β belongs to the C/EBP family of proteins that possesses a basic leucine zipper DNA-binding domain. These proteins bind DNA by dimerization and play a role in the transcriptional regulation of various cells. There are six different types of C/EBPs, and some form isoforms through the use of alternative translation initiation sites. The functional analysis of the C/EBP family is therefore difficult to achieve. Here we report on the production of specific monoclonal antibodies against mouse C/EBPβ using a rat medial iliac lymph node method. Immunoblotting using C/EBPβ monoclonal antibodies identified two types of isoforms, while immunostaining revealed a subnuclear localization for C/EBPβ. Use of this antibody should contribute to the further elucidation of the transcriptional regulatory function of C/EBPβ.
Introduction
T
C/EBPα and β generate translational isoforms by using alternative translation initiation. Three types of C/EBPβ translational isoforms have been identified in humans: p38 (a liver activating protein, LAP*), p33 (a LAP), and p20 (a liver inhibitory protein, LIP).(10,23,24) LAPs have three transactivation domains (TAD) that function as activators of transcription, but these are absent from LIP (Fig. 1).(23) C/EBPβ also contains two regulatory domains (RDs), which modulate its transcriptional activity.(19)
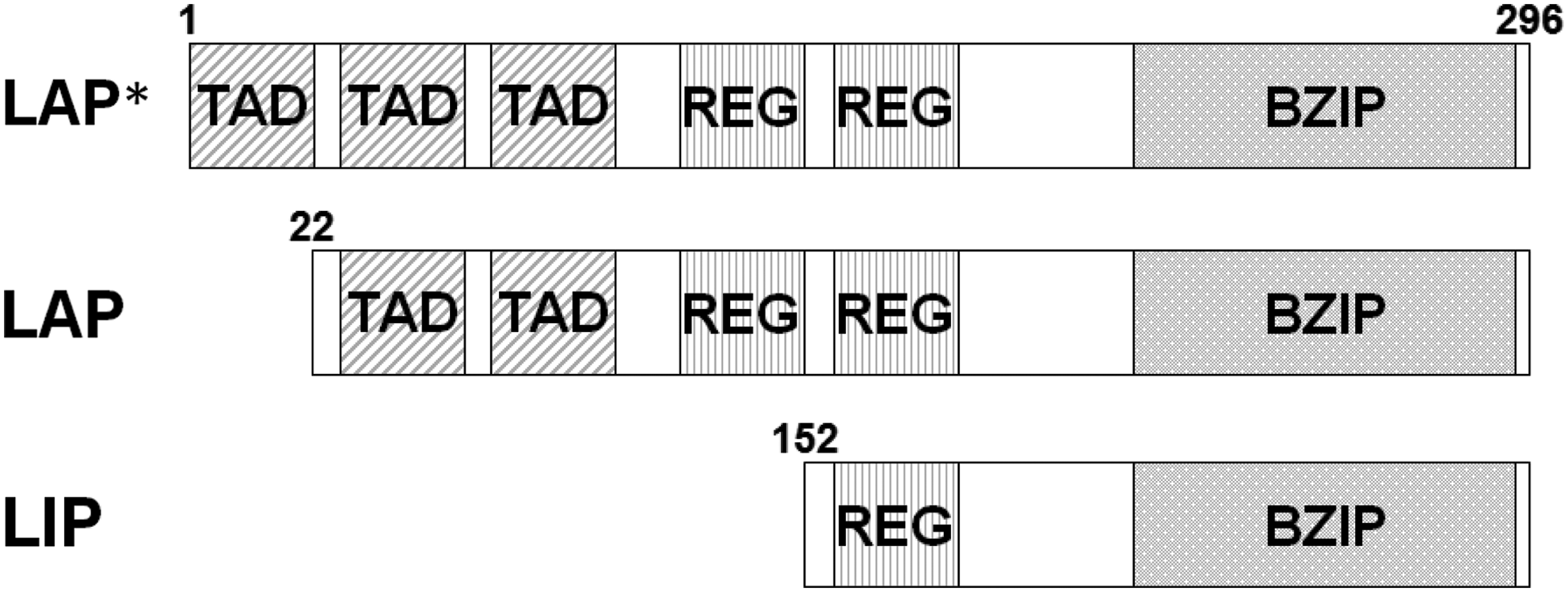
Schematic of three isoforms of C/EBPβ. mRNA of C/EBPβ directly translated initiation to alternative start sites from each N-terminal amino acid position. This results in the generation of different protein isoforms of C/EBPβ, termed LAP*, LAP, and LIP proteins, which differ in their N-terminal length causing the differential presence of N-terminal transactivation (TAD) and regulatory domains (RD) but common C-terminal basic leucine zipper domains (BZIP).
The various functions of C/EBPβ are limited by its different isoforms and post-translational modifications.(25,26) While C/EBPβ has been shown to be an important factor in cell differentiation, it remains unclear how it regulates precursor cells or differentiated cells. Therefore, to further elucidate the function of C/EBPβ, we developed a specific monoclonal antibody for mouse C/EBPβ in the present study.
Materials and Methods
Cell culture
Mouse L929 cells were derived from normal subcutaneous areolar tissue and were grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) in a humidified atmosphere of 5% CO2 at 37°C.
Production and purification of recombinant proteins
A full-length C/EBPβ fused glutathione S-transferase (GST) protein expression vector was transformed into Escherichia coli BL21(DE3) cells (Novagen, Madison, WI). Purification of the fusion protein was performed as previously described,(27) and E. coli cells were grown in LB medium containing 50 μg/mL carbenicillin (Nacalai tesque, Kyoto, Japan) at 37°C.
Rat immunization and monoclonal antibody production
The anti-C/EBPβ rat monoclonal antibody was produced using the rat lymph node method established by Sado and colleagues.(28,29) The hind footpads of 10-week-old female WKY/NCrj rats (SLC, Shizuoka, Japan) were injected with 150 μL of an emulsion containing 125 μg of GST-fused C/EBPβ protein and Freund's complete adjuvant. After 2 weeks, cells isolated from the medial iliac lymph nodes of these rats were placed in a 50% polyethylene glycol solution (PEG 1500, Roche, Mannheim, Germany) and fused with mouse myeloma SP2 cells at a ratio of 5:1. The hybridoma cells were plated in 96-well plates and selected in HAT selection medium (Hybridoma-SFM [Invitrogen, Carlsbad, CA], 10% FBS, 10% BM condimed H1 [Roche], 100 μM hypoxanthine, 0.4 μM aminopterin, and 16 μM thymidine). Seven days post-fusion, the hybridoma supernatants were screened by an enzyme-linked immunosorbent assay (ELISA) against the GST-fused C/EBPβ protein. Positive clones were subcloned and rescreened by ELISA. Monoclonal antibody (MAb) 7H5 and 7D2 immunoglobulin classes were a rat IgG2a (κ), which was identified using a rat isotyping kit.
Immunoblotting
Whole cell extracts of mouse L929 cells were separated by 10% SDS-PAGE and electrophoretically transferred to Immobilon-P PVDF membranes (Millipore, Bedford, MA). The membranes were blocked for 1 h at room temperature (RT) with a blocking solution containing 3% skim-milk in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20), and then incubated for 1 h at RT with anti-C/EBPβ rat monoclonal antibodies 7H5 and 7D2 diluted in the blocking solution. After washing with TBS-T, the membranes were incubated for 1 h at RT with alkaline phosphatase-conjugated anti-rat IgG antibody (Sigma, St Louis, MO). After washing with TBS-T, the membranes were treated with nitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolyl phosphate (BCIP).
Immunocytochemistry
L929 cells grown on coverslips were fixed with 3.7% formaldehyde for 15 min at RT, then washed twice with PBS. After a further rapid washing with PBS, cells were permeabilized with 0.5% Triton X-100 in PBS for 5 min, then incubated for 30 min in blocking buffer. The visualization of C/EBPβ was performed by incubating with MAbs 7H5 and 7D2, followed by an Alexa 488-conjugated goat anti-rat IgG (Invitrogen) for 1 h at RT. Coverslips were mounted with Prolong (Molecular Probes, Carlsbad, CA), and C/EBPβ localization was detected by fluorescence microscopy (Olympus, Tokyo, Japan).
Results and Discussion
To create a specific monoclonal antibody against mouse C/EBPβ, we utilized the rat lymph node method using full-length recombinant GST-fused mouse C/EBPβ. Two positive clones, 7H5 and 7D2, were identified by ELISA screening. Immunoblotting using whole cell extracts of mouse L929 cells revealed a strong band at 33 kDa and a weak band at 37 kDa for 7H5 and 7D2 (Fig. 2A). Full-length mouse C/EBPβ has 296 amino acids, with a deduced molecular weight of 31.4 kDa, while predicted molecular weights of mouse C/EBPβ translational isoforms are 31.4 kDa for LAP*, 29.1 kDa for LAP, and 15.6 kDa for LIP (Fig. 1). We therefore concluded that the 33 kDa band represented LAP and the 37 kDa band LAP*, as previous studies indicated that p33 is mainly translated in mouse embryonic fibroblasts (MEF), NIH3T3, and 3T3-L1 cells.(30–33) No band corresponding to LIP was detected (data not shown). Mouse C/EBPβ is known to be localized in the nucleus, which was confirmed by immunostaining using 7H5 and 7D2 monoclonal antibody clones (Fig. 2B).

C/EBPβ-specific monoclonal antibodies produced by 7H5 and 7D2 were recognized by C/EBPβ. (
C/EBPβ plays a role in many different physiological and pathological processes, including metabolism, cell survival, apoptosis, inflammation, and tumorigenesis. It also regulates the differentiation of several cell types such as adipocytes, chondrocytes, osteoblasts, hepatocytes, keratinocytes, mammary epithelial cells, B cells, macrophages, and ovarian luteal cells.(26,34,35) We predict that the generation of the specific MAbs 7H5 and 7D2 against C/EBPβ will elucidate additional functions of C/EBPβ.
Footnotes
Acknowledgments
We thank Dr. Yoshikazu Sado (Shigei Medical Institute, Okayama, Japan) for advice and Ms. Harumi Masuda for technical support. This work was supported by JSPS KAKENHI Grant Numbers 23310134, 25116010, 25118518, and 25132709 and Grant-in-Aid for JSPS fellows 25.4024.
Author Disclosure Statement
Taro Tachibana is a founder of Cell Engineering Corporation (Osaka, Japan).