
Abstract
Uncoordinated-5 homolog B receptor (UNC5B) was first found to mediate neural chemorepulsive effects by binding to its ligand netrin-1 in the nervous system. Newer evidence indicated that UNC5B also has functions outside the nervous system. In this study, we report on the generation of a monoclonal antibody specific to the outer-membrane immunoglobulin-like domains of UNC5B using the hybridoma technique. Western blot, immunofluorescence, and flow cytometry analyses showed that the antibody specifically bound to UNC5B protein. Interestingly, the antibody blocked the Netrin-1-induced inhibitory effect on the mobility of melanoma A375 cells by wound healing assay and transwell migration assay, whereas it had no effects on cell proliferation measured by CCK-8 assay. Thus, the functional antibody may provide a useful tool for the study of UNC5B expression profiles and functions outside the nervous system.
Introduction
U
In addition to modulating immune cell function, UNC5B may also participate in the regulation of tumor progression. In malignant melanoma, both netrin-1 and unc5b mRNAs were found to be upregulated. Reduction of netrin-1 expression by small interference RNA resulted in the reduction of melanoma mobility.(13) It has been considered that UNC5B belongs to the so-called dependence receptor family. UNC5B mediated P53-dependent apoptosis in the absence of netrin-1, but inhibited P53-dependent apoptosis when bound to its ligand netrin-1.(14,15) Recently, several groups reported that unc5b expression was downregulated in some cancers, such as colorectal and bladder cancer, and lower expression of unc5b in cancer tissue was correlated with high recurrence rates and poor prognosis.(16,17) The mechanisms of unc5b downregulated expression in these cancers are still not clear.
In this study, we have produced a monoclonal antibody, designated as 2C9, that binds specifically to UNC5B. The antibody regulates migration of A375 melanoma cells. Thus, this antibody can be used to study the UNC5B expression pattern and function in humans.
Materials and Methods
Materials
RPMI 1640, DMEM, and fetal bovine serum were purchased from Gibco BRL (Grand Island, NY). HAT medium and PEG solution were purchased from Sigma (St. Louis, MO). Ni2+ Sepharose column and rProtein G Sepharose 4B were from GE Healthcare (Uppsala, Sweden). BALB/c mice were kept at Soochow University of China. All cell lines (SP2/0, U87-MG, A375, and HL60) were obtained from ATCC (Manassas, VA) and cultured at 37°C in a humidified atmosphere of 5% CO2.
Expression of UNC5B fusion protein
The cDNA encoding the extracellular immunoglobulin domains of unc5b was amplified by reverse transcription-PCR using total RNA extracted from human endothelial cells. The primer sequences were 5'-C G
Immunization of mice
Three BALB/c mice (female, 8 weeks old) were chosen and each was subcutaneously injected with 100 μg purified fusion protein fully emulsified with Freund's complete adjuvant. Two weeks later, each mouse was boosted with 50 μg fusion in Freund's incomplete adjuvant. The booster injection was repeated every 2 weeks for two injection times. Finally, mice were injected intravenously with 50 μg UNC5B recombinant protein without Freund's adjuvant 4 days before cell fusion.
Establishment of hybridomas
Splenocytes of the immunized mice were fused with SP2/0 cells at a ratio of 10:1 in the presence of 50% polyethylene glycol (PEG1500). The fusion cells were cultured in HAT medium in 96-well plates for 7 to 10 days. All cell colonies were selected and their supernatants were detected for antibody against UNC5B using indirect ELISA. Positive clones were chosen and subcloned by the limiting dilution methods three times to establish stable hybridomas secreting monoclonal antibody.(18) The detail ELISA method was as follows:
Ten μg/mL purified fusion protein or pET-32a carrier protein in coating buffer (0.05 M bicarbonate, pH 9.6) were coated in the 96-well ELISA plates overnight at 4°C, respectively, then washed with PBS-T (0.1% Tween-20 in PBS) two times. The plates were blocked with PBS-BSA (2% BSA in PBS) at 37°C for 2 h. The supernatants of hybridoma cells were incubated in the two kinds of plates at 37°C for 1 h. After washing, HRP-labeled goat anti-mouse IgG (Bioworld, Minneapolis, MN) was added and incubated at 37°C for 1 h. Tetramethylbenzidine (TMB) was added and the reaction was stopped by adding 1 N H2SO4. The optical density (OD) was measured at 450 nm by a microplate reader (SpectraMax M5, Molecular Devices, Sunnyvale, CA).
MAb production
Stable hybridoma cells (2C9) were intraperitoneally injected into BALB/c mice (female, 8 weeks old) to produce a high-titer monoclonal antibody. Briefly, 0.5 mL sterile paraffin oil was injected into the peritoneum of the mouse 7 days before the hybridoma cell injection. Each mouse was injected with 2×106 cells. About 10 days later, ascites were collected, diluted with 5 volumes PBS, and centrifuged at 13,000 g for 30 min to obtain the supernatant. The supernatant was further purified by rProtein G Sepharose 4B column. The reactivity of the antibody was analyzed by ELISA as described above.
Western blot analysis and reverse transcriptase-polymerase chain reaction
HL60, U87-MG, and A375 cells were lysed in RIPA buffer (50 mM Tris-HCl, 1% Triton X-100, 10% Glycerol, 150 mM NaCl, and proteinase inhibitor cocktail) for 30 min on ice and centrifuged at 13,000 rpm for 15 min at 4°C to remove cell debris. The cell lysates with a total protein of 20 μg were loaded onto the SDS-PAGE gels and electrophoretically transferred to PVDF membranes by MiniProtean (Bio-Rad, Hercules, CA). After blocking with 5% skim milk in PBS for 2 h, the membranes were incubated with 2C9 (1 μg/mL) or rabbit anti-UNC5B polyclonal antibody (Abcam, Cambridge, MA) overnight at 4°C. The membranes were washed with PBS-T four times, and incubated with HRP-conjugated goat anti-mouse IgG (1:10,000 dilution) for 1 h at room temperature. Proteins of interest were detected by enhanced chemiluminescence (ECL, Biological Industries, Kibbutz Beit Haemek, Israel) in a Kodak Image Station GAP-201.
RNA from the three cells was isolated by centrifugation method.(19) RNA of 2 μg was used for reverse transcriptase-polymerase chain reaction (RT-PCR) using the RevertAid™ First Strand cDNA Synthesis Kit (Thermo Scientific, Illkirch Cedex, France). Semi-quantitative RT-PCR (30 cycles) was employed to analyze unc5b gene expression using the gene-specific forward and reverse primers: 5'GAGCCGAAACCGCTAATG3' and 5'CTGCCACTCCAAATGTGATAGA3'. The expression level of β-actin was used as the internal control.
Immunofluorescence and flow cytometry analysis
Immunofluorescence
Cells (5×103) were inoculated in 96-well plates the day before and fixed with 4% paraformaldehyde at 4°C for 20 min. The wells were washed with PBS three times and incubated with the 2C9 (2 μg/mL) or rabbit anti-UNC5B polyclonal antibody at 37°C for 1 h. After washing, the cells were labeled with Alexa Fluor 488-conjugated goat anti-mouse IgG or Alexa Fluor 594-conjugated goat anti-rabbit IgG antibody using 1:500 dilution (Invitrogen, Eugene, OR). Nuclei were stained with DAPI. The fluorescent images were captured by inverted microscope (DIML, Leica, Wetzlar, Germany).
Flow cytometry
A375 cells (3×106) were trypsinized, washed with PBS, and then incubated with 2C9 (2 μg/mL) or mouse IgG (2 μg/mL) at room temperature for 1 h. Cells were washed and labeled with Alexa Fluor 488-conjugated goat anti-mouse IgG antibody. Finally, the cells were resuspended in 500 μL PBS and measured by flow cytometry (BD FACS Calibur, San Diego, CA).
Wound-healing assay
Wound-healing assay was performed in 24-well plates.(20) A375 cells (2×105 in 500 μL medium) were inoculated in 24-well plates. Three parallel artificial scratches using a 200 μL pipette tip (Axygen, Union City, CA) were made for each well after complete cell confluence. Scratched wells were washed with PBS three times and 500 μL complete medium was added containing rhNetrin-1 or 2C9 antibody. The cells were then incubated in 37°C, 5% CO2. After 24 and 48 h, three photographs of each scratch were taken by inverted microscope and the scratch space was measured by Leica application suite. The wound healing length was the result of the space at 0 h minus that at a different interval, and the average was calculated with the same treatment.
Transwell migration assay
The effect of 2C9 antibody on migration of A375 cells was further analyzed using transwell chambers with 8-μm porous membrane (Corning, Corning, NY).(21) Cells (1.5×105) were loaded into the migration chamber with or without 2C9 antibody. Medium containing rhNetrin-1 at 20 ng/mL was placed in some of the lower chambers. After allowing cell migration for 16 h, cells were removed from the upper side of the membrane; migratory cells on the lower side of the membrane were fixed with ice-cold methanol and stained with hematoxylin and eosin. The number of migratory cells was counted by fluorescence microscopy (400× magnification). Each experiment was performed in triplicate.
Cell proliferation assay
Cell proliferation was measured by Cell Counting Kit-8 (Beyotime, Haimen, Jiangsu, China) as described.(22) A375 cells were seeded in 96-well plates at a density of 4000 cells per well, and incubated overnight to make sure they were adhesive. Recombinant netrin-1 and 2C9 antibody were added into the cultured medium and incubated for another 24 or 48 h. CCK-8 solution (20 μL) was added into each well, followed by incubation for 3 h at 37°C. The absorbance at 450 nm was measured. Columns represented the mean percentage of OD values relative to the negative control. All measurements were made in triplicate and repeated in at least three separate experiments.
Statistical analysis
Results are expressed as mean±SD (range) or percent. The difference between two groups was analyzed using the Student's paired t-test. A p value <0.05 was considered statistically significant. All calculations were performed using the GraphPad Prism software (GraphPad Software, San Diego, CA).
Results
Recombinant UNC5B fusion protein was expressed in E. coli and purified
To obtain protein as immunogen, the cDNA encoding the two immunoglobulin domains in UNC5B N-terminal region was cloned into pET32a vector. His-tagged fusion protein was induced in BL21 (DE3) bacteria with 1 mM IPTG. SDS-PAGE and Coomassie blue staining showed that the protein was successfully expressed at a molecular weight of about 50 kDa and mainly in soluble fraction. The fusion protein in the soluble fraction was purified by Ni2+ Sepharose column (Fig. 1).
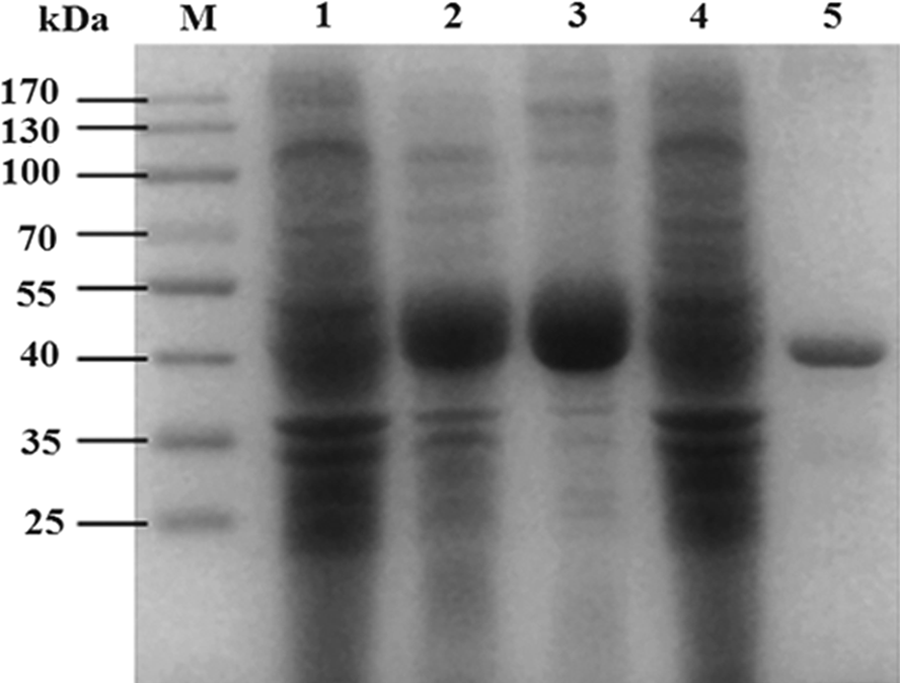
SDS-PAGE analysis of the UNC5B protein fragment expressed in BL21 (DE3). M, protein marker; lane 1, total lysate without IPTG induction; lane 2, total lysate after 1 mM IPTG induction; lanes 3, 4, soluble supernatant and insoluble fraction after 1 mM IPTG induction individually; lane 5, purified recombinant protein.
Generation of hybridomas secreting MAb against recombinant UNC5B
Monoclonal antibody in culture supernatants of hybridoma cells was detected by indirect ELISA. After three rounds of subcloning and detecting, a stable positive hybridoma clone, designated as 2C9, was chosen to produce ascites. The monoclonal antibody in ascites was purified with rProtein G Sepharose and stored at a final concentration of 1 mg/mL. The purified 2C9 antibody was further identified by the specificity by ELISA. ELISA results showed that the 2C9 antibody bound to recombinant UNC5B fragment, while mouse IgG could not. The titer of 2C9 against UNC5B was measured by indirect ELISA to be 1:128,000 (Fig. 2).
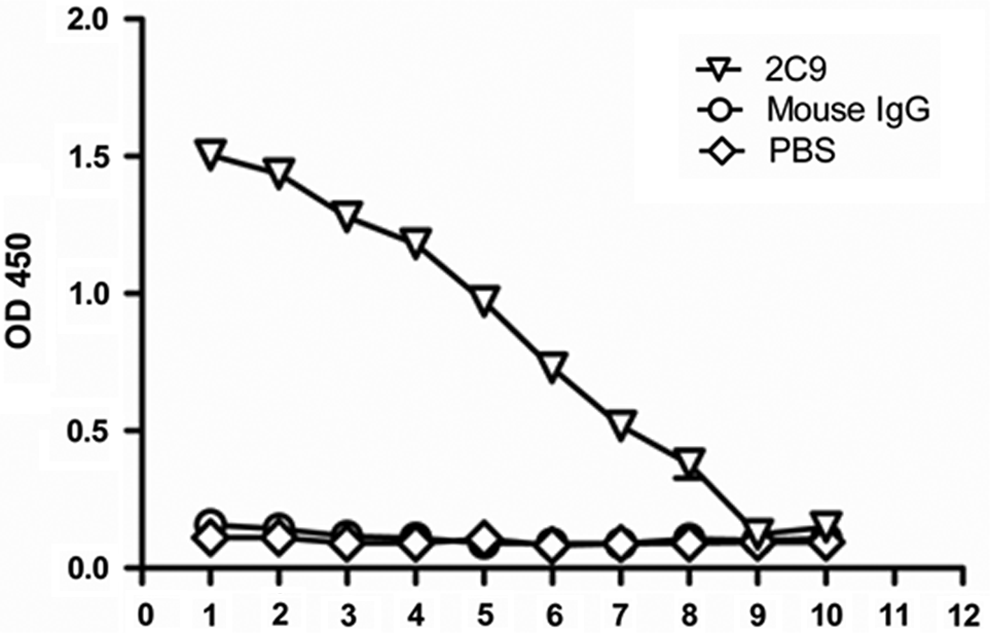
Titer of anti-UNC5B monoclonal antibody 2C9 was detected by indirect ELISA. 2C9 (1 mg/mL) monoclonal antibody was diluted sequentially. 1, 1:1000; 2, 1:2000; 3, 1:4000; 4, 1:8000; 5, 1:16,000; 6, 1:32,000; 7, 1:64,000; 8, 1:128,000; 9, 1:256,000; 10, 1:512,000.
Western blot analysis
To analyze whether 2C9 could bind native UNC5B in cells, U87-MG, A375, and HL-60 cell lysates were prepared for Western blot analysis. Results showed that 2C9 specifically recognized a protein with a molecular weight of about 104 kDa in U87-MG and A375 cells, but not in HL60 cells (Fig. 3). The result is consistent with the mRNA expression level in these cell lines. 2C9 also recognized the purified recombinant UNC5B protein fragment expressed in E. coli. The commercial anti-UNC5B polyclonal antibody also recognized the 104 kDa band in U87-MG and A375 cells, but not in HL60 cells. Because the epitopes of the polyclonal antibody were in the cytoplasmic domain of UNC5B, the antibody did not recognize the recombinant N-terminal UNC5B fragment. Based on the expression pattern and the fact that the detected band was the same as the predicted molecular weight, 2C9 was expected to specifically recognize UNC5B.
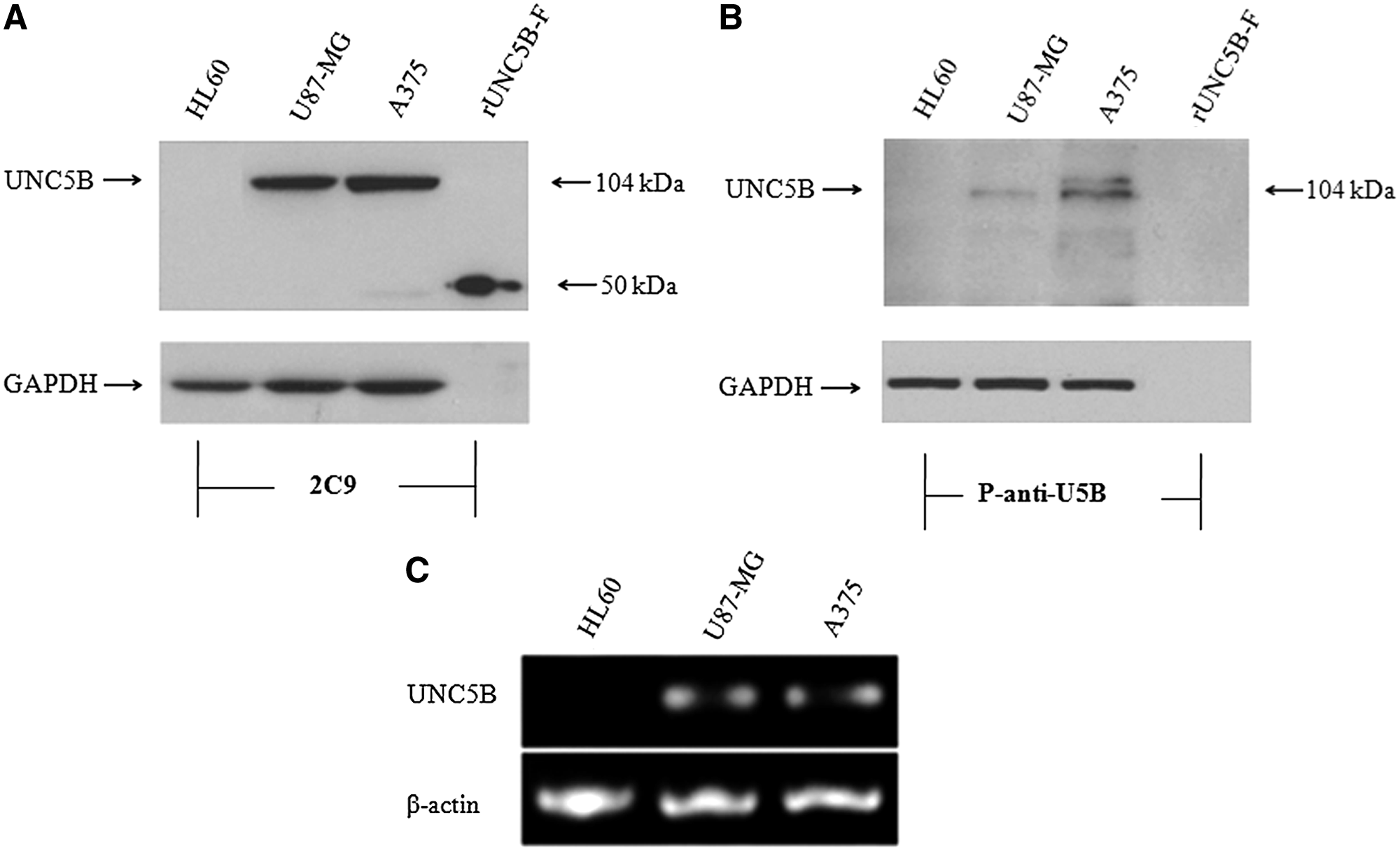
2C9 recognized UNC5B protein by Western blot analysis. (
Immunofluorescence and flow cytometry analysis
To further explore the binding specificity of 2C9 antibody with UNC5B, immunofluorescence and flow cytometry analysis were taken. Results of the immunofluorescence showed positive green fluorescence in A375 cells labeled with 2C9 (Fig. 4A, upper panel). When labeled with p-anti-U5B, A375 cells showed positive red fluorescence (Fig. 4A, lower panel). Flow cytometry results also showed positive binding of 2C9 antibody on A375 cells (Fig. 4B). These results suggest that 2C9 antibody recognizes UNC5B native protein.
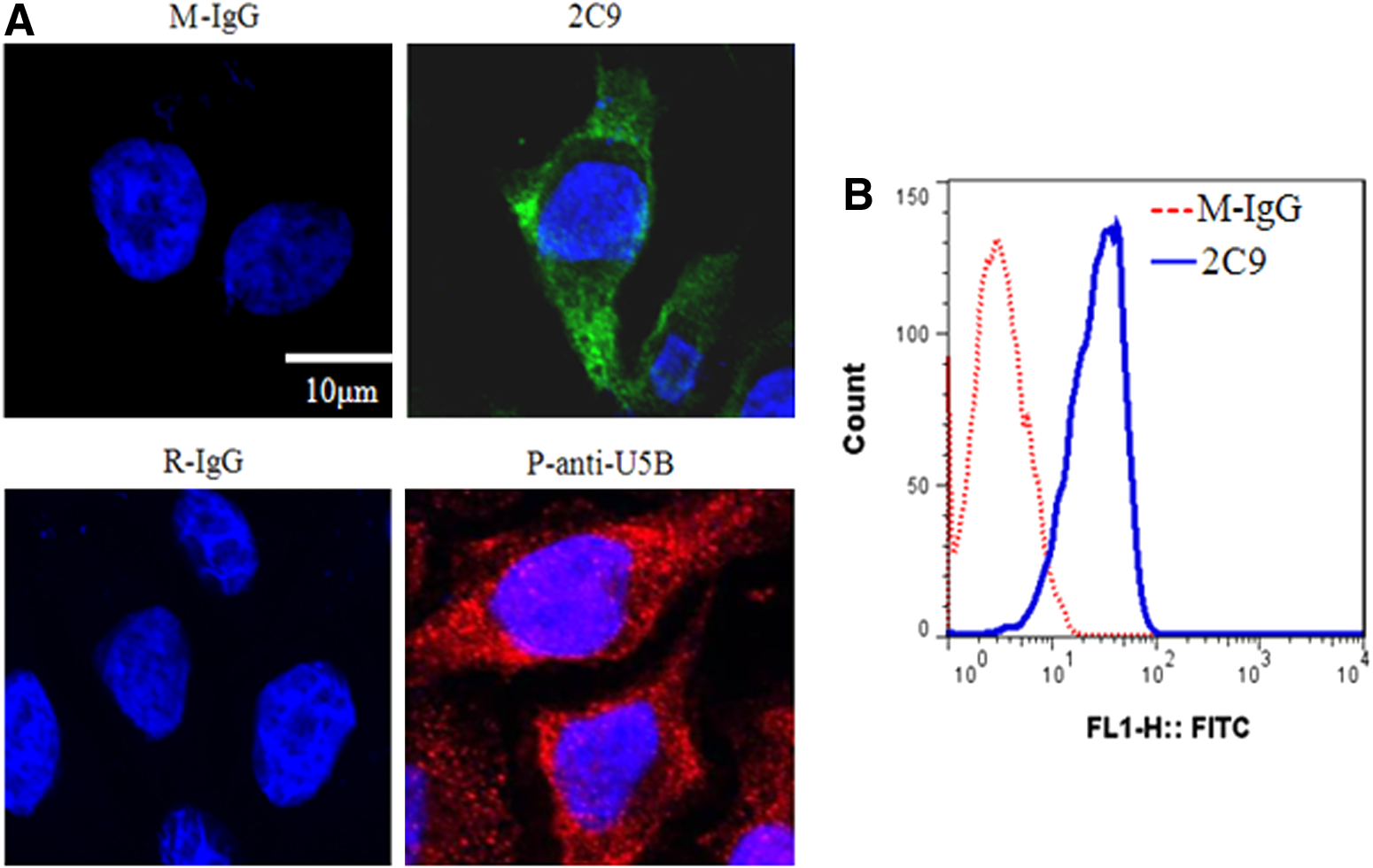
2C9 antibody binds UNC5B protein in cells. (
Effect of 2C9 on A375 cell migration
The potential function of 2C9 on tumor cell migration was characterized by scratch wound-healing assay. A375 cells were treated with netrin-1 and 2C9 antibody. The images were taken using an inverted microscope at intervals of 24 h, and representative fields are shown in Figure 5A. The results showed that netrin-1 treatment slowed the migration of A375 cells, but 2C9 antibody reversed the netrin-1-induced inhibitory effect on migration of A375 melanoma cells. Results from the transwell migration assay also suggest that 2C9 blocks the netrin-1-induced inhibitory effect on migration of A375 cells (Fig. 6).
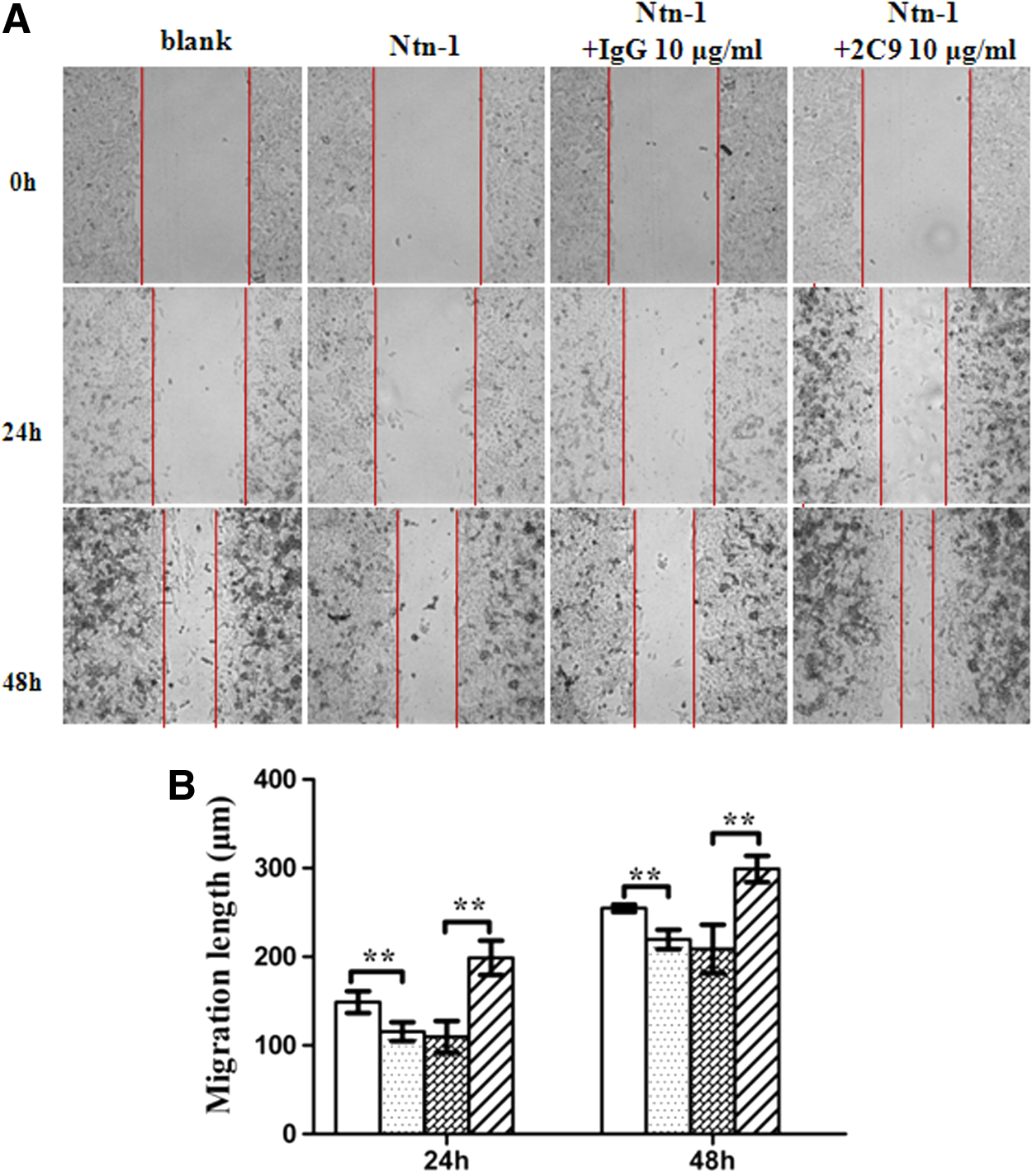
Role of 2C9 in wound healing of A375 cells. (

The role of 2C9 in the transwell migration of A375 cells. (
2C9 has no influence on A375 cell proliferation
To examine the involvement of 2C9 on cell proliferation, a proliferation assay was taken using a CCK-8 kit. The 2C9 antibody was added into A375 culture medium and incubated for different time lengths. Mouse IgG (10 μg /mL) was the negative control. Data showed that the proliferation rates of A375 cells were not different after treatment of netrin-1 and the 2C9 antibody compared to that of the blank control (100% proliferation, Fig. 7).

Effect of 2C9 on A375 cell proliferation. A375 cells were cultured with recombinant netrin-1 (Ntn) and 2C9 antibody for 24 or 48 h. Cell proliferation was measured by CCK-8 assay. Mouse IgG was negative control. Data represented as mean±SD from three independent experiments. p>0.05.
Discussion
Axon guidance molecules were first described in the nervous system to control the outgrowth direction of neuron cells. However, as an important guidance receptor, UNC5B is widely expressed outside the nervous system, where it controls cell migration, tissue development, and the vascular network establishment. In addition, it is involved in cancer development, tumor angiogenesis, and metastasis. In this study, we successfully expressed the N-terminal fragment containing two immunoglobulin domains of UNC5B in E. coli, and used it as immunogen to make hybridomas producing monoclonal antibody. After several rounds of screening, we obtained a stable hybridoma cell clone 2C9. The monoclonal antibody produced by this clone has shown UNC5B specific by Western blot, immunofluorescence, and flow cytometry.
UNC5B is a dependent receptor involved in the metastasis of tumor cells. It has been reported that netrin-1 can regulate melanoma cell mobility. Reduction of migration was seen in netrin-1 siRNA transfected melanocytes.(13) However, whether this phenomenon is directly mediated by UNC5B is not clear. In our study, we examined the UNC5B expression at both mRNA and protein levels and found that UNC5B was expressed in melanoma A375 cells. Inconsistent with previous reports,(13) our data demonstrated that netrin-1 inhibited A375 cell migration. This result may be because different melanoma cell lines were used and different netrin-1 receptors existed on different melanoma cell lines. More interestingly, the results of cell migration assay showed that the UNC5B monoclonal antibody 2C9 blocked netrin-1-induced inhibitory effect on the migration of A375 melanoma cells. This indicated that UNC5B is an important mediator in melanoma cell migration. The effect of 2C9 on the migration of A375 cells may be the result of interfering UNC5B signal transduction through blocking netrin-1–UNC5B interaction.
Moore and colleagues reported that netrin-1 promotes atherosclerosis via its receptor UNC5B by inhibiting the emigration of macrophages from plaque.(10) Blocking netrin-1–UNC5B interaction may inhibit atherosclerosis progress. Our prepared 2C9 antibody may be useful in such intervention. Potential application of 2C9 antibody in atherosclerosis treatment will need to be studied further.
UNC5B is considered to be a dependent receptor because cell proliferation and apoptosis are regulated in the presence or absence of its ligand. However, in this study, no significant difference of cell proliferation was found in A375 cells treated with netrin-1 or UNC5B blocking antibody 2C9. This result is consistent with that found by Kaufmann and colleagues.(13) They did not observe an effect on proliferation or apoptosis after netrin-1 treatment in melanocytic cells or netrin-1 siRNA knockdown in melanoma cells. This indicates that cell types reacted differently to netrin-1.
UNC5B deregulation occurs in multiple cancers, including colorectal, breast, ovary, uterus, stomach, lung, and kidney.(23) Low unc5b expression is even an independent risk factor for colorectal and bladder cancer patients.(16,17) Colorectal cancer patients with high expression of unc5b in stages II and III had a relatively low recurrence rate and good prognosis compared to those with low unc5b expression. Therefore, monoclonal antibody 2C9 may be used to detect the expression level of UNC5B in colorectal and bladder cancers for diagnosis and prognosis assessment.
In summary, a novel high-affinity and specificity monoclonal antibody against UNC5B was developed. It may be useful in the intervention of atherosclerosis and in the diagnosis and prognosis prediction of colorectal and bladder cancer.
Footnotes
Acknowledgments
This work was supported by grants from the Priority Academic Program Development of Jiangsu Higher Education Institutions of China and the National Natural Science Foundation of China (no. 81270593).
Author Disclosure Statement
The authors have no financial interests to disclose.