
Abstract
Recombinant antibody technologies are rapidly becoming available and showing considerable clinical success. However, the immunogenicity of murine-derived monoclonal antibodies is restrictive in cancer immunotherapy. Humanized antibodies can overcome these problems and are considered to be a promising alternative therapeutic agent. There are several approaches for antibody humanization. In this article we review various methods used in the antibody humanization process.
Introduction
R
Structural Features of Antibodies
Immunoglobulin molecules are composed of four polypeptide chains (two identical H and L chains).(15) Each H chain has one VH domain and C domain counted from the amino terminal. The L chain has one VL domain at the amino terminus and only one C domain.(16) Each V domain contains three hypervariable loops that are involved in the formation of the antigen binding site, referred to as complementarity-determining regions (CDRs).(17–19) Chothia and co-workers have shown that five hypervariable regions usually have one of a small number of main chain particular conformations; these have been called canonical structures.(20–22) These structures are determined by the length of the loop and the amino acids present at key sites. These key residues have been discussed by many researchers: Tomlinson and colleagues reported on the Vκ canonical structures; Chothia and Lesk and Wu and Cygler on those in VL; and Chothia and colleagues on those in VH.(23–26) For the CDR3 of heavy chain, having the most variable CDRs in length and amino acid composition, it has been more difficult to predict its canonical structure. The CDR-H3 loop conformation is modeled using CONGEN, WAM, and Modeller,(27) and figure representations were generated with Swiss PDB Viewer.(28) CDRs are supported within the variable domains by more conserved framework regions. Many authors have elegantly demonstrated that framework residues can also make contact with antigen.(29,30)
Humanization by CDR-grafting method
The first humanized antibodies were constructed by transferring murine CDRs to a closely related framework (acceptor) derived from known human antibodies using a convenient approach called CDR grafting (Fig. 1).(31,32) In CDR grafting, it is common to select a suitable human acceptor framework. Krawczyk and colleagues grafted the murine mA2c CDRs into appropriate human germline acceptor framework regions to develop a humanized antibody with the lowest possible immunogenicity.(33) Couto and colleagues used a novel positional consensus sequence for humanization of anti-BA46 monoclonal antibody.(34) Hu and colleagues selected human germline antibody genes for humanization of anti-Ricin neutralization monoclonal antibody.(35) Bowers and colleagues performed humanization of hβNGF antibody that recognizes the cytokine hβNGF by grafting of CDR3 region into a non-homologous human germline variable heavy chain to minimize the immunogenic response in humans. The variable domains of the CDR3-grafted heavy and light chains were fused together, displayed on the surface of HEK293 cell lines, and matured using in vitro somatic hypermutation. This approach could be an efficient method to generate potent humanized antibodies containing a minimal number of murine residues in the variable regions.(36) Feldmann and co-workers established the humanization of anti-CD3–anti-PSCA (the prostate stem cell antigen) bi-specific single chain antibody by CDR-grafting method. The humanized bi-specific antibody could activate both CD8+ and CD4+ T cells and mediate efficient lysis of tumor cells.(37)
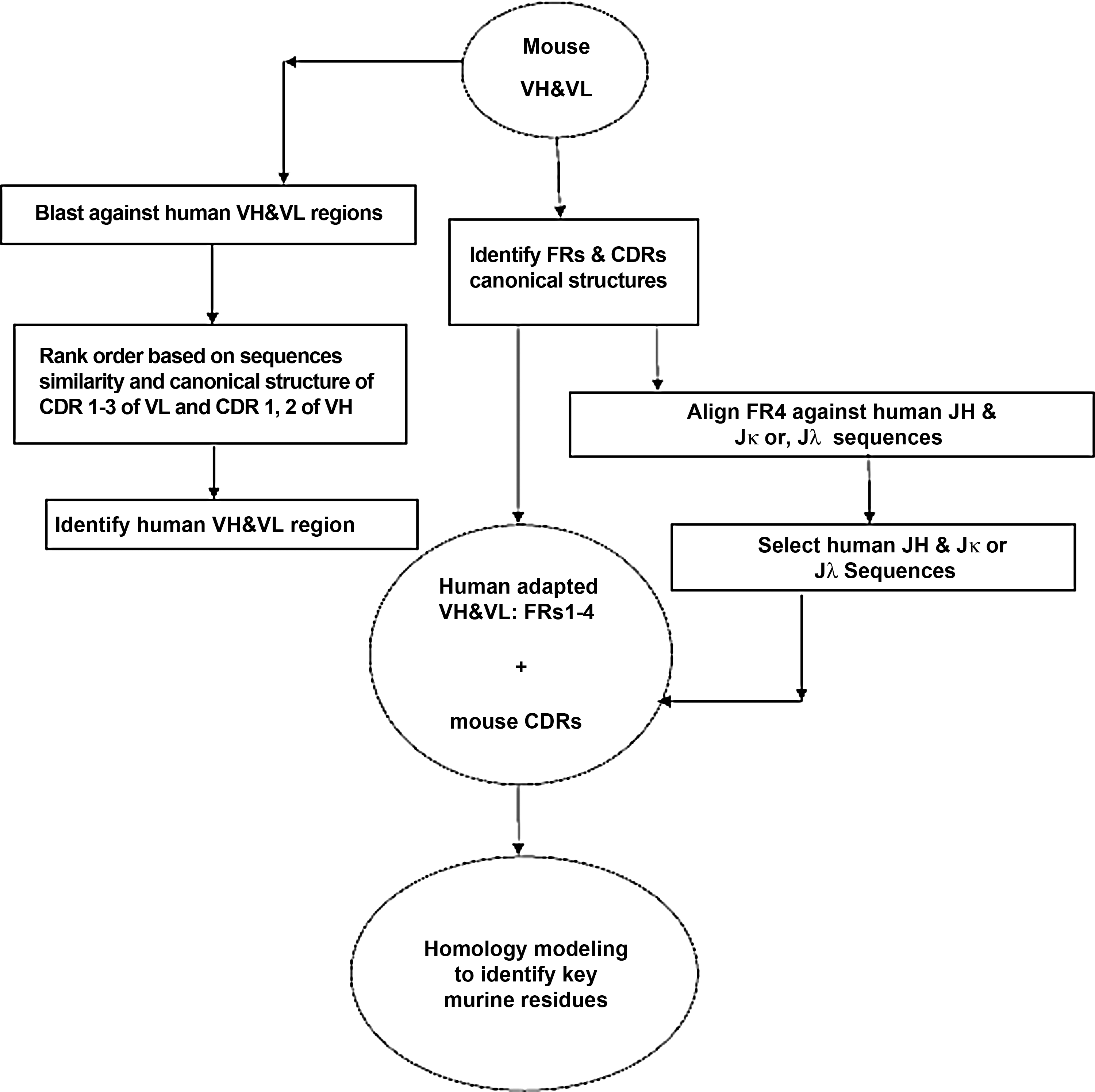
Flowchart of antibody humanization using CDR-grafting method.
Back Mutation of Key Murine Residues
The CDR-grafting method often results in reduced affinity or specificity for the target antigen.(38–40) Foote and Winter estimated that some key murine framework residues play a critical role in adjustment of the CDR structures, referred to as the Vernier zone, preserved in humanized MAbs to restore the binding affinity.(41) Notably, Makabe and colleagues have also indicated that these residues make enthalpic contributions to antigen binding during antibody humanization.(42) Chothia and colleagues identified the regions at the VH/VL interface and also noted residues involved in CDR loop conformations, which are required for affinity restoring.(43) Tiwari and colleagues reported the construction of a humanized high affinity anti-HBs antibody by grafting its antigen binding site onto a human consensus framework sequence with the highest homology. The humanized scFv retains the binding characteristic by using molecular modeling to alter not only the clearly permissible amino acids but also several minimal positional template and VH/VL interface residues.(44)
Remy and co-workers described humanization of WO-2 by grafting the murine antibody WO-2 CDRs onto the human germline sequences while preserving the Vernier zone and the resulting high-resolution X-ray crystal structure. The humanized WO-2 fragments (Fab and scFvs) restore high affinity for the antibody peptide and therefore show a molecular basis for passive immunotherapy of Alzheimer's disease.(45) Wu and colleagues selected consensus sequences of the light chain subgroup kappa I and heavy chain subgroup III, respectively, to humanize an anti-CD20 monoclonal antibody 8E4 using CDR-grafting method. They built the molecular model of the antibody to help identify structurally key framework residues in the humanized version that must be back-mutated to reconstitute the full binding activity. The humanized antibody Hu8E4 exhibited much more potent complement-dependent cytotoxicity than rituximab, while its ability in ADCC and inducing apoptosis in B lymphoma cells was similar to that of rituximab.(46)
Humanization by Resurfacing Approach
Humanization via variable domain resurfacing is another approach that emerged in the early 1990s(47); the initial antibody resurfacing technique was described by Pedersen and colleagues.(48) This method was based on identification of the accessible and protruding residues in the nonhuman antibody that need to be changed to human sequences without influencing the conformation of the CDR loops to produce humanized antibodies with reduced immunogenicity.(48–50) Fontayne and co-workers constructed a fully active humanized Fab fragment of the monoclonal antibody 6B4 against human glycoprotein IBα in comparison to the original murine antibody using variable domain resurfacing guided by computer modeling.(51) Staelens and colleagues humanized murine MAb 82D6A3, directed against the von Willebrand factor (VWF) A3-domain, via resurfacing strategy. The variable domains of humanized antibody were then grafted on the respective constant regions of a human IgG4. The resultant antibody h82D6A3 had a longer half-life, and reduced immunogenicity and bio-activity inhibition of VWF interaction to fibrillar collagen in vitro comparable to that of the murine IgG.(52) Zhang and colleagues resurfaced murine monoclonal antibody Z12 against human tumor necrosis factor-alpha (hTNF-alpha). The humanized Fab antibody version showed the same antigen binding affinity when it was compared to the original murine monoclonal antibody.(53) Chiu and associates generated the humanized murine anti-human TNF-α monoclonal antibody m357 using the following three steps: first, the molecular models of variable domains of m357; second, calculation of the solvent accessible residues for identifying the non-human-like framework surface residues using AREAIMOL program; and finally, mutation of these surface residues to the human counterparts. The results indicated that the humanized antibody h357 retained in vitro bioactivities was similar to the original antibody.(54) Zhu and colleagues fabricated a resurfaced humanized version (HAb18-huscFv) of HAb18scFv and then fused it to the human IgG1Fc fragment to form (HAb18-huscFv)(2)-Fc. The resultant antibody fragment could be a more efficient antibody with high-affinity targeted cytotoxicity and decreased immunogenicity compared to the parental antibody.(55)
Humanization of Antibodies by SDR-grafting Method
CDR-grafted humanized antibodies may still elicit an immune anti-idiotypic (anti-Id) response in patients. To minimize the anti-V region responses, the antibody can be humanized by grafting only the specificity-determining residues (SDRs) onto the human frameworks.(56,57) Padlan and colleagues identified the most variable positions within each CDR that are directly involved in interaction with the antigen.(47) Gonzales and colleagues used three various human germline V-kappa acceptor sequences and another human template for SDR grafting of the murine anti-carcinoembryonic antigen (CEA) MAb COL-1. Results showed that the binding affinity of SDR-grafted COL-1 (HuCOL-1SDR) are comparable to its corresponding CDR-grafted versions (HuCOL-1). The SDR-grafted humanized antibody HuCOL-1 compared to HuCOL-1 and the “abbreviated” CDR-grafted antibody exhibited lower reactivity in patients' sera carrying anti-V region antibodies to COL-1 while retaining its antigen-binding property of parental antibody.(58) Tamura and colleagues humanized anti-tumor-associated glycoprotein-72 MAb CC49 by SDR-grafting method. The resulting humanized CC49 antibody retained the original affinity of the murine antibody, while reactivity with the sera of patients was only minimal.(59) Yoon and associates humanized the anti-TAG-72 by grafting only SDR regions onto homologous human immunoglobulin germline segments with CDRs of the same canonical structures as those of the murine antibody while retaining two key murine framework residues. The humanized antibody AKA exhibited about two-fold less affinity compared with the parental antibody CC49 and 27-fold less sera reactivity compared with the CDR-grafted humanized antibody HuCC49.(60)
Superhumanized Antibodies
Tan and colleagues proposed a superhumanization method based on structural homologies between mouse and human CDRs in which a framework homology is irrelevant. Using this approach, they superhumanized the murine anti-human CD28 antibody 9.3. The resultant humanized antibody lost 30-fold affinity but retained biological activity.(61) Hwang and colleagues generated the superhumanized form of the antibody D1.3 with a better affinity (6-fold loss in comparison to the murine antibody) than that with the CDR-grafted variant (70-fold loss in affinity).(62) In another example reported by Mader and colleagues, a superhumanized type of chimeric antibody Ab2/3H6 lost its original binding affinity.(63) Hu and colleagues humanized variable domains of 1A4A1, a neutralizing antibody against the Venezuelan equine encephalitis virus (VEEV), by grafting SDR regions with highest similarities to murine antibody onto the human germline frameworks. The superhumanized antibody was demonstrated to retain antigen-binding specificity and neutralizing activity.(64)
Humanization Based on Human String Content Optimization
Lazar and colleagues introduced a new strategy for antibody humanization based on an immunologically relevant metric of antibody humanness termed human string content (HSC). The scores of HSC are significantly different across the framework and CDR regions of the mouse amino acids with distinct human germline sequences. The humanization of target sequence is by maximizing this quantity rather than using a global identity measure to generate multiple diverse humanized variants. The resulting variable domains are less immunogenic and showed increased binding affinities comparable to those humanized by the standard CDR-grafting method.(65) Hammond and colleagues humanized the chimeric anti-CD30 antibody cAC10 using the HSC approach. The novel Xmab2513 antibody has a significant improvement in binding affinity for FcγRIIIA receptor and CD30 antigen (4-fold higher than that of the parental antibody) and retains the potent anti-proliferative effect exhibited by the cAC10 and 5F11 antibodies.(66,67)
Other Antibody Humanization Methods
Guided selection developed by Jespers and colleagues provides a powerful tool for humanization that minimizes the immunogenic response in humans. In this approach, mouse VH and VL domains are sequentially or in parallel replaced by human variable regions, respectively, using the power of the phage-displayed antibody library. This technology is based on panning and screening of genes encoding recombinant antibodies against antigens using repeated cycles of phage capture and elution from immobilized ligand followed by phage amplification in prokaryotic or eukaryotic cells.(68) Osbourn and colleagues applied guided selection for the humanization of the mouse MAb32 against TNF-alpha antigen. The mouse VH region was combined with VL repertoire-derived human B cells. The human VL-shuffled antibody library was displayed on the phage surface, then selected against target antigen. The results demonstrated that human Fab fragments recognized the same epitope and had a binding affinity similar to the original mouse antibody.(69) Zhang generated the humanized single chain antibody from a phage-displayed library with potent neutralizing activity for high brain natriuretic peptide (BNP) level that correlated well with cerebral salt wasting syndrome (CSW).(70) Zhang and colleagues produced a human anti-γ-seminoprotein Fab antibody through guided selection approach. The resulting antibody fragment recognized the same epitope with similar affinity to the parental antibody.(71) Kim and Hong humanized mouse CC49 antibody against tumor-associated glycoprotein TAG-72 using guided selection strategy. The Fabs with the selected human light chains showed similar specificity of CC49 antibody and better antigen-binding affinities compared to the murine antibody.(72)
Dall'Acqua and co-workers reported the humanization of a mouse MAbB233, a neutralizing antibody against the cancer target receptor tyrosine kinase EphAa2 using a framework shuffling method. The six CDRs of B233 were fused into a library of a human germline framework gene. The resulting Fab combinatorial libraries were screened for binding to target antigen in a two-step selection process. This humanized IgGs possessed improved binding affinity and thermal stability.(73) Liu and colleagues used two sequential phage displays with a predetermined CDR3 gene to humanize the light chain and Fd fragment of the heavy chain of a mouse MAbE10 against human integrin αvβ3, respectively. This successful production of a humanized scFv indicated that it retained a high binding activity to target antigen, had the same specificity as the original MAb, and acted to inhibit αvβ3-mediated cancer cell growth in vitro and in vivo.(74) Zheng and co-workers humanized the murine 13D4 antibody that recognizes the hemagglutinin using CDR grafting and combinatorial framework library screening to identify the key residues that should be replaced with the murine sequence in the humanized framework. Humanized versions of 13D4 antibodies maintain high-affinity binding for pathogenic H5 avian influenza virus compared to the original murine antibody.(75)
Conclusion
Recombinant antibodies have become a major group of therapeutic classes.(76) However murine monoclonal antibodies may induce immunogenic responses, potentially limiting their application for humans. Humanization is aimed to diminish or prevent such undesirable properties of these antibodies.(77,78) Various strategies have been developed to humanize murine MAbs, among which CDR-grafting method has become the most common. This method is based on the transfer of murine CDRs onto the human acceptor frameworks.(79) It has been reported that the transfer of murine CDRs onto human frameworks results in reduction of the parental binding affinity.(80) Thus back mutations have to be introduced in the humanized version to restore binding affinity. Amino acid residues at these key positions have been demonstrated to affect the CDR conformation and therefore influence the antibody affinity.(81,82) This review briefly updates the different strategies for antibody humanization that have been attempted to increase their therapeutic efficiencies.
Footnotes
Author Disclosure Statements
There is no financial support for this study. The authors thank Dr. Kamal Veisi, Dr. Yaghoub Safdari, Dr. Leila Rahbarnia and Shiva Ahdi for editing this article.