
Abstract
Most of the functions of D-amino acids (D-AA) remain unclear because of little analytic methods for specific detection/determination. In this study, a highly specific monoclonal antibody to D-glutamic acid (D-Glu-MAb) was produced using a hybridoma method. Characterization of D-Glu-MAb by indirect enzyme-linked immunosorbent assay (ELISA) revealed that it has high selectivity against D-Glu-glutaraldehyde (GA) conjugates, while no cross-reaction was observed when 38 other kinds of AA-GA conjugates were used. Moreover, subsequent indirect competitive ELISA disclosed that an epitope of D-Glu-MAb is a D-Glu-GA molecule in the conjugates, suggesting that D-Glu-MAb could be a useful tool to investigate the functional analysis of D-Glu in immunostaining.
Introduction
A

Structures of (
In this study, therefore, MAb against D-Glu (D-Glu-MAb) was produced by the hybridization of splenocytes obtained from D-Glu-bovine serum albumin (BSA) conjugates, immunized BALB/c mice, and the mice myeloma cell line SP2/0. Their characterization based on an enzyme-linked immunosorbent assay (ELISA) revealed that D-Glu-MAb has high selectivity against D-Glu-GA conjugates, while there was no cross-reactivity against another 38 AA-GA conjugates. Moreover, the D-Glu-GA molecule was found to be an epitope of D-Glu-MAb. These results demonstrate that the D-Glu-MAb could be a promising tool to histopathologically elucidate its function and mechanism, which are related to D-Glu. Herein, the production and characterization of D-Glu-MAb are demonstrated.
Materials and Methods
Chemicals and reagents
Amino acids used in this study were obtained from Nacalai Tesque (Kyoto, Japan). Bovine serum albumin (BSA, ≥98%) and human serum albumin (HSA, ≥99%) were provided from Sigma-Aldrich (St. Louis, MO). Freund's complete and incomplete adjuvants were obtained from Difco (Detroit, MI). Glutaraldehyde (GA, 25%), formaldehyde (FA, 37%), valeraldehyde (VA), and glutaric acid, which were used as free antigen, were purchased from Nacalai Tesque. Peroxidase-labeled anti-mouse IgG was purchased from Organon Teknika Cappel Products (West Chester, PA). All other chemicals were standard commercial products of analytical reagent grade.
Animals
Five-week-old male BALB/c mice were obtained from KBT Oriental Co. (Saga, Japan). They had free access to a standard diet (MF; Oriental Yeast Co., Tokyo, Japan) and water. All the procedures and animal care were approved by the Committee on the Ethics of Animal Experiments, Graduate School of Pharmaceutical Sciences, Kyushu University, and were conducted according to the Guidelines for Animal Experiments of the Graduate School of Pharmaceutical Sciences, Kyushu University (approval no: A25-018-0).
Synthesis of D-Glu-GA-BSA conjugates for immunization
Synthesis of D-Glu-GA-BSA conjugates was done by crosslinking of D-Glu to BSA through glutaraldehyde (GA). BSA (12 mg) was dissolved in 0.1 M sodium-phosphate buffer (pH 7.0) containing 65 mM of D-Glu (1.6 mL). 2.5% aqueous GA solution (0.4 mL) was then added to the mixture, and stirred at room temperature for 2 h. Subsequently an aqueous sodium borohydride (NaBH4) solution (4 mg/mL, 2 mL) was added and stirred at room temperature for 1 h. The reaction mixture was dialyzed five times against distilled water at 4°C and then lyophilized to obtain D-Glu-GA-BSA conjugates (10.3 mg). They were kept at −20°C until use. In addition, all of the D-/L-AA-GA-HSA conjugates used as coated antigen were synthesized in the same manner using the HSA.
Determination of hapten number of D-Glu-GA-BSA conjugates by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry
The hapten number of the D-Glu-GA-BSA conjugates was determined by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS), as previously described.(15) A small amount (1–10 pmol) of an antigen conjugate was mixed with a 103-fold molar excess of sinapinic acid in an aqueous solution containing 0.15% (v/v) trifluoroacetic acid and acetonitrile in the ratio of 2 to 1. The mixture (2 μL) was spotted onto the stainless steel target, dried completely at room temperature, and subjected to a MALDI-TOF-MS spectrometer (Autoflex III, Bruker Daltonics, Bremen, Germany) irradiated with a nitrogen laser (337 nm, 200 Hz max. firing rate). The spectra were recorded in linear positive high mass mode with a mass range of 10,000–100,000 Da, and analyzed by using flexControl software (Bruker Daltonics).
Immunization, hybridization, and purification of monoclonal antibody
Production of MAb against D-Glu (D-Glu-MAb) was carried out by immunization of D-Glu-GA-BSA conjugates into 5-week-old BALB/c male mice as described in our previous study(16) with slight modification. The immunization was performed four times every 2 weeks. The first immunization (50 μg as D-Glu-GA-BSA conjugates) was administered into the abdominal cavity of the BALB/c mice as a 1:1 emulsion in Freund's complete adjuvant. The second immunization (50 μg D-Glu-GA-BSA conjugates) was administered as a 1:1 emulsion in Freund's incomplete adjuvant. On the third day after the final booster (100 μg as D-Glu-GA-BSA conjugates), the spleen was taken out and splenocytes were isolated to fuse with a mouse myeloma cell line, SP2/0, using the polyethylene glycol. The resultant hybridomas were selected by hypoxanthine-aminopterin-thymidine medium. Hybridomas producing MAb reactive to D-Glu-GA-HSA conjugates were screened by indirect enzyme-linked immunosorbent assay (ELISA) and cloned by the limited dilution method. Selected hybridomas were cultured in enriched RPMI1640-Dulbecco's-Ham's F12 (eRDF; Kyokuto Pharmaceutical Industrial, Tokyo, Japan) medium supplemented with RD-1 additives containing 5 μg/mL of insulin, 10 μg/mL of transferrin, 25 μM of ethanolamine, and 25 nM of sodium selenite, where 10% (v/v) of fetal calf serum (FCS; Gibco-Invitrogen, Carlsbad, CA) and 1% (v/v) of Zap-Hybridoma solution (InVitria, Fort Collins, CO) were contained. The selected hybridomas were finally scaled up with the same medium without FCS until they reached 500 mL.
Purification of D-Glu-MAb was performed by using a Protein G FF column (0.46×11 cm; Pharmacia Biotech, Uppsala, Sweden). The cultured medium (500 mL) containing IgG was filtrated, adjusted to pH 7.0 with 1 M Tris-HCl solution (pH 9.0), and applied to the column. The column was then washed with 10 mM phosphate buffer (pH 7.0), and adsorbed IgG was eluted with 100 mM citrate buffer (pH 2.7). The eluate was neutralized with 1 M Tris-HCl (pH 9.0) immediately after elution, and the fractions, which show that an absorbance at 280 nm is over 0.8, was collected. Subsequently, they were dialyzed three times against distilled water at 4°C and lyophilized to give 28.1 mg of D-Glu-MAb.
Indirect ELISA using D-Glu-GA-HSA conjugates
The reactivity of the MAb against D-Glu-GA-HSA conjugates was determined by indirect ELISA. A 96-well immunoplate (Nunc, Maxisorb, Roskilde, Denmark) was coated with D-Glu-GA-HSA conjugates (1 μg/mL, 100 μL/well) in 50 mM carbonate buffer (pH 9.6) and incubated for 1 h. The plate was then washed three times with phosphate buffer saline (PBS) containing 0.05% (v/v) Tween-20 (PBS-T), before being treated with 300 μL of PBS containing 5% (w/v) skimmed milk (PBS-sm) for 1 h to reduce non-specific adsorption. Subsequently, various concentrations of D-Glu-MAb (100 μL/well) were incubated for 1 h. The plate was then washed three times with PBS-T and incubated with 1000-fold diluted solution of peroxidase-labeled anti-mouse IgG (100 μL/well; Organon Teknika Cappel Products) for 1 h. After washing the plate three times with PBS-T, substrate solution consisting of 0.3 mg/mL 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) in 0.1 M citrate buffer (pH 4.0) supplemented with 0.003% (v/v) H2O2 (100 μL/well) were added and incubated for 15 min. All ELISA incubation steps were carried out at 37°C. Absorbance was measured at 405 nm with a microplate reader (Maltiskan FC, Thermo Scientific). Selectivity of D-Glu-MAb against various amino acid conjugates was analyzed by indirect ELISA using D-/L-AA-GA-HSA conjugates (1 μg/mL, 100 μL/well) as coated antigen instead of D-Glu-GA-HSA conjugates.
Synthesis of D-/L-Glu-GA, D-Glu-FA, and D-Glu-VA
Synthesis of D-/L-Glu-GA, D-Glu-FA, and D-Glu-VA was performed to elucidate the epitope of D-Glu-MAb existing in the D-Glu-GA-HSA conjugates. In the case of D-/L-Glu-GA, aqueous GA (25%, 500 μL) solution was slowly added into D-/L-Glu solution (1 mg/mL, 2 mL) and stirred at room temperature for 2 h. Then NaBH4 solution (10 mg/mL, 250 μL) was added to the mixture to reduce carbon-nitrogen double bond and stirred at room temperature for 1 h. As for D-Glu-FA, aqueous FA (37%, 125 μL) was used instead of aqueous GA (25%, 500 μL) solution. With respect to D-Glu-VA, aqueous VA (135 μL) solution was gradually added into mixture of acetic acid (100 μL) and D-Glu solution containing 10% (v/v) methanol and 10% (v/v) dimethylformamide (3 mg/mL, 2 mL) and stirred at 80°C for 3 h. Subsequently the NaBH4 solution (10 mg/mL, 250 μL) was added, stirred at room temperature for 1 h, and neutralized with 10% (w/v) sodium bicarbonate (715 μL). The reaction products were serially diluted with distilled water to be used as free antigen for indirect competitive ELISA (icELISA).
Indirect competitive ELISA using D-Glu-GA-HSA conjugates
Indirect competitive ELISA (icELISA) was also carried out to identify the binding molecule of D-Glu-MAb existing in the D-Glu-GA-HSA conjugates. The same procedures that were used in the indirect ELISA were used until the blocking step. After washing the blocked-plate three times with PBS-T, various concentrations of D-Glu, D-/L-Glu-GA, D-Glu-FA, D-Glu-VA, GA, FA, VA, and glutaric acid in distilled water (50 μL/well) were incubated with D-Glu-MAb solution (42.9 ng/mL, 50 μL/well) for 1 h. The plate was then washed three times with PBS-T, and the D-Glu-MAb bound to D-Glu-GA-HSA conjugates was reacted with a 1000-fold diluted solution of peroxidase-labeled anti-mouse IgG (100 μL/well) for 1 h. After washing the plate three times with PBS-T, 100 μL of ABTS substrate solution were added to each well and incubated for 15 min. Absorbance was measured at 405 nm using a microplate reader.
Results and Discussion
Determination of hapten numbers in D-Glu-GA-BSA by MALDI-TOF-MS
The binding numbers of the D-Glu-GA molecule to the carrier BSA proteins can be evaluated by direct determination using MALDI-TOF-MS (Bruker Autoflex III). A broad peak in the MALDI-TOF-MS spectrum of D-Glu-GA-BSA appeared at around m/z 79,130 (Fig. 2). Based on the molecular weight of BSA and D-Glu-GA (66,431 and 231, respectively), at least 54 D-Glu-GA molecules were estimated to bind to the lysine residues of BSA. This result revealed that the number of D-Glu-GA was enough to elicit an immune response for the production of MAb; thus, D-Glu-GA-BSA conjugates were used as an immunogen.
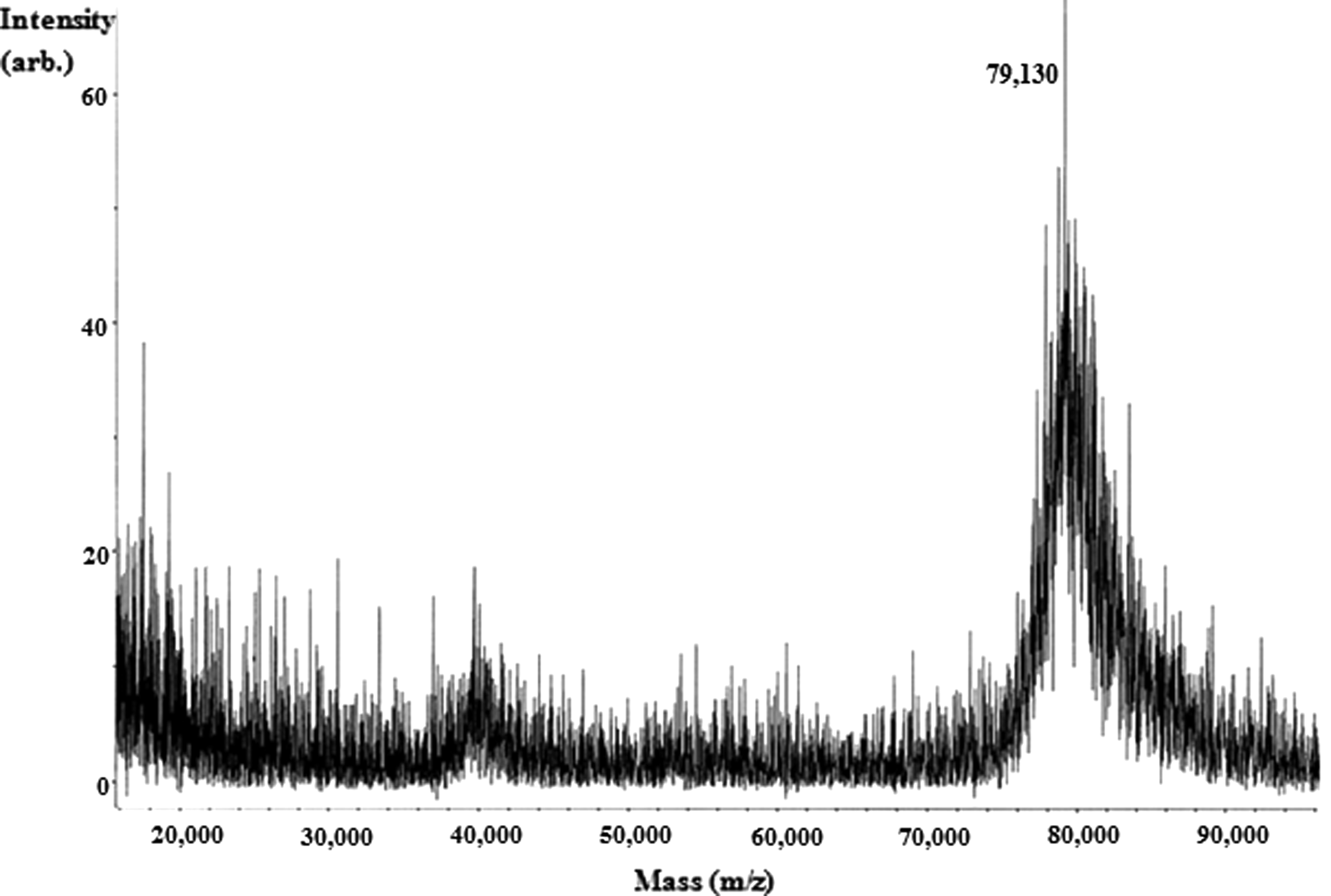
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI–TOF–MS) spectrum of D-Glu-GA-BSA conjugates.
Production and typing of MAbs against D-Glu
The D-Glu-MAb was produced by the hybridization of splenocytes obtained from D-Glu-GA-BSA immunized BALB/c mice with SP2/0 myeloma cells using the procedure developed by our laboratory.(16) One hybridoma cell line producing MAbs reactive to D-Glu-GA-HSA conjugates was screened by indirect ELISA. The typing of D-Glu-MAb using the IsoQuick Strips and Kits for Mouse Monoclonal Isotyping (Sigma-Aldrich) revealed that the MAb is classified into IgG1, which has λ light chains.
Characterization of D-Glu-MAb by indirect ELISA
The reactivity of the D-Glu-MAb to the D-Glu-GA-HSA conjugates (1 μg/mL) was primarily analyzed by indirect ELISA. The reactivity response curve was drawn by plotting absorbance against the logarithm of D-Glu-MAb concentrations in indirect ELISA. The absorbance at 405 nm was increased as MAb concentrations increased in a logarithmic manner (Fig. 3). As a result, an appropriate concentration of MAb for further indirect ELISA was found to be 21.5 ng/mL, while that for icELISA was estimated to be 42.9 ng/mL because the volume of primary antibody (50 μL) in icELISA is half of those (100 μL) in indirect ELISA as mentioned above.
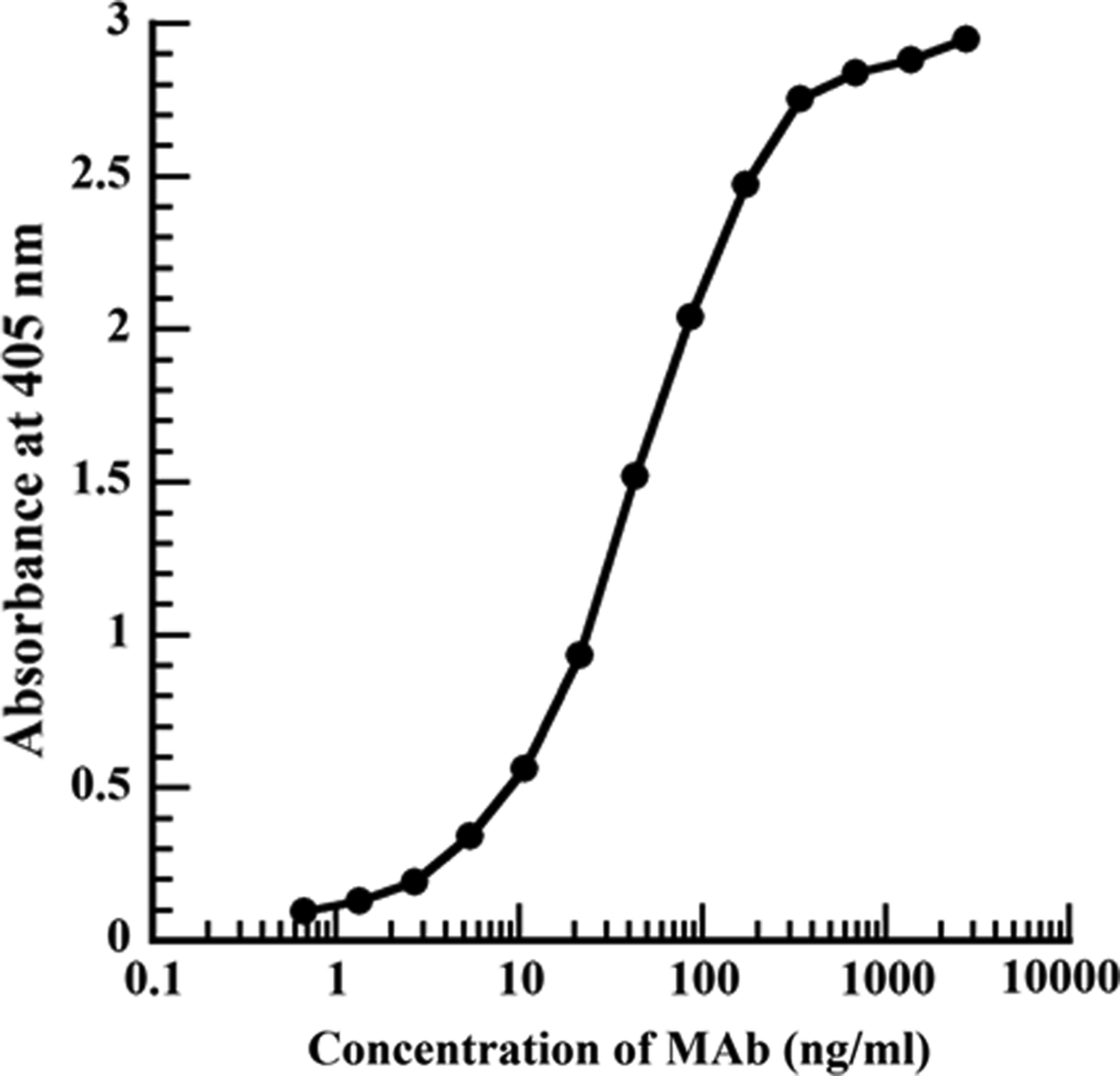
Reactivity of D-Glu-MAb to coated antigen of D-Glu-GA-HSA (1 μg/mL) conjugates by indirect ELISA. In indirect ELISA, various concentrations of D-Glu-MAb were prepared by serial double dilution with PBS-T and applied as primary antibody.
Subsequently the selectivity of D-Glu-MAb against D-/L-amino acids (AA) conjugated with HSA via GA (D-/L-AA-GA-HSA) was evaluated by indirect ELISA. In this study, 39 AA, which include each stereoisomer (D-/L-) of alanine (Ala), arginine (Arg), asparagine (Asn), aspartic acid (Asp), cysteine (Cys), glutamine (Gln), Glu, histidine (His), isoleucine (Ile), leucine (Leu), lysine (Lys), methionine (Met), phenylalanine (Phe), proline (Pro), serine (Ser), threonine (Thr), tryptophan (Trp), tyrosine (Tyr), valine (Val), and non-chiral glycine (Gly), were used as target molecule. Figure 4 shows the results of indirect ELISA, in which D-/L-AA-GA-HSA conjugates (1 μg/mL) and negative control of HSA (1 μg/mL) were used as coated antigen. Interestingly, only the wells coated with D-Glu-GA-HSA show high absorbance at 405 nm of ∼1.5, while other wells coated with D-/L-AA-GA-HSA conjugates indicate absorbance of 0.07–0.13. Considering that the absorbance of the negative control of HSA is 0.06, the D-Glu-MAb possesses high selectivity to D-Glu-GA-HSA conjugates rather than others. These results suggest that the epitope of D-Glu-MAb is estimated to be D-Glu and/or D-Glu-GA.
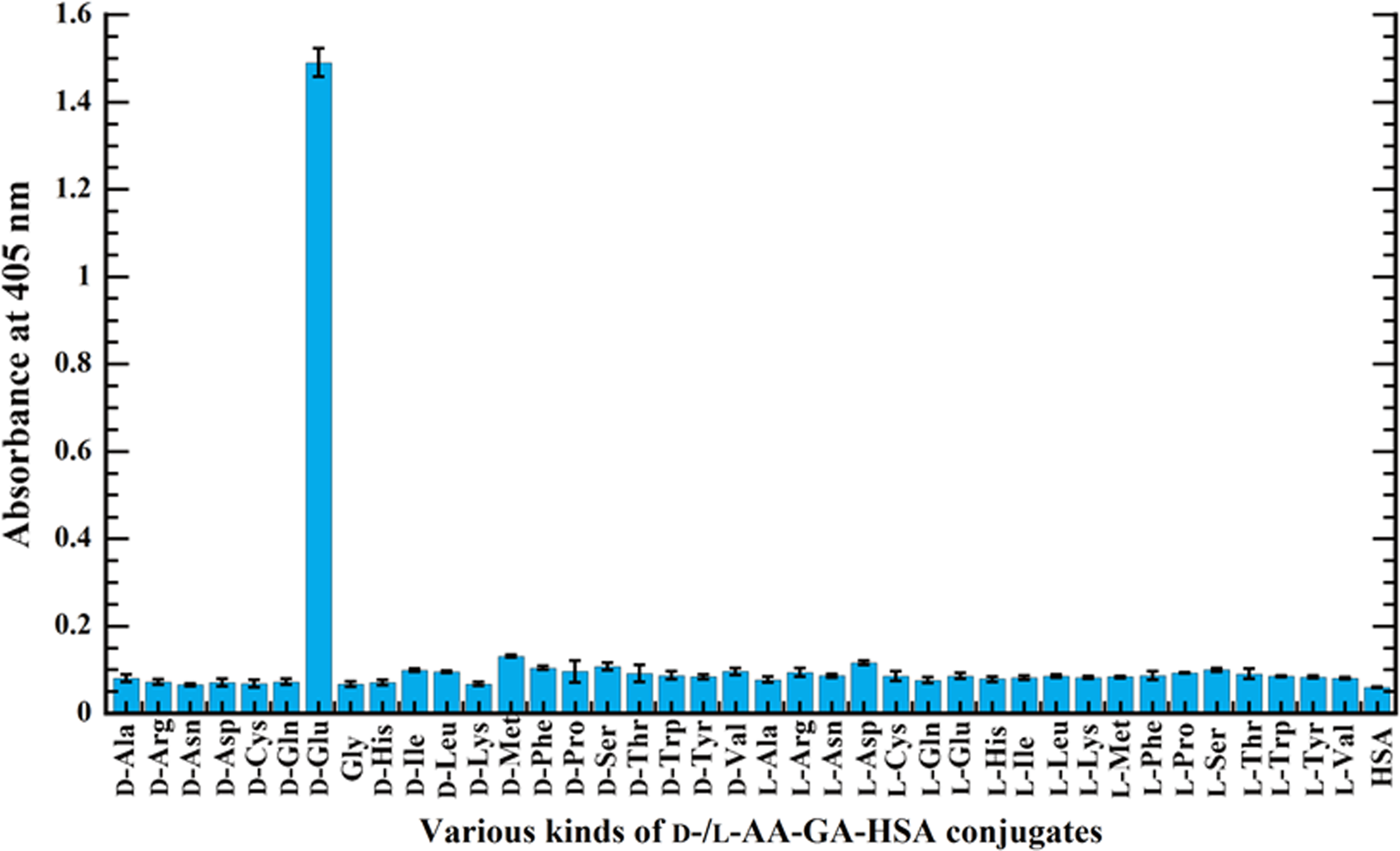
Reactivity of D-Glu-MAb to various D-/L-AA-GA-HSA (1 μg/mL) conjugates by indirect ELISA. HSA (1 μg/mL) was used as negative control.
Epitope identification of D-Glu-MAb by icELISA
To identify the epitope of D-Glu-MAb, further analysis was carried out by an icELISA using D-Glu, D-/L-Glu-GA, D-Glu-FA, D-Glu-VA, GA, FA, VA, and glutaric acid as free antigen. A D-Glu-MAb (42.9 ng/mL) was incubated with serially diluted free antigens on an immunoplate. Any D-Glu-MAb binding to free antigens was washed out, while only the D-Glu-MAb bound to the immobilized D-Glu-GA-HSA conjugates were incubated with peroxidase-labeled anti-mouse IgG and then treated with ABTS solution to develop color. As a result, any inhibitory activity was observed when D-Glu, L-Glu-GA, and glutaric acid were used as free antigen (data not shown), whereas inhibitory activity was observed in the case of D-Glu-GA, D-Glu-FA, and D-Glu-VA (Fig. 5), and GA, FA, and VA. When the 50% inhibitory concentration values (IC50) were compared among three compounds (D-Glu-GA, D-Glu-FA, and D-Glu-VA), IC50 of D-Glu-GA (0.54 μg/mL) exhibited ∼300 times lower than those of D-Glu-VA (149 μg/mL) and ∼600 times lower than those of D-Glu-FA (327 μg/mL), suggesting that inhibitory activity of D-Glu-MAb becomes stronger as the length of the side chain structurally similar to D-Glu-GA. Although inhibitory activity was also confirmed in the high concentration of GA, FA, and VA with the IC50 of 23.6, 21.1, and 37.7 mg/mL, respectively (data not shown), it is concluded that this inhibitory activity is attributed to non-specific binding of aldehyde to the amino group of D-Glu-MAb because there are many aldehyde-reactive sites in IgG molecules; this phenomenon was also observed when other MAbs produced by a completely different immunogen such as daidzin, glycyrrhizin, and plumbagin were used instead.(17,18) Collectively, these results suggest that the epitope of D-Glu-MAb is D-Glu-GA. GA and FA are frequently used as the reagent for fixation in immunostaining; thus, D-Glu-MAb can be a helpful tool to investigate the functional analysis of D-Glu in vivo and in vitro.

Calibration curves of (
Conclusion
In this study, we successfully produced the D-Glu-MAb, which is highly specific to D-Glu. Considering the epitope of D-Glu-MAb is D-Glu-GA, it can be applied to not only histopathological immunostaining but also a probe for determination of D-Glu by preparing the D-Glu-GA conjugates in an immunosorbent assay. In addition, various antibodies such as single-chain variable fragment antibody (scFv), fragment antigen binding (Fab), bis-scFv, and minibody can be genetically engineered by using a hybridoma producing D-Glu-MAb.(19) This makes it possible to transform and express the antibodies against D-Glu in various hosts (bacteria, plant, and mammal) to investigate the dynamic behavior of D-Glu in situ. Since very little research has been conducted on D-Glu in the body, the D-Glu-MAb may help to reveal functional aspects of D-Glu that remain unclear.
Footnotes
Acknowledgments
This study was funded by the Sasagawa Scientific Research Grant from the Japan Science Society.
Author Disclosure Statement
The authors have no financial interests to disclose.