
Abstract
Podoplanin (PDPN) is a platelet aggregation-inducing factor, which is known as an endogenous ligand of C-type lectin-like receptor-2 (CLEC-2). PDPN is also expressed in several normal tissues, such as lung type I alveolar cells and kidney podocytes. Although many monoclonal antibodies (MAbs) against human PDPN (hPDPN) or mouse PDPN (mPDPN) have been established, anti-rat PDPN (rPDPN) MAbs, especially against platelet aggregation-stimulating (PLAG) domain (29-54 amino acids) of rPDPN, have not been developed. Therefore, functional analysis of rPDPN in normal tissues has been limited. Here, we immunized mice with rPDPN peptides (38-51 amino acids) and developed a novel mouse anti-rPDPN MAb, PMab-2 (IgG1, kappa), which possesses high affinity compared with anti-hPDPN or mPDPN MAbs. The KD of PMab-2 was determined to be 5.9 × 10−10 M. PMab-2 is useful, not only in flow cytometry and Western blot analysis against endogenous rPDPN, which is expressed in rat dermal fibroblast, but also in immunohistochemistry against normal tissues. PMab-2 showed extraordinarily high sensitivity in immunohistochemistry, indicating that PMab-2 is very advantageous for functional analysis of rPDPN.
Introduction
P
Materials and Methods
Cell lines
Chinese hamster ovary (CHO)-K1 and P3U1 were purchased from the American Type Culture Collection (ATCC, Manassas, VA). Rat dermal fibroblast was obtained from Cell Applications (San Diego, CA). CHO-K1 was transfected with rPDPN, hPDPN, or mPDPN plasmids (CHO/rPDPN, CHO/hPDPN, CHO/mPDPN) using Lipofectamine 2000 (Thermo Fisher Scientific, Waltham, MA).(3) CHO-K1, CHO/rPDPN, CHO/hPDPN, CHO/mPDPN, and P3U1 were cultured in RPMI 1640 medium including RPMI 2 mM L-glutamine (Nacalai Tesque, Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (FBS, Thermo Fisher Scientific), 100 U/mL penicillin, 100 μg/mL streptomycin, and 25 μg/mL amphotericin B (Nacalai Tesque), and rat dermal fibroblast was cultured in Fibroblast Growth Medium 2, including FBS, basic fibroblast growth factor (1 ng/mL), and insulin (5 μg/mL; PromoCell, Heidelberg, Germany) at 37°C in a humidified atmosphere of 5% CO2 and 95% air.
Animals
Female BALB/c mice (four-weeks-old) were purchased from CLEA Japan (Tokyo, Japan). Animals were housed under pathogen-free conditions. The Animal Care and Use Committee of Tohoku University approved the animal experiments described herein.
Hybridoma production
BALB/c mice were immunized by intraperitoneal (i.p.) injection of 100 μg of synthetic peptides (rpp3851: GDDMV NPGLEDRIE
ELISA
Synthetic peptides were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific) at 1 μg/mL for 30 min. After blocking with SuperBlock T20 (PBS) Blocking Buffer, the plates were incubated with culture supernatant followed by 1:1000 diluted peroxidase-conjugated anti-mouse IgG (Dako, Glostrup, Denmark). The enzymatic reaction was conducted with a 1-Step Ultra TMB-ELISA (Thermo Fisher Scientific). The optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Philadelphia, PA). These reactions were performed with a volume of 50 μL at 37°C.
Western blot analyses
Cell lysates (10 μg) were boiled in SDS sample buffer (Nacalai Tesque). The proteins were electrophoresed on 5–20% polyacrylamide gels (Wako Pure Chemical Industries, Osaka, Japan) and were transferred onto a PVDF membrane (EMD Millipore, Billerica, MA). After blocking with 4% skim milk (Nacalai Tesque), the membrane was incubated with primary antibodies, 43 and then with peroxidase-conjugated secondary antibodies (Dako; 1:1000 diluted), and developed with Pierce Western Blotting Substrate Plus (Thermo Fisher Scientific) or ImmunoStar LD Chemiluminescence Reagent (Wako Pure Chemical Industries) using a Sayaca-Imager (DRC Co., Tokyo, Japan).
Flow cytometry
Cells were harvested by brief exposure to 0.25% Trypsin/1 mM EDTA (Nacalai Tesque). After washing with PBS, the cells were treated with primary MAbs (1 μg/mL) for 30 min at 4°C, followed by treatment with Oregon green-conjugated anti-mouse IgG or anti-rat IgG (Thermo Fisher Scientific). Fluorescence data were collected using a Cell Analyzer EC800 (Sony Corp., Tokyo, Japan).
Determination of binding affinity by flow cytometry
Binding affinity was determined as described previously.(44) Briefly, cells (2 × 105 cells) were resuspended at 100 μL of serially diluted antibodies (0.02–100 μg/mL), followed by secondary antibodies (Thermo Fisher Scientific). Fluorescence data were collected using a Cell Analyzer EC800. The dissociation constants (KD) were obtained by fitting the binding isotherms using the built-in one-site binding models in GraphPad PRISM 6 (GraphPad Software, La Jolla, CA).
Immunohistochemical analyses
Four-μm-thick histologic sections were deparaffinized in xylene and rehydrated. Sections were incubated with PMab-2 for 1 h at room temperature followed by treatment with Envision+ kit (Dako). Color was developed using 3,3-diaminobenzidine tetrahydrochloride (DAB, Dako) for 5 min, and then the sections were counterstained with hematoxylin (Wako Pure Chemical Industries).
Results
Production of monoclonal antibody against rPDPN
We first immunized mice with a synthetic peptide of rat PDPN; then ELISA screening was performed. Among 23 ELISA-positive wells, 11 wells reacted with CHO/rPDPN in flow cytometry. Furthermore, three wells recognized a specific band in Western blot analysis. After limiting dilution, clone PMab-2 (mouse IgG1, kappa) was established. PMab-2 detected about 40 kDa band of rPDPN-transfected CHO cells (CHO/rPDPN), not with hPDPN-transfected CHO cells (CHO/hPDPN), mPDPN-transfected CHO cells (CHO/mPDPN), and CHO cells in Western blot analysis (Fig. 1A). In contrast, NZ-1 and PMab-1 detected about 40 kDa bands of CHO/hPDPN and CHO/mPDPN, respectively, indicating that three MAbs are species-specific. PMab-2 reacted with CHO/rPDPN, not with CHO/hPDPN, CHO/mPDPN, and CHO cells in flow cytometry (Fig. 1B). NZ-1 and PMab-1 specifically reacted with CHO/hPDPN and CHO/mPDPN, respectively. Taken together, PMab-2 is a specific MAb against rPDPN, and is very useful in Western blot and flow cytometric analyses. We next investigated whether PMab-2 can detect endogenous rPDPN. Because PDPN has been reported to be expressed in mouse or human fibroblast cells, we checked the PMab-2 reactivity against rat dermal fibroblast. As shown in Figure 2A, PMab-2 detected not only exogenous rPDPN of CHO/rPDPN but also an endogenous one, which is expressed in rat dermal fibroblast. PMab-2 also reacted with rat fibroblast in flow cytometry (Fig. 2B). These results indicate that PMab-2 is useful for detecting endogenous rPDPN. We further performed a kinetic analysis of the interaction of PMab-2 with CHO/rPDPN using flow cytometry. As shown in Figure 3, KD of PMab-2 was determined to be 5.9 × 10−10 M using CHO/rPDPN. In contrast, KD of NZ-1 was determined to be 2.9 × 10−9 M using CHO/hPDPN and KD of PMab-1 was determined to be 3.2 × 10−9 M using CHO/mPDPN, indicating that the binding affinity of PMab-2 is much higher than that of NZ-1 and PMab-1.

Specific binding of anti-PDPN MAbs. (
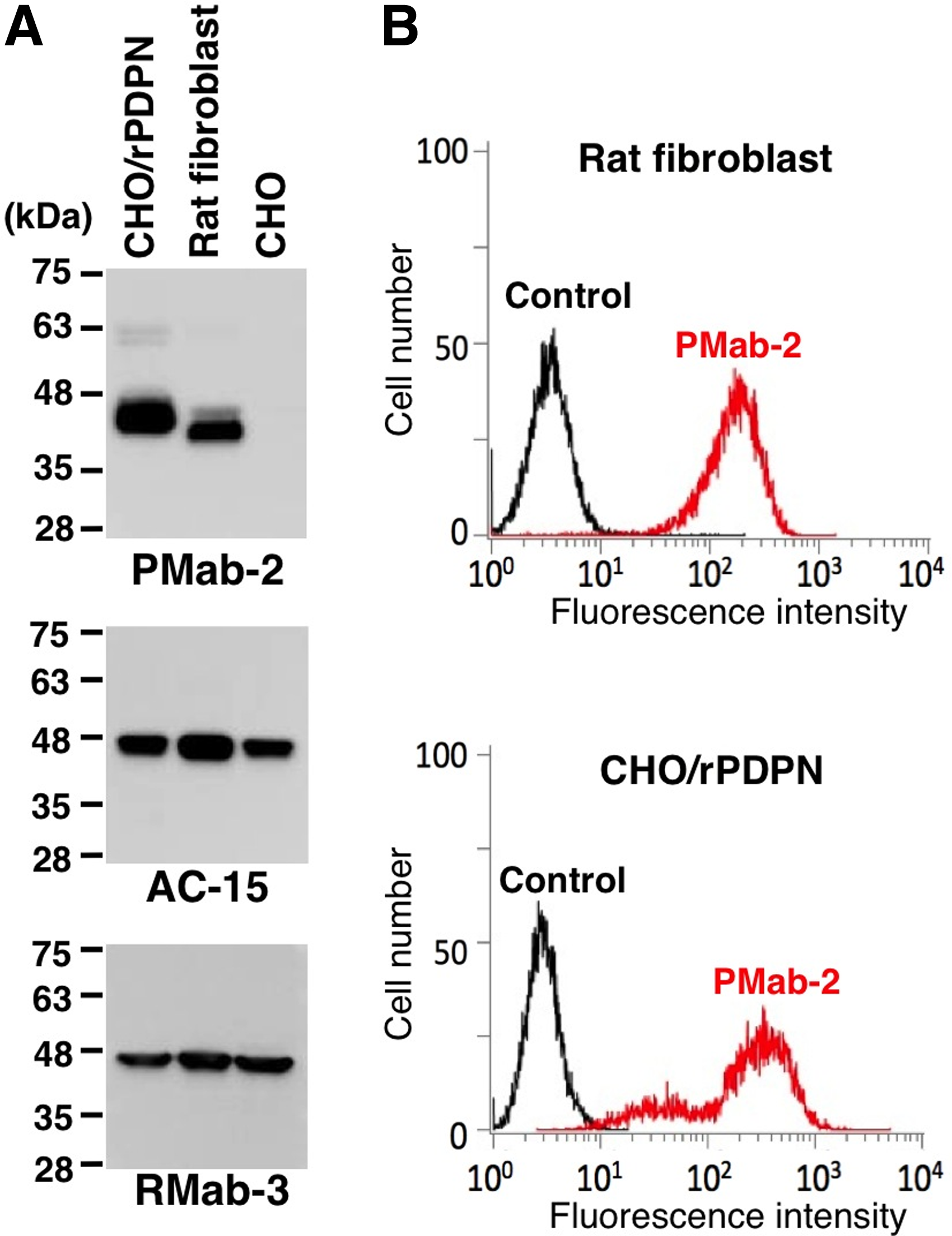
Endogenous rPDPN is detected by PMab-2. (
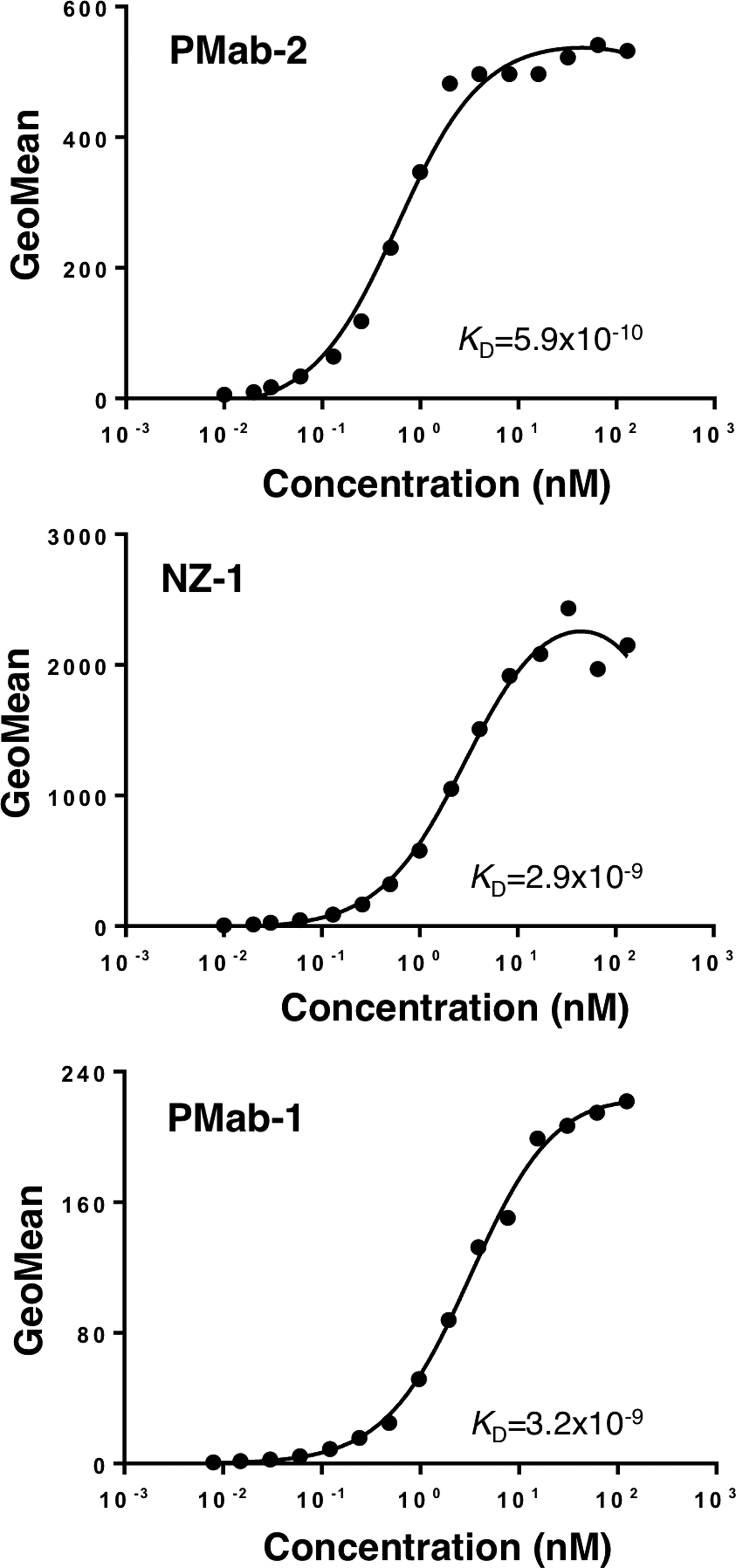
Determination of binding affinity using flow cytometry. CHO = rPDPN, CHO = hPDPN, and CHO = mPDPN (2 × 10s cells) were re-suspended at 100 μL of serially diluted PMab-2, NZ-1, and PMab-1 (0.02–100 μg/mL, respectively. Fluorescence data were collected using a Cell Analyzer EC800. KD was obtained by fitting binding isotherms using built-in one-site binding models in Prism software.
Immunohistochemical analysis using PMab-2
We next investigated whether PMab-2 can detect rPDPN of normal tissues in immunohistochemistry. PMab-2 reacted with type I alveolar cells of lung, lymphatic endothelial cells of colon, podocytes, and Bowman's capsule epithelial cells of kidney (Fig. 4). Because PMab-2 showed high affinity against rPDPN, we next checked the dose dependency of PMab-2 in immunohistochemistry using three different tissues. As shown in Figure 5, PMab-2 detected rPDPN of lung type I alveolar cells at 0.001 μg/mL, rPDPN of colon lymphatic endothelial cells at 0.005 μg/mL, and rPDPN of kidney podocytes at 0.01 μg/mL, indicating that PMab-2 is very sensitive against rPDPN.

Immunohistochemical analysis by PMab-2. Sections of rat lung, colon, and kidney were incubated with 0.5 μg/mL of PMab-2 or control PBS, followed by Envision+ kit, and color was developed using DAB and counterstained with hematoxylin. Arrows, lymphatic vessels; arrowheads, glomerulus. Scale bar, 100 μm.
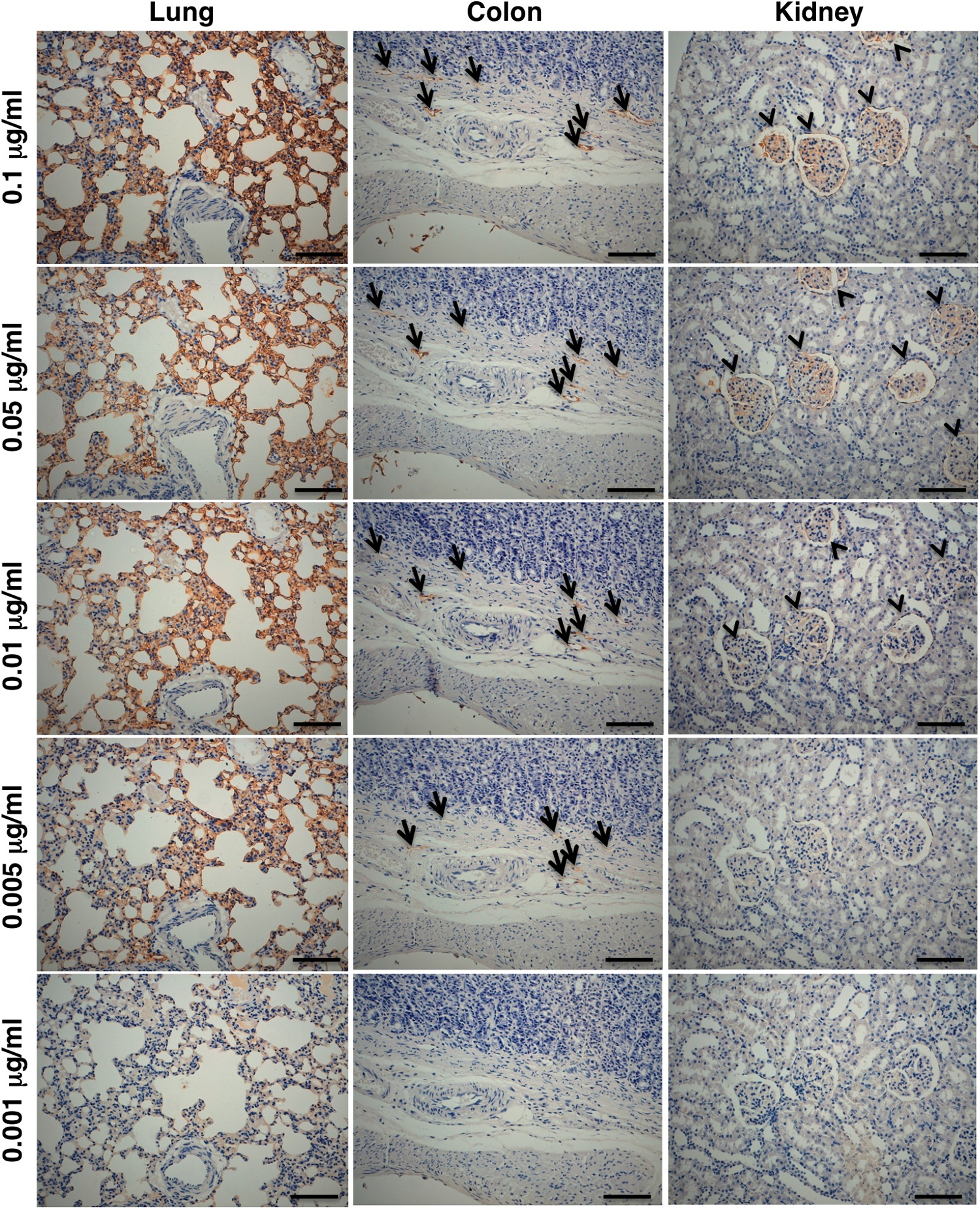
Immunohistochemical analysis by PMab-2 in a dose-dependent manner. Sections of rat lung, colon, and kidney were incubated with 0.001-0.1 μg/mL of PMab-2, followed by Envision+ kit, and color was developed using DAB and counterstained with hematoxylin. Arrows, lymphatic vessels; arrowheads, glomerulus. Scale bar, 100 μm.
Discussion
In this study, we immunized mice with a synthetic peptide (rpp3851), which is included in platelet aggregation-stimulating (PLAG) domain of rPDPN. We previously produced an anti-hPDPN MAb (NZ-1)(17) and an anti-mPDPN MAb (PMab-1)(39) by immunizing rats with PLAG domain of hPDPN and mPDPN, respectively. Because these MAbs are not only very sensitive and specific but also very useful for almost all experiments including flow cytometry, Western blot, immunoprecipitation, immunocytochemistry, and immunohistochemistry, MAbs against PLAG domain of rPDPN was thought to be advantageous. As expected, established PMab-2 is a specific MAb against rPDPN, and is very useful in Western blot and flow cytometric analyses. Furthermore, PMab-2 detected not only exogenous rPDPN but also an endogenous one of dermal fibroblast in both Western blot and flow cytometry, which leads to the advantage in immunohistochemistry. By kinetic analysis of the interaction of PMab-2 using flow cytometry, KD of PMab-2 was determined to be 5.9 × 10−10 M, indicating that the binding affinity of PMab-2 is extraordinarily high. In rat normal tissues, rPDPN was reported to be highly expressed in lymphatic endothelial cells, lung type I alveolar cells, or kidney podocytes.(1) Expectedly, PMab-2 reacted with type I alveolar cells of lung, lymphatic endothelial cells of colon, and podocytes of kidney in immunohistochemistry. A previous study showed that PMab-1 detects mPDPN in immunohistochemistry; however, antigen retrieval procedure was necessary.(39) Furthermore, NZ-1 detects hPDPN in immunohistochemistry without antigen retrieval; however, it does not react with type I alveolar cells of lung and podocytes of kidney.(20) In contrast, PMab-2 does not need antigen retrieval and detects normal rPDPN, which is expressed in various tissues. Surprisingly, PMab-2 detected rPDPN of lung type I alveolar cells at a low concentration of 1 ng/mL, indicating that PMab-2 is extraordinarily sensitive against rPDPN.
Taken together, PMab-2 is very useful for all experiments such as flow cytometry, Western blot, and immunohistochemistry. Furthermore, the epitope of PMab-2 is PLAG domain of rPDPN, indicating that PMab-2 is advantageous for investigating the platelet aggregation activity of rPDPN or rPDPN-CLEC-2 interaction.
Footnotes
Acknowledgments
We thank Takuro Nakamura, Noriko Saidoh, Hazuki Kanno, and Kanae Yoshida for their excellent technical assistance. This work was supported in part by the Platform for Drug Discovery, Informatics, and Structural Life Science (PDIS) from Japan Agency for Medical Research and Development, AMED (Y.K.); by the Basic Science and Platform Technology Program for Innovative Biological Medicine from AMED (Y.K.); by the Regional Innovation Strategy Support Program from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) of Japan (Y.K.); and by JSPS KAKENHI Grant Number 26440019 (M.K.K.) and Grant Number 25462242 (Y.K.).
Author Disclosure Statement
The authors have no financial interests to disclose.