
Abstract
Integrins are a major family of adhesion molecules, consisting of heterodimers (α and β subunits). Several reports have suggested the presence of splice variants in the cytoplasmic domain of certain integrin subunits. In the present study, we detected mRNA of integrin α3 splice variants (α3A and α3B) by RT-PCR using total RNA from the human brain as a template. The α3B variant lacks the sequence coded by exon 25 and appears to be generated by alternative splicing. We established mouse hybridomas producing monoclonal antibodies (both of which are of IgG1 class) specific for each variant. Each antibody exhibited specific reactivity towards the corresponding integrin α3 variant in Western blotting and immunoprecipitation experiments, suggesting it to be a useful tool for detection of the respective integrin variant.
Introduction
T
The presence of splice variants of integrin α subunits (e.g., α3, α4, α5, α6, α7, and α9) has been reported,(8–13) and some of these exhibit differences in amino acid sequences of the cytoplasmic domains (e.g., α3A/3B, α4/4B, α6A/6B, and α7A/7B/7C).(8,9,11,12) Because they possess identical extracellular domains but distinct cytoplasmic domains, each splice variant might be involved in a specific signal transduction pathway. The outside-in signals mediated by the interaction between α3β1 integrin and laminin-511/521 were reported to promote cell migration and suppress apoptosis induced by serum starvation;(14,15) these processes are believed to be mediated by the α3Aβ1 heterodimer because of the relative abundance of the α3A subunit in most cells. It would therefore be quite intriguing to characterize the functional difference between these splice variants. The splice variant α3B subunit has been identified only in mouse cells and has not been found in human cells. In this study, therefore, we attempted to identify mRNA of the human α3B subunit and to generate monoclonal antibodies that can distinguish the α3A and α3B subunits.
Materials and Methods
Reagents
Restriction endonucleases were purchased from Thermo Scientific (Rockford, IL). TRIzol reagent, SuperScript III reverse transcriptase, and Dynabeads protein G were purchased from Life Technologies (Grand Island, NY). The PrimeSTAR MAX DNA polymerase was a product of Takara Bio (Shiga, Japan). Nonidet P-40 was purchased from Nacalai Tesque (Kyoto, Japan). The two-dimensional silver stain II kit was from Daiichi Pure Chemical (Tokyo, Japan). Polyethylenimine was purchased from Polysciences (Warrington, PA). Oligonucleotides were supplied by Fasmac Co. (Kanagawa, Japan). Peptides and keyhole limpet hemocyanin (KLH)-conjugated peptides were synthesized by Scrum Co. (Tokyo, Japan) and Sigma (St. Louis, MO). Horseradish peroxidase-conjugated goat antibodies to mouse IgG and rabbit IgG were purchased from Kirkegaard & Perry Laboratories (Guildford, United Kingdom) and Caltag Laboratories (Burlingame, CA), respectively. Anti-actin antibody (rabbit polyclonal antibody) was purchased from Sigma.
Mice
BALB/c mice at 6 weeks of age were purchased from Japan SLC (Shizuoka, Japan). All procedures for experiments using mice were conducted in accordance with the Guide for the Care and Use of Laboratory Animals of the Hoshi University School of Pharmacy and Pharmaceutical Sciences (Tokyo, Japan).
Reverse transcription-polymerase chain reaction
Total RNA was isolated from several mouse tissues with TRIzol reagent, and cDNA was synthesized using a Transcriptor First Strand cDNA Synthesis kit (Roche Diagnostics, Indianapolis, IN). Polymerase chain reaction (PCR) was performed with a KAPATaq EXtra PCR kit (Nippon Genetics Co., Tokyo, Japan) and the following primers: itga3, 5′-AAGCCAAGTCTGAGACTGTG-3′ (ITGA3-Ex21_F) and 5′-CTTTGGACTGTCGGAGTCATAA-3′ (mITGA3-Ex26_R); and gapdh, 5′-TGAAGCAGGCATCTGAGGG-3′ and 5′-CATGTAGGCCATGAGGTCCAC-3′.
Human brain total RNA was supplied by Clontech Laboratories (Mountain View, CA) and reverse-transcribed with SuperScript III reverse transcriptase using an oligo-dT primer. In order to detect α3A and α3B integrin mRNA, PCR was conducted with the following set of primers: ITGA3-Ex21_F (described above) and 5′-CTGTGGACTGTCAGAGGCATAA-3′ (hITGA3-Ex26_R). The PCR products were separated in agarose gel (2%) electrophoresis and stained with ethidium bromide. The DNA sequence was determined by the dideoxy method (Fasmac DNA Sequence Service, Tokyo, Japan).
Establishment of hybridomas
Monoclonal antibodies specific for human α3A or α3B integrin were generated by fusing PAI myeloma cells with splenocytes of BALB/c mice immunized with a peptide Cys-YEAKRQKAEMKSQ or Cys-QIMPKYHAVRIREEERY conjugated with KLH. The conjugation of the peptides with KLH and immunization of mice were performed as described previously.(16) Hybridomas were selected in hypoxanthine/aminopterin/thymidine (HAT) medium (Sigma), and the culture supernatants were screened by ELISA using the same peptides as used for immunogens. Monoclonal antibodies from clone 2F9 (IgG1/κ) against the α3A integrin subunit and from clone 4B4-5 (IgG1/κ) against the α3B integrin subunit were selected in this study. These hybridomas were cultured in a serum-free medium (ASF medium 104, Ajinomoto Co., Tokyo, Japan), and antibodies were purified with a HiTrap protein G HP column (GE Healthcare, Cleveland, OH). The isotypes of monoclonal antibodies were determined by using an IsoStrip mouse monoclonal antibody isotyping kit (Roche Diagnostics).
Cell culture and transfection
HEK293FT (Invitrogen, Carlsbad, CA) and PAI myeloma cells were grown in RPMI1640 medium (Sigma) supplemented with 10% (v/v) heat-inactivated fetal calf serum (Biosera, Kansas City, MO) at 37°C under humidified 5% CO2. The plasmid encoding α3A or α3B integrin was introduced into HEK293FT cells with polyethylenimine MAX (PEI, Mw 40,000).(17) Briefly, 30 μg of plasmid DNA and 150 μL of PEI (0.323 g/L) were mixed in 500 μL of RPMI1640 medium (serum-free). The DNA-PEI complex was added to a culture of HEK293FT cells (2 × 107 cells/15 cm plate), and the cells were cultured at 37°C for 48 h.
Electrophoresis and immunoblotting
HEK293FT transfectant cells were incubated in the lysis buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM EDTA, 1% Nonidet P-40) at 4°C for 30 min, and the cell lysate was centrifuged at 20,000 g for 30 min. The resultant supernatant was mixed with an equal volume of 2x SDS sample buffer (100 mM Tris-HCl [pH 6.8], 2% SDS, 10% glycerol, 0.02% bromophenol blue) for SDS-polyacrylamide gel electrophoresis (PAGE). SDS-PAGE and immunoblotting were conducted as described previously.(18)
Immunoprecipitation
Anti-α3A or anti-α3B antibody (3 μg) was incubated with Dynabeads protein G (15 μL) with gentle agitation in a rotator for 30 min at room temperature. The conjugate of Dynabeads protein G and antibody thus obtained was mixed with the lysate of HEK293FT transfectant cells, and the mixture was incubated at 4°C for 16 h. The beads were washed five times with the lysis buffer to remove unbound proteins. Proteins bound to the beads were recovered by the treatment with an SDS-sample buffer at 95°C for 3 min and subjected to SDS-PAGE.
Construction of plasmids
The cDNA for a full-length human integrin α3 subunit gene (ITGA3) was amplified by PCR using cDNA derived from the human brain as a template. PCR was conducted with the following set of primers: 5′-tggcggccgctcgagATGGGCCCCGGCCCCAGCCG-3′ (inf_hITGA3-Ex1_F) and 5′-ccctctagactcgagCTGTGGACTGTCAGAGGCATAA-3′ (inf_hITGA3-Ex26_R). The product was cloned into the Xho I restriction site of pcDNA6-V5HisA (Invitrogen) by using an in-fusion HD cloning kit (Takara Bio). To identify α3A and α3B variant cDNA, PCR was conducted using primers with sequences corresponding to exon 21 (ITGA3-Ex21_F) and exon 26 (hITGA3-Ex26_R). The full-length cDNA sequences of the α3A and α3B subunits were then determined by the dideoxy method.
Results
Expression of α3A and α3B integrin mRNA in the human brain
Our previous study indicated that the mouse integrin α3 subunit gene (Itga3) is encoded by 26 exons, and that the cytoplasmic domain splice variants of the α3 subunits (α3A and α3B) are generated by alternative exon usage.(19) The mouse α3B subunit lacks the sequence coded by exon 25 but includes an alternative cytoplasmic sequence coded by exon 26 (Fig. 1A). Since the mRNA for the α3B subunit lacks the sequence corresponding to exon 25, it can be distinguished from that for the α3A subunit by RT-PCR using a set of primers with sequences corresponding to exons 21 and 26 (Fig. 1A). It was also reported that the α3A subunit was widely expressed in almost all tissues, whereas expression of the α3B subunit was limited to the brain and heart in mice.(8) Therefore, we first examined the expression profiles of α3A and α3B subunits in several mouse tissues. The results demonstrated that the mRNAs for both the α3A and α3B subunits were detected in the brain but that other organs expressed α3A mRNA exclusively (Fig. 1B). These results are consistent with those from the previous study by Tamura and colleagues.(20) We next examined whether human organs also expressed a splice variant similar to the mouse integrin α3B subunit. Because the genomic structures of human and mouse integrin α3 subunit are quite similar,(19,21) we conducted RT-PCR using primers with sequences corresponding to exons 21 and 26 and cDNA derived from human brain total RNA. As shown in Figure 2, we obtained doublet bands of PCR products in electrophoresis when brain-derived mRNA was used as a template, whereas we detected only a single PCR product corresponding to α3A from human umbilical vein endothelial cell (HUVEC) or MKN1 (human adenosquamous carcinoma) cell-derived mRNAs (Fig. 2). The nucleotide sequence analysis of these PCR products derived from the human brain revealed that the upper band (743 bp) was composed of the authentic α3A integrin and that the sequence of the lower band (601 bp) was consistent with an α3B integrin variant generated by skipping exon 25 (Fig. 3). These results indicate that the α3B integrin splice variant with a distinct cytoplasmic sequence coded by exon 26 was also expressed in the human brain as well as in the mouse. To our knowledge, this is the first time the presence of an α3B integrin variant has been demonstrated in humans. We then screened several human malignant tumor cell lines for expression of the α3B variant, but none of the cell lines tested, including A172 glioblastoma, A375 malignant melanoma, EJ-1 bladder carcinoma, HepG2 hepatocellular carcinoma, HT1080 fibrosarcoma, SN12C renal carcinoma, and U-251MG glioblastoma, gave the PCR product corresponding to the α3B integrin variant (data not shown).

Expression of α3A and α3B integrin variants in various murine organs. (
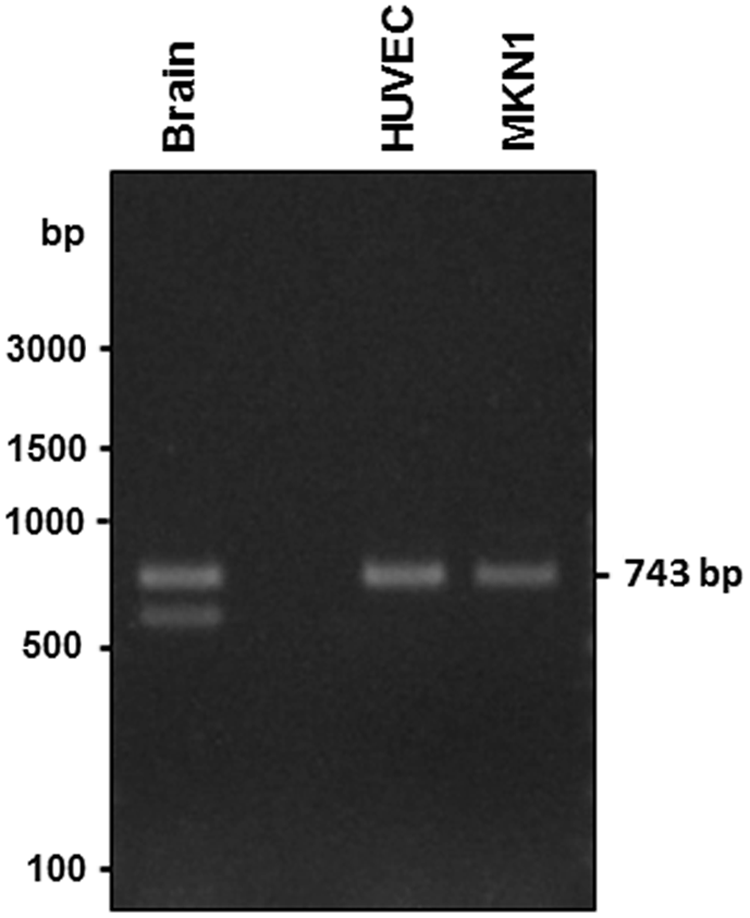
Expression of α3A and α3B integrin variants in the human brain. mRNAs expressed in the human brain and HUVEC and MKN1 cells were analyzed by RT-PCR. PCR products (743 and 601 bp) from mRNA for splice variants of ITGA3 (α3A and α3B integrins, respectively) were detected when mRNA from the human brain was used as a template.

Nucleotide and deduced amino acid sequences of PCR products derived from α3 integrin variants. Nucleotide sequences of PCR products shown in Figure 2 are described. Upper and lower PCR product bands were deduced to be α3A (left) and α3B (right) variants, respectively, because the lower band was deficient in the sequence corresponding to exon 25. The upper-case letters and asterisks indicate primer sequences and a stop codon, respectively.
We next cloned the full-length cDNA to α3A and α3B subunits from a human brain cDNA library. The RT-PCR was conducted using primers with sequences corresponding to exons 1 and 26, and the products were subcloned into pcDNA6-V5HisA. The nucleotide sequence analysis of the clones indicated that the clones of the α3A and α3B subunits had identical sequences in the extracellular and transmembrane domains, and the difference in nucleotide sequence between these variants existed only in the cytoplasmic domain (Fig. 3).
Establishment of hybridomas producing anti-α3A and α3B integrin antibodies
We next attempted to establish hybridomas producing monoclonal antibodies specific for the α3A and α3B integrin subunits. The difference in amino acid sequence between the α3A and α3B variants was found to reside in the cytoplasmic domain (Fig. 4). The two peptides with underlined sequences in Figure 4 were used in the immunization for splice variant-specific antibodies to α3 integrin. Then, hybridomas were screened for α3A and α3B integrins by enzyme-linked immunosorbent assay (ELISA) using the same peptides as used for immunogens. After repeated limiting dilution, we obtained hybridomas producing variant-specific anti-α3 integrin monoclonal antibodies, clone 2F9 (IgG1/κ) against α3A integrin and clone 4B4-5 against α3B integrin (IgG1/κ).

Amino acid sequences of cytoplasmic domain of α3 integrin variants. The α3A and α3B integrin variants differed in amino acid sequence of the cytoplasmic domain coded by exon 25 or 26. Underlined amino acid sequences indicate peptides used for immunization of mice.
Application of monoclonal antibodies against α3A and α3B integrin variants
To analyze the specificity of the anti-α3A integrin (2F9) and anti-α3B integrin (4B4-5) antibodies, we performed immunoblotting using HEK293FT cells that had been transfected with cDNA for the α3A or α3B integrin subunit. As shown in Figure 5, the anti-α3A antibody (2F9) reacted with α3 integrin in the lysate of α3A-transfected HEK293FT cells but not with α3 integrin in the lysate of α3B-transfected cells; the reverse was true for anti-α3B integrin antibody (4B4-5), which reacted with α3 integrin in the lysate of α3B-transfected cells but not with α3 integrin in the lysate of α3A-transfected cells. The molecular size of each band showed no significant difference and was approximately 130 kDa. These results clearly indicate that the antibodies established in this study possess specific reactivity to each of the two splicing variants, α3A and α3B integrins.

Immunoblotting analysis with splice variant-specific antibodies against α3 integrin. Cell lysates of the HEK293FT transfectants (mock, α3A or α3B) were separated by SDS-PAGE (7.5% polyacrylamide gel) and analyzed by immunoblotting with α3A and α3B integrin-specific antibodies. Anti-actin antibody was used as a control.
We next examined whether these antibodies could be applied to an immunoprecipitation assay. Dynabeads protein G conjugated with anti-α3A antibody precipitated an intensely stained and rather broad band of approximately 130 kDa from the lysate of α3A-transfected cells in SDS-PAGE; no such intense band was obtained from mock or α3B-transfected cells (Fig. 6A). By contrast, the protein G/anti-α3B antibody conjugate immunoprecipitated an intense band of approximately 130 kDa from α3B-transfected cells, but no specific band around 130 kDa was detected from the lysates of mock cells or α3A-transfectants (Fig. 6B). Taken together, these results show that the monoclonal antibodies against α3 integrin newly developed in this study successfully distinguished α3 integrin splice variants, and would be effective tools for use in ELISA, immunoblotting, or immunoprecipitation assays.
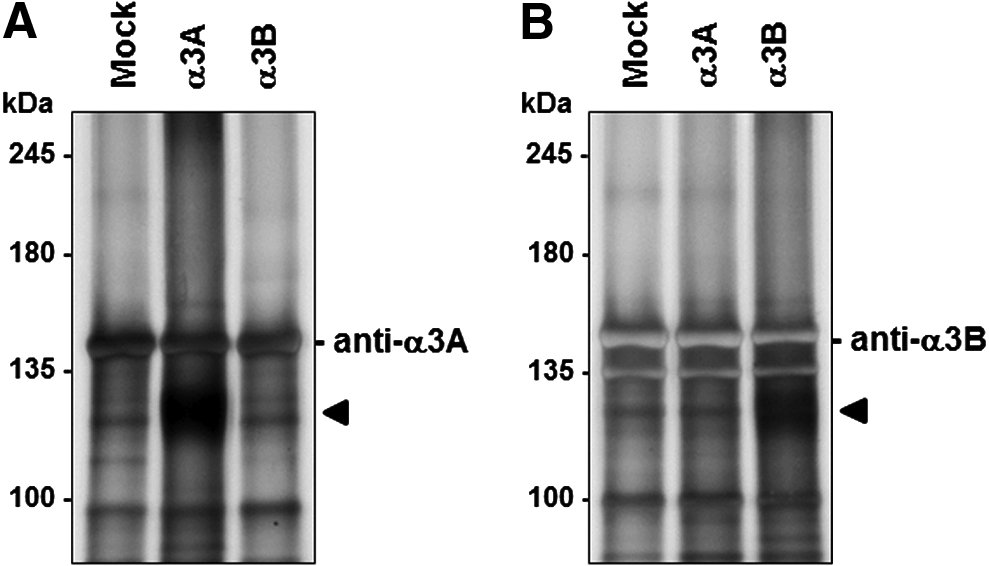
Immunoprecipitation analysis with splice variant-specific antibodies against α3 integrin. α3A and α3B integrin variants were immunoprecipitated from the lysates of transfected HEK293FT cells (mock, α3A or α3B) by Dynabeads protein G conjugated with α3A or α3B integrin variant-specific antibody. Immunoprecipitates were separated by SDS-PAGE (5% polyacrylamide gel) and analyzed for proteins by the silver staining method.
Discussion
In this study, we demonstrated the expression of α3 integrin splicing variants in the human brain. However, only very limited information is available on the functions and tissue distribution of individual splice variants of α3 integrin. It is thus hoped that the novel antibodies developed in this study will be applicable to future analyses of α3 integrin splicing variants at the protein level. In our previous study, we constructed cDNA for the α3B variant through DNA manipulation with the deletion of exon 25 from α3A and expressed the α3B variant in cultured cells.(22) Thus far, however, we have been unable to observe any substantial difference in adhesive characteristics between the α3A- and α3B-expressing cells. These results may not be particularly surprising, since both variants possess an identical extracellular domain. However, it is possible that these splice variants associate with different proteins in the cytoplasm and are involved in different signal transduction pathways. Since it is suggested that glycosylation of α3 integrin modulates its functions,(23,24) the difference in the glycosylation profiles would seem to be of interest. Moreover, it was also suggested that the α3 integrin subunit mRNA was processed in the brain by unknown mechanisms, since the α3B variant was detected only in the brain. Although the α3 integrin has been suggested to be closely linked to cancer invasion and metastasis, we did not detect an α3B variant in the cancer cells tested in this study.
A recent report has suggested that the integrin α4B variant acts as a dominant negative mutant and suppresses the function of the authentic integrin α4 subunit.(9) There is a possibility that the α3B variant similarly controls the functions of α3 integrin. Both subunits might cooperatively transduce intracellular signaling processes. In a future study, we will apply the novel splice variant-specific antibodies to α3A/3B integrins for the analysis of their physiological and pathological roles.
Footnotes
Acknowledgments
We thank Mr. Hiroki Kenmotsu and Mr. Shotaro Yasuda for their excellent technical assistance. This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan and by Hoshi University Otani Research Grants.
Author Disclosure Statement
The authors have no financial interests to disclose.