
Abstract
To produce monoclonal antibodies (MAbs) against NS1 protein of duck Tembusu virus (TMUV), the NS1 gene sequence was cloned into the prokaryotic expression vector pET-28a(+). The prokaryotic protein NS1 was successfully expressed in BL21 (DE3) and used as the immunogen in mice. Six MAbs against NS1 protein were obtained by using the standard hybridoma technique. All MAbs can react with the denatured NS1 protein in the Western blot assay and the native NS1 protein from the TMUV-infected BHK-21 cells in the immunofluorescence assay. The ELISA titers of the cell supernatants and ascites of MAbs were at a high level. The subtypes of the MAbs were determined by the Rapid Mouse Isotyping Kit-Gold series. Six MAbs possessed higher specificity and sensitivity, which indicated that MAbs against NS1 protein of TMUV may be used as valuable tools for analysis of the protein functions and pathogenesis of TMUV.
Introduction
D
TMUV is a mosquito-borne Flavivirus of the Ntaya virus group, which belongs to the Flavivirus genus within the Flaviviridae family.(9) The virus has a 10,990 nt single-stranded, positive-sense RNA genome including an open reading frame (ORF) of 10,278 nt.(10) The ORF encodes a single polyprotein, which contains three structural proteins (C, prM, and E) and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5).(11) Among them, NS1 protein is a glycoprotein that is closely related to the membrane function and may play an important role in the early replication, assembly, and release of the virus.(12,13) NS1 protein is involved in the immune response to viral infection, which contains multiple T cell and B cell epitopes and can induce the body to develop non-neutralizing protective antibodies.(14,15) NS1 protein does not mediate antibody-dependent enhancement, which provides a valuable tool for development of subunit vaccines, preparation of protective antibodies, and protein functions.(4)
In this study, six monoclonal antibodies against NS1 protein were acquired through a series of screening and subcloning, which can provide an effective tool for further studying NS1 protein function, viral pathogenic mechanism, and control measures of the virus.
Materials and Methods
Virus and RNA extraction
The SDSG strain of duck Tembusu virus was isolated from a duck farm in Shandong Province in 2013. The total viral RNA was extracted with Trizol (TransGen, Beijing, China) according to the manufacturer's instructions, and complementary DNA (cDNA) was synthesized using a PrimeScript first strand cDNA Synthesis Kit (TaKaRa, DaLian, China).
Design of primer
The primers were designed to amplify NS1 gene sequence using Primer 5.0 software according to the gene sequences in GenBank (KM102539.1, KP742476.1, KM275941.1, KJ740748.1, KJ740747.1). The primers were as follows: NS1-F, 5′-CG
Prokaryotic expression and purification of NS1 protein
The NS1 gene fragment was amplified with duck TMUV cDNA templates. After agarose gel purification and restriction enzyme digestion, the target fragment was cloned into the PET-28a(+) expression vector. The recombinant plasmid was named PET-28-NS1 and identified by enzyme digestion and DNA sequencing. Subsequently, the recombinant plasmid PET-28-NS1 and the control plasmid PET-28a(+) were transformed into BL21 (DE3) cells and induced with isopropyl-thiogalac-topyranoside (IPTG) for 1–6 h. After centrifugation, the bacterial cells were washed with phosphate buffered saline (PBS, pH 7.4) and lysed by sonication. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to analyze the recombinant expression protein. The target protein was purified by washing in different concentrations of urea solutions,(22) and the concentration was measured using a BSA Protein Assay Kit (Kang Wei, China). Finally, the protein was divided into small aliquots and stored in −80°C.
Development of indirect ELISA
To determine the optimal antigen coating concentration and serum concentration, a checkerboard titration was carried out. Each well of 96-well plates was coated with different amounts of purified NS1 protein (ranging from 400 to 25 ng/well), diluted in 0.1 M carbonate buffer (pH 9.6), and incubated at 4°C overnight. After washing three times with PBST (0.5% Tween-20 in PBS), the plates were blocked with 5% skimmed milk at 37°C for 2 h. Followed by three washes with PBST, positive and negative serum samples, serially diluted two-fold from 1:10 to 1:1280, were respectively added into the wells and incubated at 37°C for 1 h. After the plates were rinsed three times with PBST, a goat anti-mouse HRP-conjugated polyclonal serum, diluted 1:5000 with 5% skimmed milk (TransGen, FS101-02, Beijing, China), was added and incubated for 1 h at 37°C. After washing, TMB was added into the plates as the substrate for HRP and incubated in dark for 15 min. Two M H2SO4 were used to stop the reaction and data of wells were recorded by Microplate Absorbance Reader (BioTek, Winooski, VT). The antigen coating concentration and test serum concentration were determined according to the maximum ratio of P/N (positive/negative).
Monoclonal antibody production
The MAbs against NS1 protein of TMUV were generated as previously described.(16) Six-week-old female SPF BALB/c mice were inoculated subcutaneously with the purified NS1 protein (100 μg) emulsified with Freund's complete adjuvant. Later, mice were boosted twice with the same dose of immunogen and Freund's incomplete adjuvant at 2-week intervals. After three immunizations, the mouse serum was detected. If the serum was positive, the mice were boosted with 200 μg of the purified NS1 protein. Splenocytes from the immunized mice were fused with sp2/0 murine myeloma cells by using 50% polyethylene glycol (PEG 4000, Sigma) on the third day after the last immunization. The fusion cells were separated into the 96-well plates and screened in HAT medium (DMEM medium containing 10% fetal bovine serum, 100 mg/mL streptomycin, 100 IU/mL penicillin, 100 mM hypoxanthine, 16 mM thymidine, and 400 mM aminopterin). On day 5, 50 μL of HAT medium were added in each cell and HAT medium was replaced with fresh HT medium in each cell on day 12. After HAT medium screening, cell supernatants were detected for antibody reactivity and specificity by using the indirect enzyme-linked immunosorbent assay (ELISA) and Western blotting. The positive hybridoma cells were subcloned by limiting dilution method until monoclonal hybridoma cells were acquired. Positive hybridoma cells were cultured in the abdominal cavity of paraffin-primed BALB/c mice to obtain ascitic fluid.
Western blot analysis
The prokaryotic expression protein was subjected to gel electrophoresis on 12% SDS-PAGE after denaturation with 2× SDS loading buffer at 100°C for 3 to 5 min. The protein bands were transferred onto a nitrocellulose membrane, and the non-specific antibody binding sites were blocked with 5% skimmed milk in PBST (0.5% Tween-20 in PBS) at 37°C for 2 h. After washing three times in PBST, the membranes were immersed in the monoclonal antibodies against NS1 protein and incubated at 37°C for 1 h. After three washes with PBST, the membranes were incubated with a goat anti-mouse HRP-conjugated polyclonal serum diluted 1:5000 with 5% skimmed milk (TransGen, FS101-02) for 1 h at 37°C. After washing in the same manner, the membranes were immersed in DAB color liquid in dark for 1 to 3 min. Running water was used to stop the reaction.
Immunofluorescence assay
BHK-21 cells were separated into a 24-well plate (Costar Corning, Corning, NY) and inoculated with TMUV (103.5 ELD50/0.2 mL duck embryo allantoic fluid). When approximately 80% cytopathic effect (CPE) of cells occurred, the cells were fixed with methanol and acetone (1:1) for 15 min at −20°C. After washing, the monoclonal antibody against NS1 protein of TMUV diluted 1:100 with PBS was added and incubated for 1 h at 37°C. After washing three times, cells were inoculated with a goat anti-mouse IgG(H+L) with FITC conjugate (TransGen) for 1 h at 37°C. Finally, the reaction result was observed under the fluorescence microscope (Nikon, Eclipse, TE2000-S, Tokyo, Japan). BHK-21 cells without inoculation with the virus were used as the control.
Identification of MAb subtype
The Rapid Mouse Isotyping Kit-Gold series (RayBiotech, Norcross, GA) were used to identify the MAb subtypes according to the manufacturer's instructions.
Results
Prokaryotic expression and purification of NS1 protein
The recombinant plasmid PET-28-NS1 was transformed into BL21 cells and induced with 1.0 mmol/L IPTG at 37°C for 5 h. SDS-PAGE showed that the expressed protein was about 46 kDa and existed in inclusion bodies, which was consistent with the molecular weight of fusion protein His-NS1 (Fig 1). Inclusion bodies were washed by different concentrations of urea solutions,(22) and the prokaryotic expression protein NS1 was effectively purified. The purified NS1 protein was dissolved in 8 mol/L urea solution. The concentration was 4.2 μg/μL, which was measured by using a BSA Protein Assay Kit.
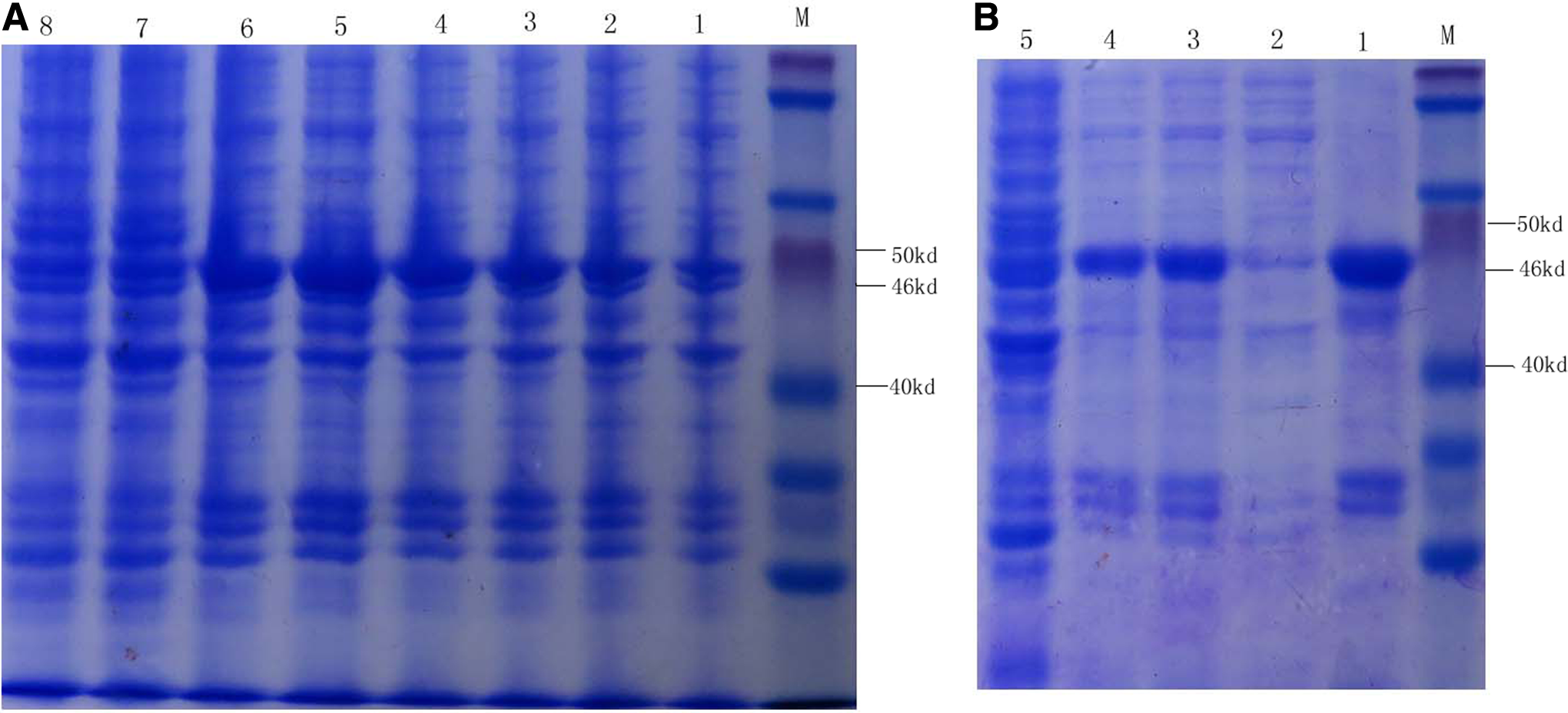
SDS-PAGE analysis of recombinant protein NS1 of TMUV. Bacteria containing PET-NS1 were induced by IPTG, and recombinant protein NS1 was purified by serial concentrations of urea solutions. Results were analyzed by SDS-PAGE. (
Generation of MAbs against NS1 protein
Six hybridoma cell lines were acquired by screening and subcloning in this study. The hybridoma cell lines were named 2A9, 2C5, 2H4, 3C5, 3G2, 4H4, respectively.
Indirect ELISA and MAb titers
The indirect ELISA assay was developed successfully in order to screen and identify the MAbs against NS1 protein. Each well of 96-well plates was coated with 100 ng of the purified NS1 protein. The ELISA titers of cell supernatants of 2A9, 2C5, 2H4, 3C5, 3G2, and 4H4 MAbs were 1:160, 1:160, 1:5000, 1:2560, 1:1280, and 1:300, respectively (Table 1). The ELISA titers of ascites of 2A9, 2C5, 2H4, 3C5, 3G2, and 4H4 MAbs were 1:12,800, 1:25,600, 1:81,920, 1:102,400, 1:204,800, and 1:25,600, respectively (Table 1).
TMUV, duck Tembusu virus.
Western blot analysis
The specificity of MAbs was analyzed by using the western blot assay. The results showed that there was a specific binding reaction between the six MAbs (2A9, 2C5, 2H4, 3C5, 3G2, and 4H4) and the NS1 protein of TMUV, respectively. No reaction was observed between the MAbs and the pET-28a protein (Fig. 2).
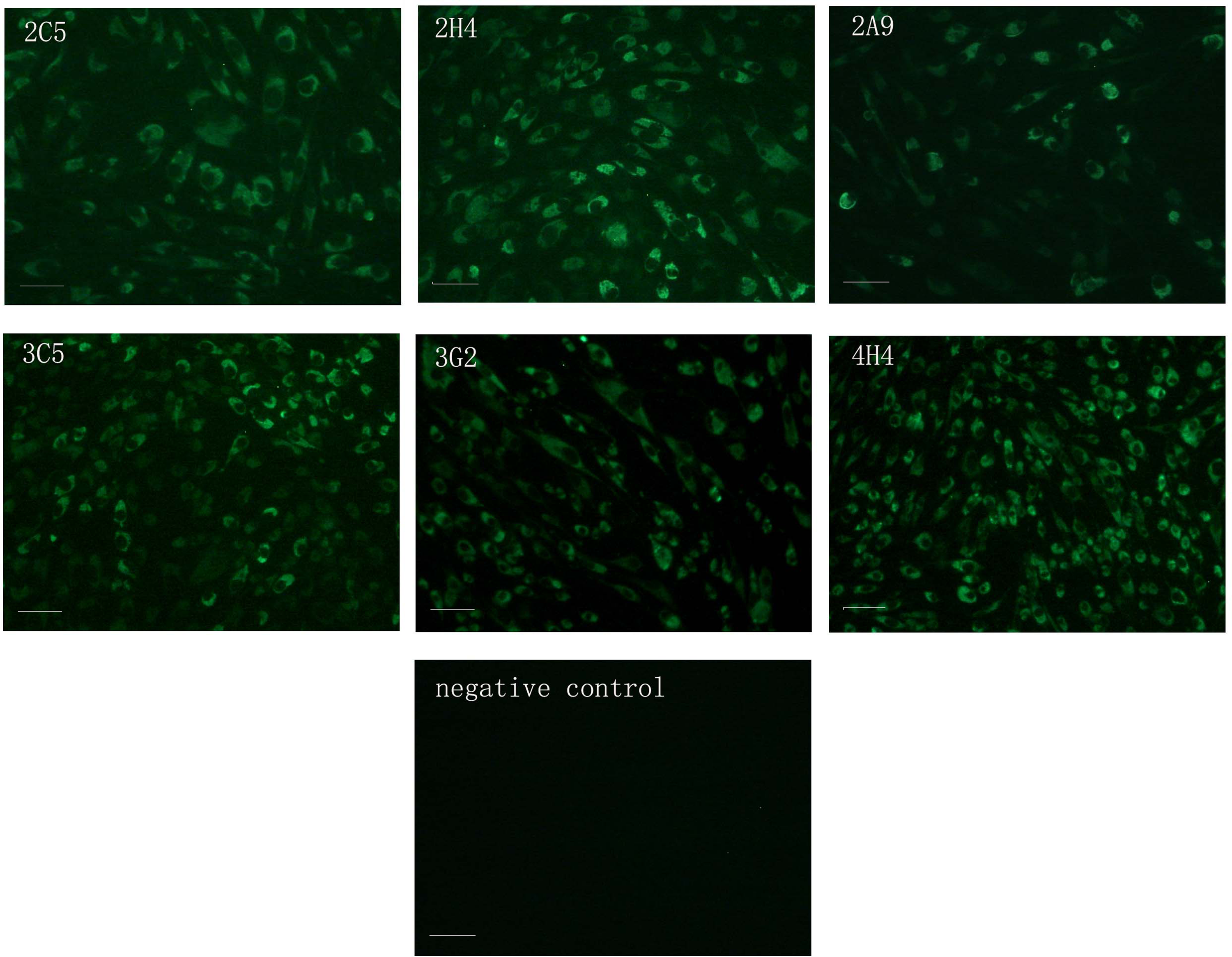
Identification of six MAbs by IFA. Monoclonal antibody against TMUV NS1 protein was used to perform IFA on TMUV-infected BHK-21 cells. BHK-21 cells infected with TMUV yielded significant fluorescence with six MAbs in the cytoplasm. Control BHK-21 cells did not yield fluorescence. Bars, 100 μm.
Immunofluorescence assay
The virus-infected BHK-21 cells yielded immunofluorescence in the cytoplasm with the six MAbs (2A9, 2C5, 2H4, 3C5, 3G2, and 4H4) (Fig. 3). The control cells did not produce any immunofluorescence (Fig. 3).

Western blot analysis of monoclonal antibodies against NS1 protein. To identify the specificity of six MAbs, Western blot was conducted with the purified fusion protein. Six MAbs were able to bind NS1 protein of TMUV specifically.
Identification of MAb subtype
The subtypes of the MAbs were identified by using a Rapid Mouse Isotyping Kit-Gold series. The results showed that two MAbs belonged to the subtype of IgG1, two MAbs belonged to the subtype of IgG2a, and two belonged to the subtype of IgG2b (Table 1). The light chains of six MAbs were all kappa (Table 1).
Discussion
In the present study, the purified prokaryotic expression protein was used as the immunogen of BALB/c mice in order to produce monoclonal antibodies against NS1 protein. Six MAbs were obtained by using ELISA, which showed a higher specificity and sensitivity.
TMUV is a member of the Falvivirus genus, which includes Japanese encephalitis virus, dengue virus, West Nile virus, yellow fever virus.(1,9,17) NS1 protein of TMUV is a non-structural protein, containing 1056 nt in length and encoding 352 aa. The molecular weight is about 40 kDa. NS1 protein is not only expressed on the virus-infected cell surface but also secreted into the environment outside the cells.(18) NS1 protein can induce the body to produce antibodies earlier, which can be used for the early diagnosis of TMUV infection.(4) The antibodies against non-structural proteins are only present in the virus-infected hosts but not in the hosts inoculated with inactivated vaccine, so it can be used to distinguish the live virus-infected animals and inactivated virus-immunized animals.(19) Thus far monoclonal antibodies against the NS1 protein of Japanese encephalitis virus have been developed, and five B-cell epitopes have been screened by using them, which lays the foundation for further study of the antigenic structure of NS1 protein and the potential clinical application of epitope-based methods.(20) The development of monoclonal antibodies against NS1 protein of dengue virus has provided an effective tool for early diagnosis and classification of dengue virus infections clinically. The function of NS1 protein and mechanism of protein-protein interaction, once ascertained, may ultimately lead to the production of effective antiviral agents. In this study, monoclonal antibodies against TMUV NS1 protein were successfully developed, which can provide an effective tool for further research of the NS1 protein function, viral pathogenic mechanism, and protein-protein interaction.(21)
In the present study, six MAbs against NS1 protein generated strong positive signals by Western blot assay and IFA, which showed a high specificity of the six MAbs. The ELISA titers in cell supernatants and ascites were at a higher level. In particular, in the 2H4, 3C5, 3G2 MAbs, the titers of ascites were about 105, which showed a high sensitivity. In conclusion, six MAbs against the NS1 protein of TMUV were developed and characterized in this study. These six MAbs may be used as powerful tools for study of replication mechanism of TMUV, screening of B epitopes of NS1 protein, and establishment of serological detection methods.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (31272583), the China Agriculture Research System (CARS-43-34), the Modern Agro-industry Technology Research System (nycytx-45-11), and the Shandong Modern Agro-industry Technology Research System (SDAIT-13-011-05).
Author Disclosure Statement
The authors have no financial interests to disclose.