
Abstract
Monoclonal antibodies (MAbs) provide scope for the development of better therapeutics and diagnostic tools. Herein, we describe the binding and neutralization profile(s) for a panel of murine MAbs generated against influenza A H1N1 viruses elicited by immunization with pandemic H1 recombinant hemagglutinin (rHA)/whole virus or seasonal H1 rHA. Neutralizing MAbs, MA-2070 and MA-M, were obtained after pandemic A/California/07/2009 (H1N1) virus/rHA immunization(s). Both MAbs reacted specifically with rHA from A/California/07/2009 and A/England/195/2009 in ELISA. MA-2070 bound rHA of A/California/07/2009 with high affinity (KD = 51.36 ± 9.20 nM) and exhibited potent in vitro neutralization (IC50 = 2.50 μg/mL). MA-2070 bound within the stem domain of HA. MA-M exhibited both hemagglutination inhibition (HI, 1.50 μg/mL) and in vitro neutralization (IC50 = 0.66 μg/mL) activity against the pandemic A/California/07/2009 virus and showed higher binding affinity (KD = 9.80 ± 0.67 nM) than MA-2070. MAb, MA-H generated against the seasonal A/Solomon Islands/03/2006 (H1N1) rHA binds within the head domain and bound the seasonal H1N1 (A/Solomon Islands/03/2006 and A/New Caledonia/20/1990) rHAs with high affinity (KD; 0.72–8.23 nM). MA-H showed high HI (2.50 μg/mL) and in vitro neutralization (IC50 = 2.61 μg/mL) activity against the A/Solomon Islands/03/2006 virus. All 3 MAbs failed to react in ELISA with rHA from various strains of H2N2, H3N2, H5N1, H7N9, and influenza virus B, suggesting their specificity for either pandemic or seasonal H1N1 influenza virus. The MAbs reported here may be useful in developing diagnostic assays.
Introduction
I
Majority of the infections due to influenza A virus are uncomplicated and resolve within a week. However, some infections may progress to complications such as severe pneumonia and multi-organ failure; as was also observed in individuals with pandemic H1N1 (pH1N1) or H5N1 infections.(5,7,8) Further, antiviral resistance has been observed in almost all seasonal H1N1 strains to oseltamivir whereas about 1% of 2009 pH1N1 viruses were oseltamivir resistant.(9) Antiviral resistance acquisition poses a serious threat to human health and calls for regular monitoring of influenza virus infections. Hence, there is an urgent need to develop alternate therapeutic and diagnostic tools that can be effective against divergent influenza virus strains and readily accessible during a pandemic outbreak. The development of sensitive diagnostic tools will also be useful for surveillance purposes.
Several studies suggest that influenza virus neutralizing monoclonal antibodies (MAbs) to HA protein can be effective for therapeutic and diagnostic purposes.(10–12) Hence, there is considerable promise in the development of high affinity neutralizing antibodies against influenza virus HA. In this direction, we have been able to isolate murine MAbs against influenza A viruses that bind HA with high affinity and mediate neutralization via distinct mechanism(s).
Materials and Methods
Cell lines, antigens, and virus
Madine Darby Canine Kidney (MDCK) cell line (CCL-34) was obtained from American Type Culture Collection (Manassas, VA) and cultured in Dulbecco's modified Eagle's medium (Sigma Aldrich, Inc., St. Louis, MO). Sp2/O mouse myeloma cell line was obtained from National Centre for Cell Science (Pune, India) and cultured in RPMI-1640 medium (Sigma Aldrich, Inc.). Media were supplemented with 10% fetal bovine serum and an antibiotic–antimycotic cocktail [penicillin (100 U/mL), streptomycin (100 μg/mL), and amphotericin B (0.25 μg/mL); Biological Industries, Kibbutz beit, Haemek, Israel]. Both the cell lines were cultured at 37°C under humidified conditions with 5% CO2.
Full-length mammalian cell-expressed recombinant HA (rHA) proteins were purchased from Sino Biologicals, Inc. (Beijing, China). A list of rHA proteins used in this study corresponding to various viruses is given here: A/California/07/2009 (pH1N1), A/England/195/2009 (pH1N1), A/Solomon Islands/03/2006 (H1N1), A/New Caledonia/20/1999 (H1N1), A/Puerto Rico/8/1934 (H1N1), A/Canada/720/2005 (H2N2), A/Aichi/2/1968 (H3N2), A/Perth/16/2009 (H3N2), A/turkey/Turkey/1/2005 (H5N1), A/Hong Kong/213/2003 (H5N1), A/Bar headed goose/Qinghai/14/2008 (H5N1), A/Anhui/1/2013 (H7N9), and influenza virus B, B/Brisbane/60/2008.
pH1N1 NYMCX-179A (A/California/07/2009: Influenza virus infectious NYMCX-179A, NIBSC Code 09/124) and seasonal H1N1 (A/Solomon Islands/03/2006: Influenza virus infectious IVR-145, NIBSC code 07/144) received from NIBSC, UK and passaged in MDCK cells in presence of TPCK-trypsin (2 μg/mL; Sigma Aldrich, Inc.) were obtained from Serum Institute of India Limited (Pune, India). Titer of the virus stock was calculated using Reed and Muench method.(13)
Generation of MAbs
Female BALB/cJ mice (8–10 weeks old; Small Animal Experimental Facility, National Institute of Immunology, New Delhi, India) were immunized subcutaneously either with 7.5 μg equivalent of HA of inactivated, alum adsorbed strain of pH1N1 virus (A/California/07/2009) (Serum Institute of India Limited) or 50 μg of pH1N1 (A/California/07/2009) or seasonal H1N1 (A/Solomon Islands/03/2006) rHA protein along with the adjuvant, Montanide Pet Gel A (Seppic, Puteaux Cedex, France). Animals were boosted intraperitoneally two times at 4 week intervals with the same amount of antigen. The animals were kept in the conventional containment conditions and fed ad libitum. All the experiments were performed after due approval from the Institutional Animal Ethical Committee, National Institute of Immunology, New Delhi (IAEC#247/10) following its guidelines. Immunized animals were used to generate MAbs as per the protocol described elsewhere.(14) Hybrid cell clones secreting MAbs reacting with rHA from either pandemic A/California/07/2009 or seasonal A/Solomon Islands/03/2006 were identified by screening culture supernatants in ELISA as described later in this section and single cell clones were obtained by two to three rounds of limiting dilution. Specific hybrid cell clones secreting MAb neutralizing pandemic A/California/07/2009 or seasonal A/Solomon Islands/03/2006 viruses were grown as ascites in BALB/cJ mice primed with Pristane (2,6,10,14–tetramethylpentadecane; Sigma Aldrich, Inc.). The MAbs were purified from ascites using Protein-G Sepharose (GE Healthcare Biosciences AB, Uppsala, Sweden) as per the manufacturer's instructions. Isotyping of the antibodies was carried out using murine MAb isotyping kit (Sigma Aldrich, Inc.) by indirect ELISA as per the instructions provided with the kit.
ELISA reactivity studies
The reactivity of the purified MAbs in ELISA was determined against rHA proteins corresponding to various influenza viruses essentially as described before.(14) In brief, 96-well plates (Greiner Bio-One, GmbH, Frickenhausen, Germany) were coated with rHA proteins (160 ng/well; Sino Biological, Inc.) and ovalbumin (200 ng/well; as negative control) and ELISA plates further processed to study the reactivity of MAbs to the coated antigen as described previously.(14) The optical density recorded for the ovalbumin wells were subtracted from the optical densities of the respective test wells and results are expressed as mean of duplicate readings.
Binding affinity studies using surface plasmon resonance
The binding affinity of purified MAbs (ligand) to recombinant protein(s) (analyte) was determined by surface plasmon resonance (SPR) experiments performed with a Biacore 3000 optical biosensor (Biacore, Uppsala, Sweden) at 25°C. MAb at 600 resonance units (RU) was immobilized on an activated sensor channel of a research-grade CM5 chip (GE HealthCare Biosciences AB) by amine coupling. Ovalbumin was immobilized on another sensor channel that served as a negative control for each binding interaction. A concentration (trimer) series of the analyte(s) in the range 12.5–200 nM were passed over each channel to determine the equilibrium dissociation constant (KD). The analyte dilutions were made in a running buffer of phosphate-buffered saline (PBS; pH 7.4) containing 0.05% P20 surfactant. The binding experiments were performed at a flow rate of 30 μL/min. The sensor surfaces were regenerated with 4 M MgCl2 after every binding event. Each binding curve was analyzed after correcting for nonspecific interaction measured by signals obtained in the negative-control flow channel. The kinetic parameters of binding (mean ± standard deviation) were obtained by fitting the data to the simple 1:1 Langmuir interaction model using BIA EVALUATION 3.1 software. The binding experiments were repeated at least thrice.
Antibody competition using SPR
The competition between MAbs binding to rHA of A/California/07/2009 was monitored by SPR experiments performed using a Biacore 2000 optical biosensor (Biacore) at 25°C. The broadly neutralizing human Mab (bnCR6261)(15) was immobilized at 500 RU on an activated sensor channel of a research-grade CM5 chip (GE HealthCare Biosciences AB) by amine coupling. The binding of rHA from A/California/07/2009 to immobilized bnCR6261 was monitored in the absence and presence of MA-2070, MA-M and bnCR6261.
Antibody pull-down assay
MA-2070 binding to pH1HA10-Foldon, a “headless” HA stem mimetic(16) was probed in an IgG pull-down assay using Protein-G beads (GE HealthCare Biosciences AB). pH1HA10-Foldon (6.5 μM) was incubated with MA-2070 (IgG1) (2.5 μM) for 2 hours at 4°C. This was followed by the addition of Protein-G beads (10 μL) and incubation for 1 hour at 4°C under mild mixing conditions to capture the antibody. The beads were spun down at 3000 g for 10 minutes (4°C) and the supernatant was collected. The beads were washed with PBS (pH 7.4). Next, the antibody bound to the beads was eluted with 10 μL of 100 mM glycine.HCl, pH 3.0 and neutralized with 2.5 μL of 1 M Tris.HCl, pH 9.0. The unbound and eluted fractions were analyzed using a 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis under nonreducing conditions and resolved proteins stained with Coomassie brilliant blue (Sigma Aldrich, Inc.).
In vitro microneutralization assay
In vitro microneutralization assay was performed using MDCK cell line as per the WHO guidelines, as described previously.(17) MDCK cells (1.5 × 104/well) were seeded in a 96-well cell culture plate (Greiner Bio-One, GmbH) and incubated overnight under standard culture conditions. The next day, an equal volume of purified antibody at varying concentrations and 100 TCID50 units of the pandemic A/California/07/2009 (H1N1) or seasonal A/Solomon Islands/03/2006 (H1N1) viruses were incubated for 1 hour at room temperature. After 1 hour, virus-antibody mixture was added onto the seeded MDCK cells and plates were further incubated for 24 hours at 32/37°C under humidified atmosphere and 5% CO2. Subsequently, cells were fixed with 80% acetone and probed with 1:4000 dilution of anti-influenza A–nucleoprotein antibody (Millipore, Billerica, MA). After three rounds of washing with PBST, the plates were incubated with 1:10,000 dilution of HRP conjugated goat-anti-mouse antibody (Pierce Biotechnology, Inc., Rockford, IL) and incubated for 1 hour at 37°C. After washing three times with PBST, reaction was developed as mentioned above for ELISA. The 50% inhibitory concentration (IC50) of the neutralizing antibody was calculated using the nonlinear regression program of GraphPad Prism software (Version 6.0; GraphPad Software, La Jolla California, USA).
Hemagglutination inhibition assay
Hemagglutination inhibition (HI) assay was performed according to the WHO guidelines using four HA units of the influenza virus and 0.5% guinea pig red blood cells (RBCs) as described previously.(17) The HI titer of the MAb represents the lowest concentration of the antibody showing HI activity.
Results
Isolation and ELISA reactivity profile of MAbs against H1N1 influenza HA
MAb, MA-2070 (IgG1) generated by immunization with pandemic A/California/07/2009 (H1N1) virus bound homologous pH1N1 rHA in ELISA even at low concentration of the MAb (100 ng/mL). In addition, it also showed binding with rHA of pandemic A/England/195/2009 (H1N1) virus (Fig. 1a). MA-2070 showed high affinity for binding with rHA from A/California/07/2009 in SPR (Fig. 1b and Table 1). MA-2070 did not react in ELISA with rHA from A/Solomon Islands/03/2006 (H1N1), A/New Caledonia/20/1999 (H1N1), A/Puerto Rico/8/1934 (H1N1), A/Canada/720/2005 (H2N2), A/Aichi/2/1968 (H3N2), A/Perth/16/2009 (H3N2), A/turkey/Turkey/1/2005 (H5N1), A/Hong Kong/213/2003 (H5N1), A/Bar headed goose/Qinghai/14/2008 (H5N1), A/Anhui/1/2013 (H7N9), and influenza virus B, B/Brisbane/60/2008 (data not shown). The MAb, MA-M (IgG1) isolated after pandemic A/California/07/2009 rHA protein immunization also exhibited similar reactivity pattern in ELISA as observed with MA-2070 but showed higher affinity (KD = 9.80 ± 0.67 nM) than MA-2070 (KD = 51.36 ± 9.20 nM; Fig. 2a, b and Table 1).
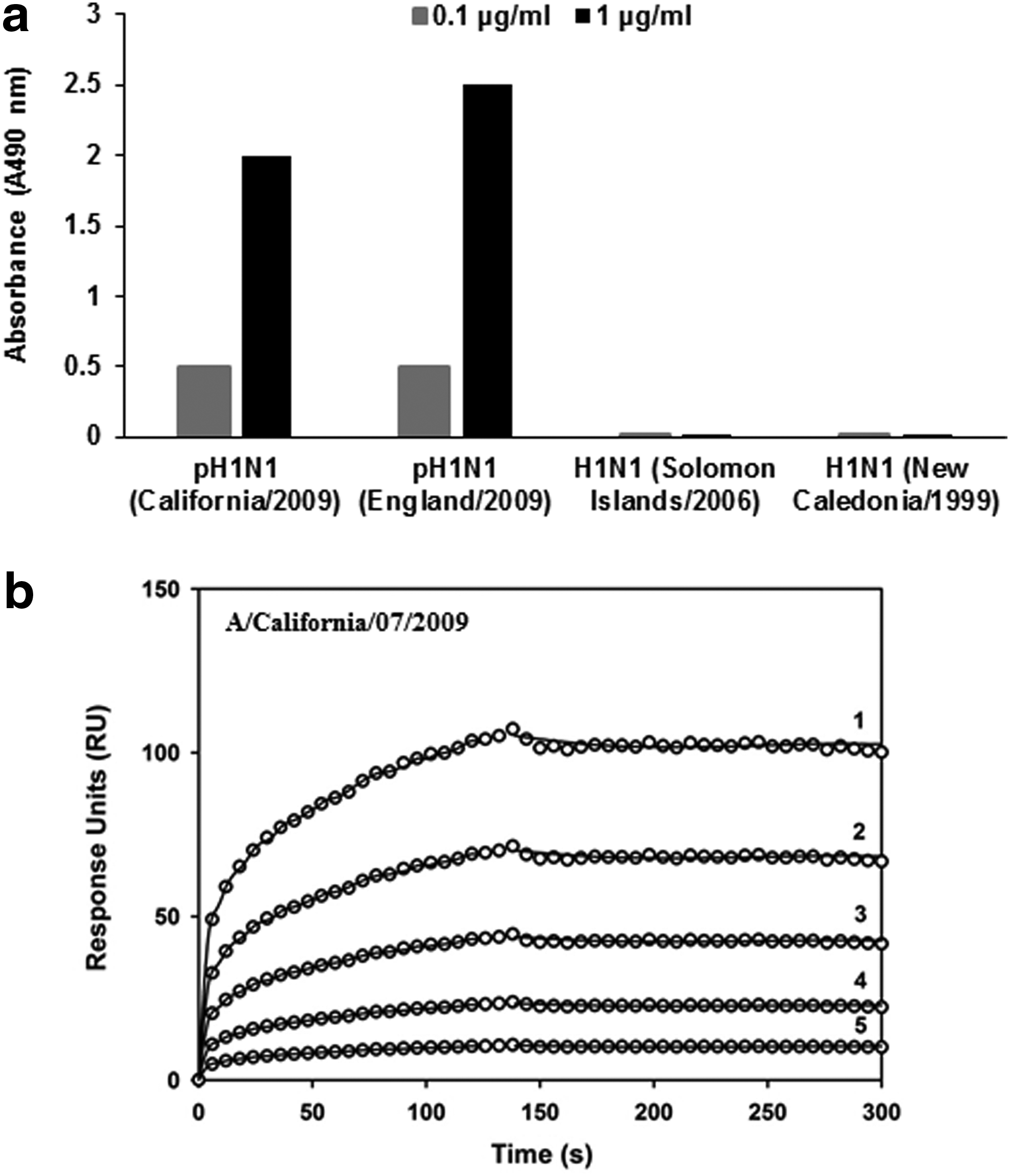
MA-2070 generated against pandemic A/California/07/2009 (H1N1) virus binds rHA protein with high affinity.
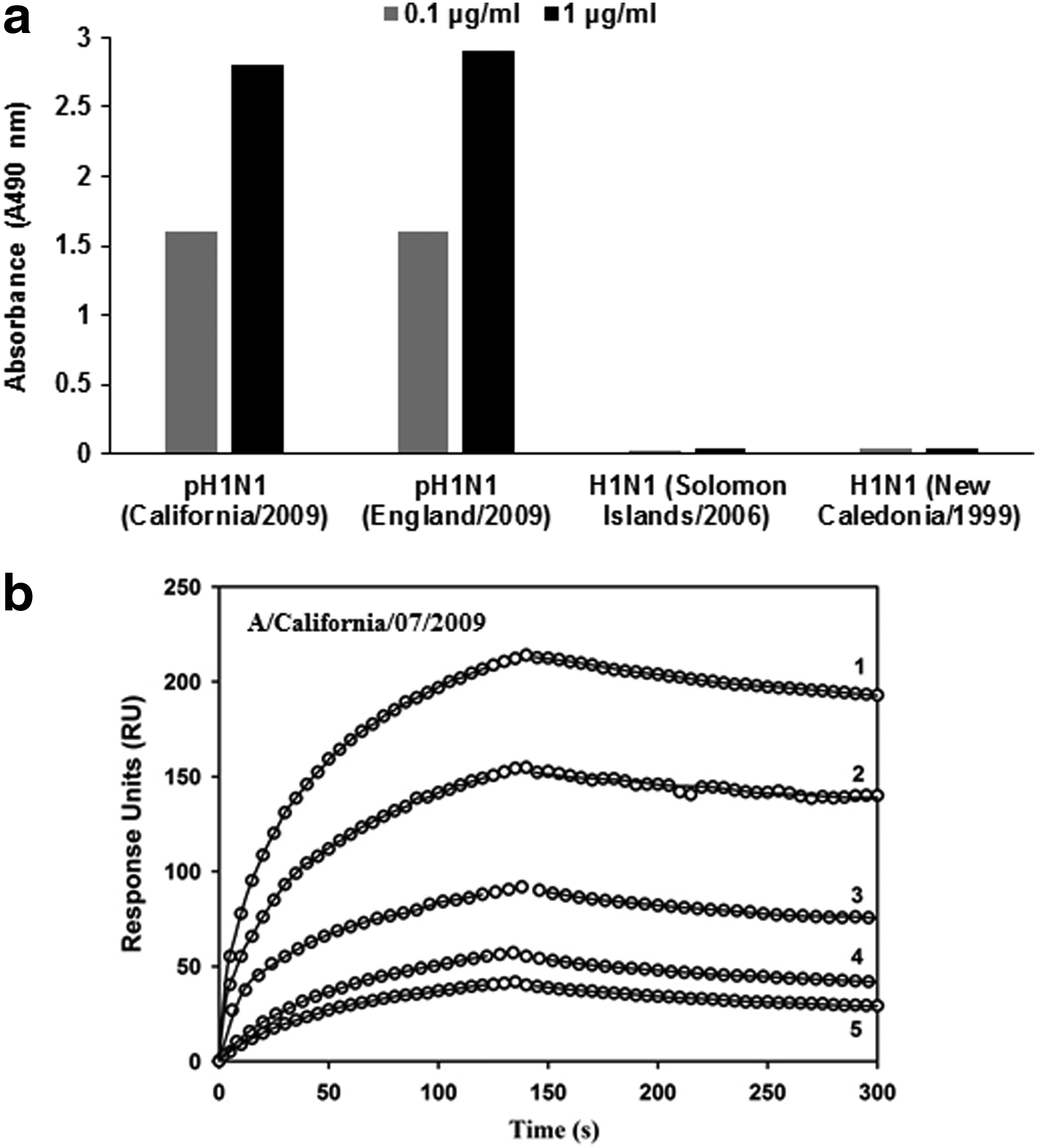
Binding profile of MA-M generated against pandemic A/California/07/2009 (H1N1) rHA protein.
600 RUs of the indicated MAbs were immobilized on the surface of a CM5 chip. A concentration series of the analyte(s) were passed over the immobilized MAb.
Reported kinetic parameters are the mean ± standard deviation of triplicate experiments.
MAbs, monoclonal antibodies; RU, resonance units.
The MAb elicited by the seasonal A/Solomon Islands/03/2006 (H1N1) rHA protein, MA-H (IgG1) showed significant ELISA reactivity against the homologous protein even at a concentration of 100 ng/mL (Fig. 3a). Additionally, the MAb also exhibited reactivity in ELISA with rHA from A/New Caledonia/20/1999 (H1N1) virus (Fig. 3a). However, MA-H failed to show any significant reactivity with rHA from another seasonal H1N1 virus namely A/Puerto Rico/8/1934 (data not shown). It failed to react with the rHA of pH1N1 (A/California/07/2009 and A/England/195/2009) even when tested at the highest concentration of 100 μg/mL. Further, MA-H also failed to react in ELISA with rHA from A/Canada/720/2005 (H2N2), A/Aichi/2/1968 (H3N2), A/Perth/16/2009 (H3N2), A/turkey/Turkey/1/2005 (H5N1), A/Hong Kong/213/2003 (H5N1), A/Bar headed goose/Qinghai/14/2008 (H5N1), A/Anhui/1/2013 (H7N9), and influenza virus B, B/Brisbane/60/2008 (data not shown).

HA reactivity profile of MAb generated against seasonal A/Solomon Islands/03/2006 (H1N1) rHA.
The kinetic parameters for the binding of MA-H to H1 rHAs were determined by SPR. In agreement with ELISA results, MA-H bound the rHA from A/Solomon Islands/03/2006 and A/New Caledonia/20/1999 with high affinity (KD; 0.72–8.23 nM) (Fig. 3b, c and Table 1). The low off-rates (koff; ∼4 × 10−5 s−1) further indicate the formation of a stable complex.
MA-2070 prevents in vitro pandemic A/California/07/2009 (H1N1) virus infection, binds within the HA stem, and competes with the bnCR6261
MAb, MA-2070 neutralized the pandemic A/California/07/2009 (H1N1) virus in an in vitro microneutralization assay with an IC50 value of 2.50 μg/mL (Fig. 4 and Table 2). However, MA-2070 was negative in an HI assay indicating that the MAb probably targets the HA stem. We further validated our hypothesis by measuring the binding of MA-2070 to pH1HA10-Foldon, a “headless” rHA stem mimetic from pandemic A/California/07/2009.(16) The low equilibrium dissociation constant (KD = 43.5 ± 9.1 nM) of MA-2070 binding to pH1HA10-Foldon indicates the formation of a stable complex (Fig. 5a). The antigen–antibody complex between pH1HA10-Foldon and MA-2070 was also detected in an IgG-specific pull-down assay using Protein G beads (Fig. 5b).

In vitro microneutralization activity of MAbs generated against pandemic and seasonal H1N1. Varying concentrations of protein-G purified MAbs were used to evaluate in vitro neutralization of the pandemic A/California/07/2009 (H1N1) virus and seasonal A/Solomon Islands/03/2006 (H1N1) virus as described in Materials and Methods section. Neutralization curve for pandemic A/California/07/2009 (H1N1) virus by MA-2070 and MA-M and seasonal A/Solomon Islands/03/2006 (H1N1) virus by MA-H are shown. The values represent mean ± standard deviation of experiment performed in triplicate.

MA-2070 binds within the HA stem.
NS, no significant activity.
The bnCR6261 binds the stem of influenza HA and prevents virus infection.(15) We tested the ability of MA-2070 to compete with bnCR6261 by SPR. The decreased signal with rHA from A/California/07/2009 binding to immobilized bnCR6261 when co-incubated with MA-2070 suggests that both the MAbs bind to sterically overlapping regions on HA (Fig. 6).
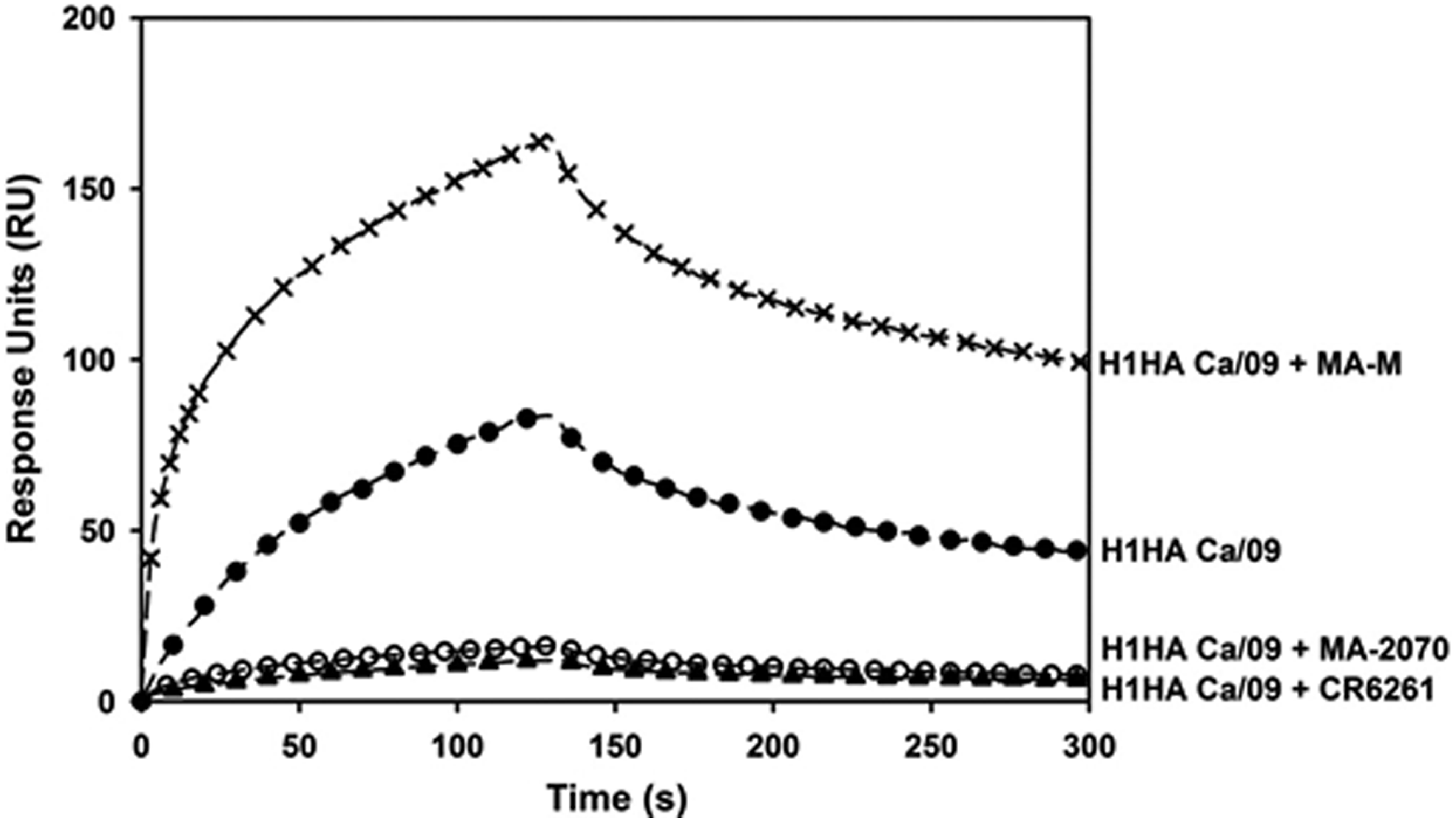
MA-2070 competes with the bnCR6261. The binding of rHA from A/California/07/2009 (H1N1) [Ca/09] to immobilized bnCR6261 was monitored by surface plasmon resonance in the absence and presence of either MA-2070 or MA-M as indicated. The decreased signal of A/California/07/2009 (H1N1) rHA binding to immobilized CR6261 when preincubated with MA-2070 suggests that both the MAbs (CR6261 and MA-2070) bind to sterically overlapping regions on HA.
MA-M and MA-H are effective in HI and in vitro virus neutralization
MA-M showed potent neutralization of the pandemic A/California/07/2009 (H1N1) virus in an in vitro microneutralization assay (IC50 = 0.66 μg/mL). MA-M also showed potent activity in an HI assay (HI titre, 1.50 μg/mL) suggesting that the antibody is probably targeting the head domain of HA (Fig. 4 and Table 2). MA-M and bnCR6261 binds rHA from A/California/07/2009 at sterically nonoverlapping regions as inferred from the binding of rHA from A/California/07/2009 virus complexed with MA-M to immobilized bnCR6261 (Fig. 6). Further, the lack of any detectable binding with pH1HA10-Foldon indicates MA-M is a head-directed antibody (data not shown). MA-H neutralized the seasonal A/Solomon Islands/03/2006 (H1N1) virus (IC50 = 2.61 μg/mL) and also inhibited hemagglutination of RBCs by the A/Solomon Islands/03/2006 virus (HI titre, 2.50 μg/mL) suggesting that it is also a head-directed antibody (Fig. 4 and Table 2).
Discussion
Considering the public health significance of the emergence of pandemic influenza virus strains, it is imperative to search for newer diagnostic and therapeutic modalities for identification and treatment of the disease. Neutralizing antibodies are the critical immunological parameter for protection against the influenza virus infection. Historically, MAbs have been fundamental tools for diagnosis and epitope mapping of influenza viruses and their importance as an alternative treatment option is also being realized. Influenza HA is the primary target of neutralizing antibodies postvaccination or natural infection and mediate protection via multiple mechanism(s).(18) It consists of a long helical chain anchored in the membrane known as stem, which is topped by a globular head domain. Herein, we attempted to isolate anti-HA neutralizing MAbs and determine their binding specificities by ELISA/SPR.
MA-2070 and MA-M react with rHA from pH1N1, A/California/04/2009 and A/England/195/2009 in ELISA, which may be due to their high (99%) sequence similarity. In addition, MAb, MA-H reacting to rHA from seasonal A/Solomon Islands/03/2006 (H1N1) virus has also been reported. MA-H also reacted in ELISA with rHA from A/New Caledonia/20/1999 but failed to react with rHA from A/Puerto Rico/8/1934. Sequence analysis revealed that among these three viruses, A/Solomon Islands/03/2006 and A/New Caledonia/20/1999 are closely related (showed 98% sequence similarity), whereas A/Puerto Rico/8/1934 had drifted apart and showed sequence similarity of 87% with both the above seasonal viruses, which might explain nonreactivity of MA-H with rHA of A/Puerto Rico/8/1934. Interestingly, all the three MAbs described in the present article failed to react with rHA from representative strains of H2N2, H3N2, H5N1, H7N9, and influenza virus B, suggesting that these MAbs react specifically to either pandemic or seasonal H1N1 influenza virus HA protein with limited breadth. All three MAbs showed low dissociation constant ranging from 0.72 to 51.36 nM suggesting that these are high affinity antibodies.
The anti-HA antibodies mediate virus neutralization via distinct mechanisms. The antigenic sites on influenza HA have been extensively characterized.(19) The antibodies targeted to the receptor binding site (RBS) in the head domain of HA inhibit virus attachment to the cell surface receptors by steric occlusion. These antibodies test positive in the HI assay. On the contrary, HA stem-directed antibodies mediate virus neutralization by inhibiting the fusion of the virus and host cell membrane. These antibodies test positive in a virus neutralization assay but negative in an HI assay. Therefore, a simple boolean read-out of the HI and virus neutralization assays in tandem can provide preliminary information regarding the antibody binding site on HA. The HI and virus neutralization assays are also the gold standard in evaluating the therapeutic potential of an MAb.
Our studies suggest that MA-2070 binds to a neutralizing epitope on the HA stem as it showed potent in vitro neutralization activity but failed to show any significant activity in HI assay. MA-2070 indeed reacts with the stem domain of HA as confirmed by its binding with pH1HA10-Foldon,(16) a stem mimetic of pH1N1, in SPR and by antibody pull-down assay. Further, MA-2070 bound to the overlapping epitope being recognized by human MAb, bnCR6261, which has been mapped to the stem domain of HA.(15) The observed immunological specificity of MA-2070 for pandemic HA is surprising since the HA stem is well conserved. Further investigations to delineate the epitope of MA-2070 at residue-specificity will aid in understanding the precise mechanism(s) that contribute to virus escape.
Both MA-M and MA-H in addition to in vitro neutralization also showed activity in the HI assay suggesting that these antibodies bind in the head domain of HA and mediate neutralization likely by inhibiting RBS. Previous studies have demonstrated that anti-HA head-directed antibodies are more potent than stem-directed antibodies.(20) MA-M, which may be head directed against pandemic A/California/07/2009 influenza virus HA indeed has higher in vitro neutralization (IC50 = 0.66 μg/mL) as compared to MA-2070 (IC50 = 2.50 μg/mL). Further, neutralizing antibodies targeting head region may be recognizing either Sa/Sb (proximal to RBS) or Ca/Cb (distal to RBS) antigenic sites. This can further affect their neutralization efficiency/potency.(21) MA-H though has higher binding affinity than MA-M, but has lower in vitro neutralization potency. Similar observations have been reported for antibodies that bind outside the virus RBS.(22) Therefore, residue level delineation of the epitope(s) for the reported antibodies is necessary to correlate the observed binding affinities and in vitro neutralization potencies. Influenza neutralizing MAbs characterized to date by an in vitro microneutralization assay have IC50 values in the range of 0.08 to >10 μg/mL.(10,17,23–25) Burioni et al., have reported pH1N1 neutralizing human MAbs with IC50 in the range of 2.8–4.0 μg/mL.(25) The three MAbs described in the present article have IC50 values in the range of 0.66–2.61 μg/mL. Neutralizing MAbs against pH1N1 virus with IC50 as low as 2.5 ng/mL have been reported.(26) However, in this case neutralization capacity was assessed by employing plaque reduction assay instead of microneutralization assay.(26) Whether the high neutralization capacity of the antibodies as observed in the above study was due to different assays being used for assessment needs to be resolved.
In conclusion, we report the isolation and characterization of high affinity MAbs against influenza A H1N1 viruses with potent in vitro neutralization activity although with limited breadth of immunological cross-reactivity.
Footnotes
Acknowledgments
This work was funded by the Department of Biotechnology (DBT), Government of India (BT/BIPP/0213/04/09) and National Institute of Immunology, New Delhi. S.K.G. would like to acknowledge Tata Innovation Fellowship awarded to him by DBT, Government of India. The funding agencies had no role in study design, collection, analysis, or interpretation of the data.
Author Disclosure Statement
No competing financial interests exist.