
Abstract
CXCR1 and CXCR2 are chemokine receptors that have different selectivity of chemokine ligands, but the distinct role of each receptor is not clearly understood. This is due to the absence of specific inhibitors in guinea pigs, which are the appropriate species for investigation of CXCR1 and CXCR2 because of their functional similarity to humans. In this study, we generated and evaluated monoclonal antibodies that specifically bound to guinea pig CXCR1 (gpCXCR1) and guinea pig CXCR2 (gpCXCR2) for acquisition of specific inhibitors. To assess the activity of antibodies, we established CHO-K1 cells stably expressing either gpCXCR1 or gpCXCR2 (CHO/gpCXCR1 or CHO/gpCXCR2). CHO/gpCXCR1 showed migration in response to guinea pig interleukin (IL)-8, and CHO/gpCXCR2 showed migration in response to both guinea pig IL-8 and guinea pig growth-regulated oncogene α. The receptor selectivities of the chemokines of guinea pigs were the same as the human orthologs. The inhibitory activities of the anti-gpCXCR1 and anti-gpCXCR2 monoclonal antibodies on cell migration were observed in a concentration-dependent manner. In conclusion, we successfully obtained inhibitory antibodies specific to gpCXCR1 and gpCXCR2. These inhibitory antibodies will be useful to clarify the physiological roles of CXCR1 and CXCR2 in guinea pigs.
Introduction
CXCR1
Guinea pigs have often been utilized for the understanding of inflammatory diseases in which IL-8, CXCR1, and CXCR2 are thought to be involved(13,14); however, the distinct functions of CXCR1 and CXCR2 have not been elucidated because their specific inhibitors did not exist. Recently, the inhibitory effect of human CXCR2 (hCXCR2) selective antagonists against gpCXCR1 and gpCXCR2 has been studied and proved to inhibit gpCXCR1 as well as gpCXCR2.(15) Some inhibitory antibodies specific to human CXCR1 (hCXCR1) and hCXCR2 exist,(16–19) but no clones have been reported to be cross-reactive to those of guinea pigs possibly due to the low identity of these receptors in extracellular domains.
In this study, we aimed to produce monoclonal antibodies against gpCXCR1 and gpCXCR2, which can inhibit their function specifically. To generate them, DNA immunization of mice was conducted because it was a suitable immunization method for multitransmembrane protein.(20) Monoclonal antibodies against gpCXCR1 and gpCXCR2 were successfully obtained and the results suggest that these antibodies selectively inhibited the migration of cells to specific ligands in a chemotaxis assay.
Materials and Methods
Construction of expression plasmid
The expression plasmids of gpCXCR1, gpCXCR2, gpIL-8, and gpGROα were constructed by the standard genetic engineering procedure. In brief, guinea pig Cxcr1 (NCBI Refseq: NM_001173416, [gpCXCR1]) and guinea pig Cxcr2 (NCBI Refseq: NM_001172875, [gpCXCR2]) genes were amplified with polymerase chain reaction from a cDNA mixture of guinea pig lung, skeletal muscle, heart, and brain and inserted into multiple cloning sites of pcDNA3.1(+), resulting in pcDNA3.1/gpCXCR1 and pcDNA3.1/gpCXCR2. Guinea pig Cxcl8 (NCBI Refseq: NM_001173399, [gpIL-8]) and guinea pig Cxcl1 (NCBI Refseq: NM_001172938, [gpGROα±]) genes were codon optimized and synthesized linking to His-tag for protein purification. These genes were inserted into multiple cloning sites of pET11d and pET22b, resulting in pET11d/gpIL-8 and pET22b/gpGROα, respectively.
Mouse immunization and establishment of hybridomas
Six-week-old female BALB/c mice were used for DNA immunization. Intramuscular injection of the expression plasmid, followed by in vivo electroporation, was conducted as reported previously.(21) In brief, lower leg muscles were pretreated with bovine hyaluronidase. Then, 50 μg of pcDNA3.1/gpCXCR1 or pcDNA3.1/gpCXCR2 was injected into the same site. Two electrode needles were inserted into the same site, and electric pulses (200 V, 50 ms, 6 times) were delivered with ECM830 (BTX, Holliston, MA). This DNA immunization protocol was repeated several times every 2 weeks. Blood samples were collected from the tail vein every 2 weeks. Antibody response of mouse serum was evaluated by flow cytometric analysis to choose a mouse that produced the anti-gpCXCR1 or gpCXCR2 antibody after DNA immunization. Lymph nodes of the selected immunized mice were harvested, and hybridomas were established by conventional hypoxanthine-aminopterin-thymidine (HAT) selection and cloning according to the ClonaCell HY (STEMCELL Technologies, Vancouver, Canada) instructions using SP2 myeloma as fusion partner. All experimental procedures were performed in accordance with the in-house guidelines of the Institutional Animal Care and Use Committee of Daiichi Sankyo Co., Ltd.
Monoclonal antibody production and purification
Culture supernatants of the hybridoma clones were used for flow cytometric screening to pick up clones binding to gpCXCR1 or gpCXCR2. Positive hybridomas were expanded, and media were exchanged to Hybridoma SFM (Thermo Fisher Scientific, Waltham, MA) supplemented with low IgG fetal bovine serum (FBS) 20%. After 6 days of culturing, the supernatant containing antibodies was collected and filtered. The antibodies were purified using a HiTrap protein G column (GE Healthcare, Buckinghamshire, United Kingdom), followed by desalting with PD-10 column (GE Healthcare). The antibodies were concentrated to ∼5 mg/mL with Amicon Ultra (Merck Millipore, Darmstadt, Germany). Concentration and purity were determined by high-performance liquid chromatography (HPLC) (Agilent Technologies, Santa Clara, CA). Endotoxin level was determined using an Endosafe®-PTS™ (Charles River Laboratories, Wilmington, MA). Isotyping of the antibodies was conducted by using the mouse monoclonal antibody isotyping test kit (Bio-Rad, Hercules, CA).
Chemokine production and purification
Plasmids, pET11d/gpIL-8 and pET22b/gpGROα, were transformed into Origami B strain of Escherichia coli. The expanded transformants were induced with isopropyl β-D-1-thiogalactopyranoside (IPTG) for gpIL-8 and gpGROα expression. After overnight culturing at 25°C, the transformants were collected and lysed. The lysates were purified with HisTrap HP (GE Healthcare), followed by desalting with a PD-10 column and fractionation of objective peak by reversed-phase chromatography using an ODS-120T column. The samples were lyophilized and dissolved to ∼50 μM in phosphate-buffered saline (PBS) supplemented with 0.1% bovine serum albumin (BSA) for assay. The calculated molecular sizes of the recombinant gpIL-8 and gpGROα were 11.1 and 9.7 kDa, respectively. Concentration was determined by the Bradford protein assay, and purity was confirmed by SDS-PAGE. Endotoxin level was determined using Endosafe-PTS.
Cell culture and transfection
HEK293T, human embryonic kidney cell line stably transfected with SV40 large T antigen, was cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS and 1% penicillin/streptomycin at 37°C and 5% CO2. CHO-K1, Chinese hamster ovary cell line, was cultured in F-12 supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C and 5% CO2.
As for transient transfection, cells were transfected with expression plasmid by Lipofectamine 2000 (Thermo Fisher Scientific). CHO-K1 stably expressing gpCXCR1 or gpCXCR2 was established by standard method. In brief, CHO-K1 was transfected with pcDNA3.1/gpCXCR1 or gpCXCR2 using Lipofectamine 2000. The transfected CHO-K1 was incubated in F-12 supplemented with 10% FBS, 100 U/mL penicillin, 100 μg/mL streptomycin, and 200 μg/mL G418, and surviving cells were cloned.
Flow cytometry analysis
The binding activity of mouse serum, hybridoma supernatant, and the anti-gpCXCR1 and gpCXCR2 antibodies was assessed by flow cytometric analysis. Cells expressing gpCXCR1 or gpCXCR2 were detached using TrypLE Express (Thermo Fisher Scientific) and resuspended in staining buffer, PBS supplemented with 5% FBS. The samples were diluted with staining buffer to indicated concentration. Cells were stained with samples for 30 minutes at 4°C and washed with staining buffer in each staining step; 10 μg/mL of AlexaFluor488-conjugated goat anti-mouse IgG was used as a secondary antibody. As a control antibody, mouse IgG (Thermo Fisher Scientific) was used. LIVE/DEAD Fixable Far Red Dead Cell Stain Kit (Thermo Fisher Scientific) was used for exclusion of dead cells. Fluorescence was measured by FC500 (Beckman Coulter, Brea, CA).
Migration assay
CHO-K1, CHO/gpCXCR1, and CHO/gpCXCR2 were detached using TrypLE Express and suspended in assay buffer, DMEM/F-12 without phenol red supplemented with 0.1% BSA, at concentration of 105 cells/mL and indicated concentration of antibodies. GpIL-8 and gpGROα were diluted to the indicated concentrations in figures with the assay buffer. Before the assay, FluoroBlok™ HTS 24-well multiwell permeable support system with 8.0 μm high-density polyethylene terephthalate (PET) membrane (Corning, Corning, NY) was coated with 10 μg/mL of fibronectin. The coated chamber was set onto the 24-well plate, and 250 μL of cell suspension with or without antibody was added to the chamber, followed by addition of 750 μL of chemoattractant to the lower well of the 24-well plate. The chamber was incubated for 4 hours at 37°C and 5% CO2. After the incubation, the chamber was stained with 4 μg/mL calcein AM for 15 minutes at 37°C, and multipoint fluorescence of the underside of the chamber was measured by using SpectraMax M3 (Molecular Devices, Sunnyvale, CA) at 480 nm excitation and 530 nm fluorescence.
Results
Mouse immunization and establishment of hybridoma producing anti-gpCXCR1 or anti-gpCXCR2 antibody
To produce anti-gpCXCR1 or anti-gpCXCR2 antibodies, we conducted DNA immunization of BALB/c mice. After immunization, 1/200 diluted serum of gpCXCR1-immunized and gpCXCR2-immunized mice exhibited binding to HEK293T transiently expressing gpCXCR1 (293T/gpCXCR1) and gpCXCR2 (293T/gpCXCR2), respectively (Fig. 1A, B). Both serum samples showed specific binding to each antigen. Hybridomas were established using lymph nodes from gpCXCR1-immunized and gpCXCR2-immunized mice, and binding of supernatants was evaluated against 293T/gpCXCR1 or 293T/gpCXCR2 with flow cytometric analysis. As a result of the supernatant screening, hybridoma CR1-002, which produced antibody that bound to 293T/gpCXCR1, and hybridoma CR2-004, which produced antibody that bound to 293T/gpCXCR2, were obtained (Fig. 1C, D).
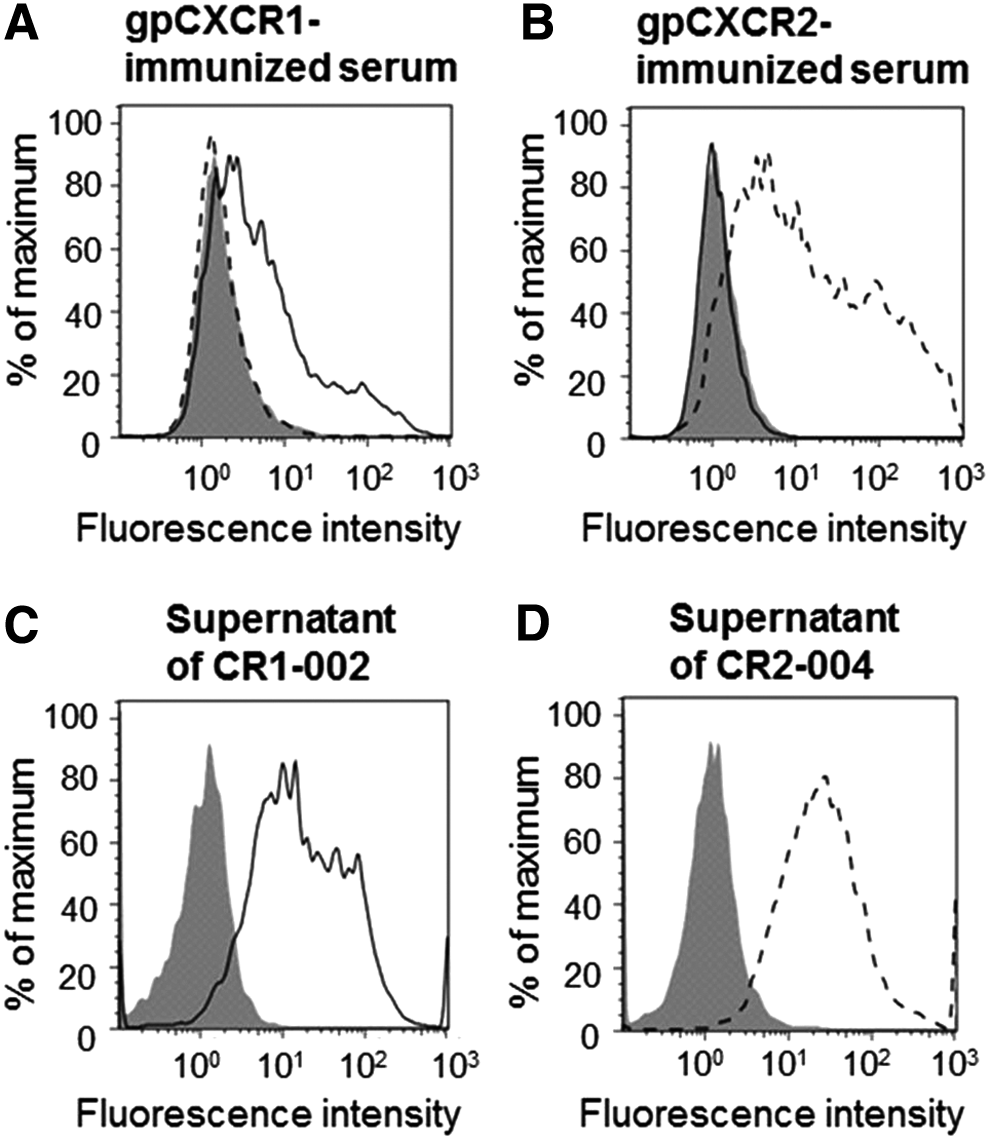
Binding of serum samples from immunized mice and hybridoma supernatants to cells transiently expressing gpCXCR1 or gpCXCR2. Binding of 1/200 diluted serum from
Binding specificity of anti-gpCXCR1 or anti-gpCXCR2 antibody against gpCXCR1 and gpCXCR2 stably expressing cells
Monoclonal antibodies, AbCR1 and AbCR2, were purified from supernatants of the hybridoma CR1-002 and CR2-004. Purity of monomeric IgG of AbCR1 and AbCR2 was 97.7% and 97.8%, respectively. Endotoxin levels were both <0.5 EU/mL, in which the in vitro effect of endotoxin could not be observed.(22) The isotype of AbCR1 and AbCR2 was IgG2a and IgG2b, respectively. For assessment of antibodies, we established CHO-K1 cells stably expressing either gpCXCR1 (CHO/gpCXCR1) or gpCXCR2 (CHO/gpCXCR2). Binding activities of AbCR1 and AbCR2 to CHO-K1, CHO/gpCXCR1, and CHO/gpCXCR2 were evaluated using flow cytometry. Purified AbCR1 and AbCR2 bound only to specific antigens, gpCXCR1 and gpCXCR2, respectively (Fig. 2). Therefore, antibodies that can distinguish gpCXCR1 and gpCXCR2 were successfully obtained. The binding histogram suggests that CHO/gpCXCR1 expressed gpCXCR1 heterogeneously and CHO/gpCXCR2 expressed gpCXCR2 homogeneously (Fig. 2C, D).
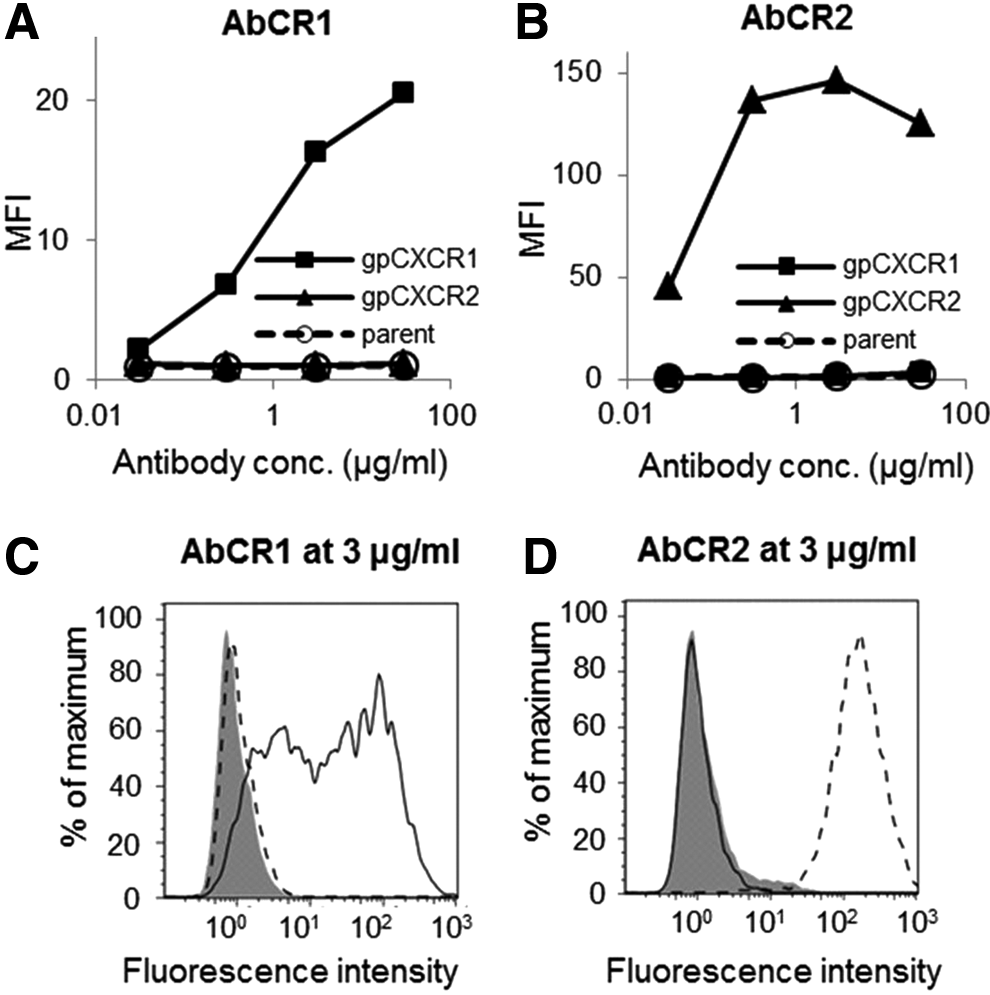
Binding of anti-gpCXCR1 and anti-gpCXCR2 antibodies to cells stably expressing gpCXCR1 or gpCXCR2. Binding of
Migration of CHO/gpCXCR1 or gpCXCR2 to gpIL-8 and gpGROα
To evaluate the activity of antibodies, we investigated whether CHO/gpCXCR1 and CHO/gpCXCR2 migrate toward gpIL-8 and gpGROα. Recombinant gpIL-8 and gpGROα were produced. The molecular sizes of gpIL-8 and gpGROα were estimated to be 11 and 10 kDa, respectively, in SDS-PAGE under reducing conditions and they were matched with the calculated molecular weight (data not shown). No band except the objective protein was observed in the lanes loaded with the IL-8 and GROα samples. Endotoxin levels were both <0.5 EU/mL. In the migration system of the Boyden chamber coated with fibronectin, gpIL-8 induced migration of CHO/gpCXCR1 in a concentration-dependent manner from 4 nM, but gpGROα did not induce migration of CHO/gpCXCR1 (Fig. 3A, B) at 100 nM. On the other hand, both gpIL-8 and gpGROα induced migration of CHO/gpCXCR2 in a concentration-dependent manner from 4 nM (Fig. 3C, D). The receptor selectivity of gpGROα was found to be consistent with the human GROα. The parent CHO-K1 did not migrate in response to gpIL-8 or gpGROα (data not shown). These results indicate that the inhibitory activity of anti-gpCXCR1 or gpCXCR2 antibodies can be evaluated by a migration system of gpIL-8, which induced migration of both CHO/gpCXCR1 and CHO/gpCXCR2, or a migration system of gpGROα, which induced migration of only CHO/gpCXCR2, due to their consistent relationship to humans among receptors and ligands.
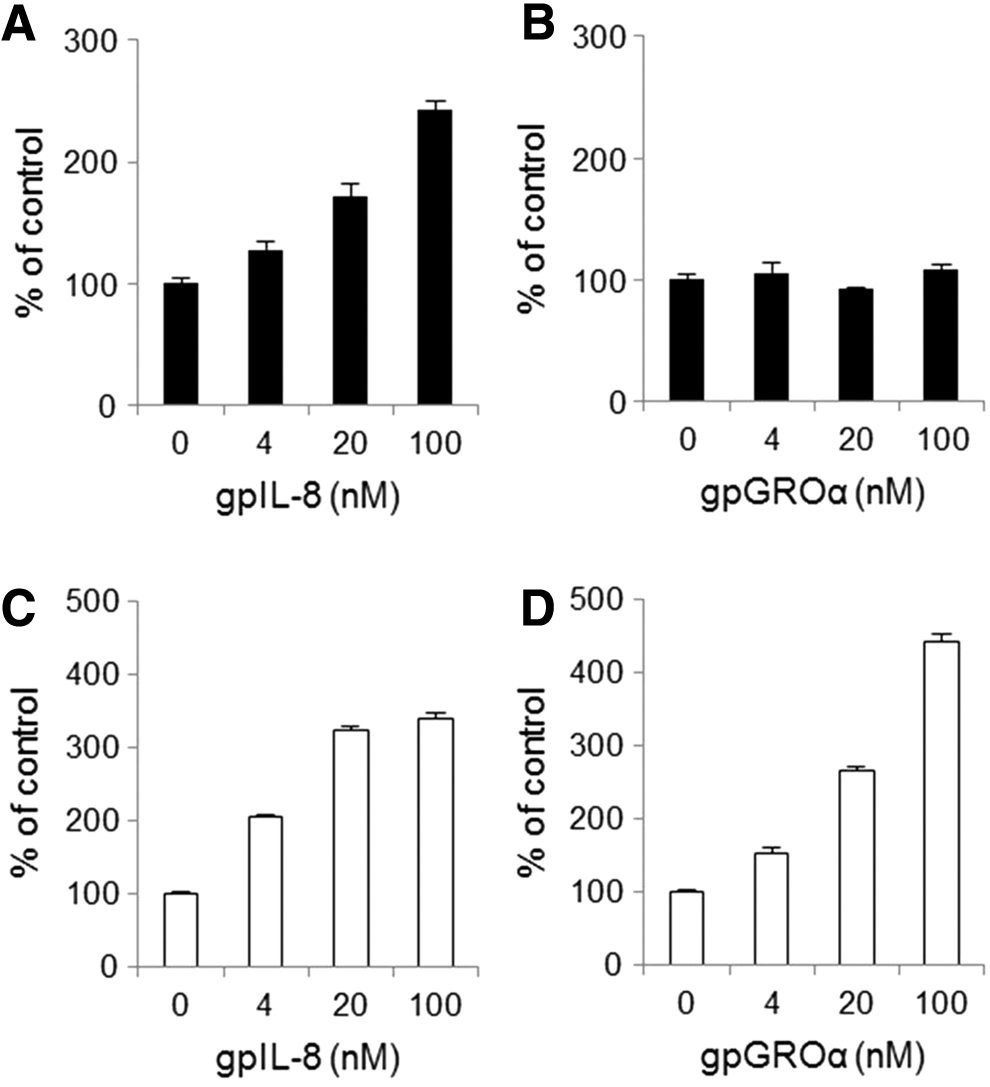
Migration of cells stably expressing either gpCXCR1 or gpCXCR2 induced by chemokine ligand. Migration of CHO/gpCXCR1 (closed bar) induced by
Inhibitory activity of anti-gpCXCR1 or gpCXCR2 antibody in the migration assay
Inhibitory activity of AbCR1 and AbCR2 was evaluated in the migration assay using CHO/gpCXCR1 and CHO/gpCXCR2. Chemotactic responses of the cells were induced by gpIL-8 or gpGROα at a concentration that we observed for sufficient migration to evaluate the inhibitory activity of antibodies. AbCR1 inhibited CHO/gpCXCR1 migration induced by 10 nM of gpIL-8 in a concentration-dependent manner (Fig. 4A). AbCR2 inhibited CHO/gpCXCR2 migration induced by 10 nM of gpIL-8 and also inhibited the cell migration induced by 30 nM of gpGROα in a concentration-dependent manner (Fig. 4B, C). The control mouse IgG did not show inhibitory effects on the migration of CHO/gpCXCR1 or CHO/gpCXCR2. These results indicate that AbCR1 and AbCR2 possess the inhibitory activity to each specific antigen, gpCXCR1 and gpCXCR2, respectively.
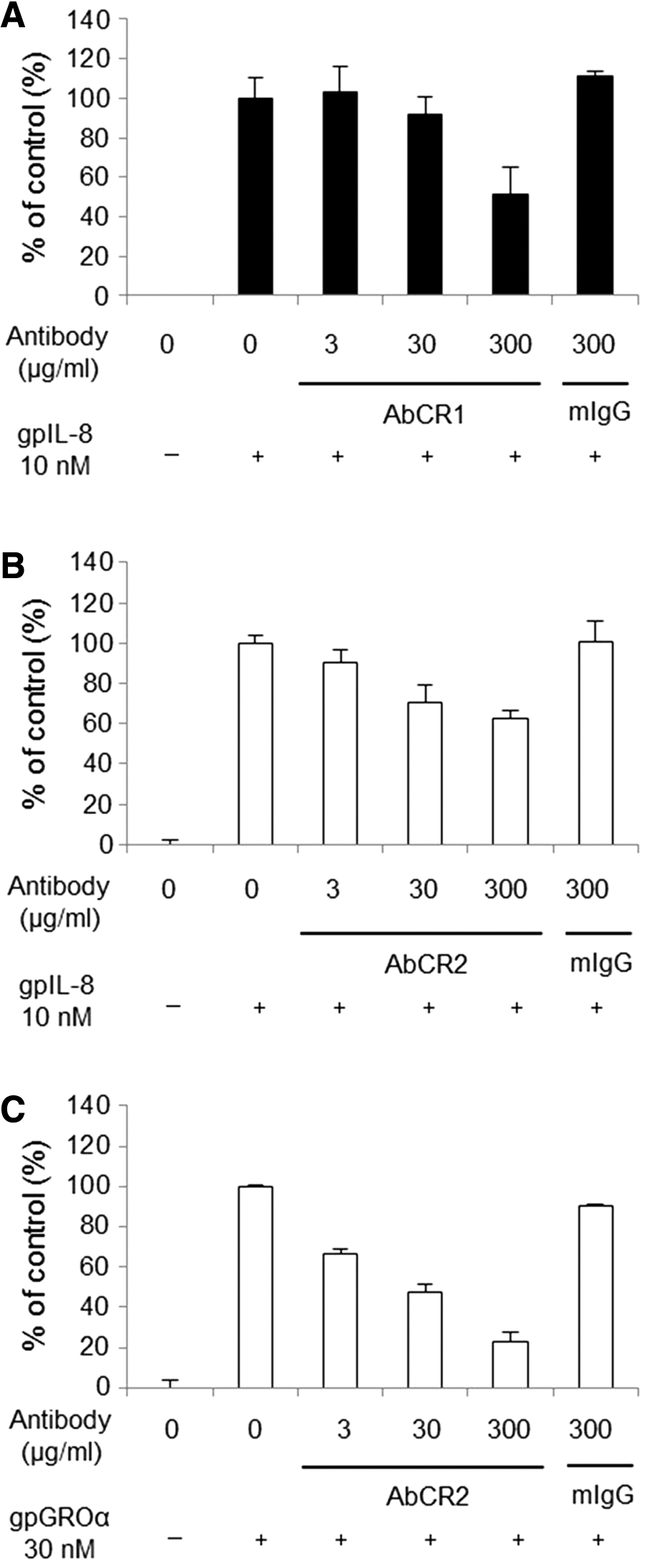
Inhibitory activity of anti-gpCXCR1 and gpCXCR2 antibodies against migration of cells expressing either gpCXCR1 or gpCXCR2.
Discussion
This study aimed to acquire inhibitory monoclonal antibodies specific to gpCXCR1 and gpCXCR2. We conducted DNA immunization of mice, screened, and selected AbCR1 and AbCR2, monoclonal antibodies specific to gpCXCR1 and gpCXCR2, respectively. We further demonstrated the inhibitory activity of AbCR1 and AbCR2 by establishing the cell migration assay in which gpIL-8 induced chemotactic responses of CHO/gpCXCR1 and CHO/gpCXCR2 or gpGROα induced chemotactic responses of CHO/gpCXCR2. As far as we know, this is the first report to show anti-gpCXCR1 and gpCXCR2 monoclonal antibodies with inhibitory activity.
There has been no report on the acquisition of anti-gpCXCR1 and anti-gpCXCR2 antibodies so far other than our present report. As for anti-hCXCR1 or hCXCR2 antibodies, some reports show an inhibitory effect to hCXCR1 or hCXCR2.(16–19) To investigate whether anti-hCXCR1 or hCXCR2 antibodies would bind to gpCXCR1 or gpCXCR2, we collected four anti-hCXCR1 antibodies and five anti-hCXCR2 antibodies, which were available from commercial venders, and assessed their binding to CHO/gpCXCR1 or gpCXCR2, but none of the antibodies showed binding. Conversely, our anti-gpCXCR1 and gpCXCR2 antibodies did not show binding to hCXCR1 or hCXCR2. Sequence identity of the extracellular domain of CXCR1 and CXCR2 between humans and guinea pigs is 53% and 53%, respectively, and this probably leads to binding discrepancy between humans and guinea pigs.
Our study of cell migration assay using recombinant gpIL-8 and gpGROα revealed a consistency between humans and guinea pigs in terms of chemokine ligand specificity of CXCR1 and CXCR2. In our study, gpIL-8 induced migration of both CHO/gpCXCR1 and CHO/gpCXCR2. In contrast, gpGROα induced migration of only CHO/gpCXCR2. These results are consistent with human CXCR1 and CXCR2. Takahashi et al. previously reported that gpIL-8 induced migration of HEK293 cells stably expressing gpCXCR1 and also HEK293 cells expressing gpCXCR2, but they could not show migration concerning gpGROα because of its unavailability.(12) Thus, our result is the first to demonstrate functional validity of gpCXCR2 in migratory response toward gpGROα. These results indicate that AbCR1 and AbCR2 are authentic inhibitory antibodies against gpCXCR1 and gpCXCR2.
The inhibitory effects of AbCR1 on chemotactic responses of CHO/gpCXCR1 toward gpIL-8 and that of AbCR2 on chemotactic responses of CHO/gpCXCR2 toward gpIL-8 were both partial. One possible reason is that antibody concentration was not sufficient to inhibit the migration completely. We could not increase the concentration of the antibodies due to the limitation of concentration of stock antibody. Another plausible reason is the epitope difference between antibodies and ligands. Modified gpCXCR1 and gpCXCR2 whose extracellular loops are substituted with the corresponding extracellular loops of human ones will be useful for epitope identification according to the previous report, which attempted to define the epitope of chemokine to its receptor.(23) This examination of the epitope remains a subject for further study concerning anti-gpCXCR1 and gpCXCR2 antibodies.
The specific antibodies that can distinguish gpCXCR1 and gpCXCR2 are helpful to elucidate the function of CXCR1 and CXCR2, especially in neutrophilic respiratory diseases because guinea pigs are often utilized for respiratory disease models.(14) The expression pattern of CXCR1 and CXCR2 can be detected by these antibodies. The physiological roles of CXCR1 and CXCR2 can also be elucidated. Until now, a variety of small compounds inhibiting CXCR1/CXCR2 have been studied, but no group has clarified the CXCR1 contribution to the neutrophilic disease because of the absence of IL-8 in typical animal models using mice and rats.(8–10) Recently, Planagumà et al. evaluated the inhibitory activity of these selective antagonists in guinea pigs to elucidate CXCR1 function. They found that the compounds suppressed both gpCXCR1 and gpCXCR2 function at similar efficacy due to the interspecies difference, but they failed to elucidate the contribution of gpCXCR1.(15) Our anti-gpCXCR1 and gpCXCR2 antibodies will overcome this hurdle because they show clear specificity to each receptor.
In conclusion, we successfully obtained specific antibodies against gpCXCR1 and gpCXCR2 with inhibitory activity. These antibodies will be valuable for elucidation of physiological roles of CXCR1 and CXCR2 in guinea pigs.
Footnotes
Acknowledgments
The authors would like to thank Ms. Ikue Iino for her technical contribution. They would also like to thank Dr. Toshinori Agatsuma and Dr. Futoshi Nara for their critical review of the manuscript. This research did not receive any specific grant from funding agencies in public, commercial, or not-for-profit sectors.
Author Disclosure Statement
No competing financial interests exist.