
Abstract
DNAM-1 (CD226) is an activating immunoreceptor expressed on lymphocytes and myeloid cells. CD155 and CD112 are the ligands for DNAM-1. DNAM-1 plays an important role in tumor immunity mediated by CD8+ T cells and NK cells. Moreover, the interaction of DNAM-1 with the ligands contributed to the development of acute graft versus host disease (GVHD) and treatment with anti-DNAM-1 monoclonal antibodies (mAb) dramatically improved acute GVHD in a mouse model, suggesting that DNAM-1 may be a good molecular target for therapy to acute GVHD in human. In this study, we generated and characterized five novel clones of anti-human DNAM-1 mAbs, named TX94, TX95, TX96, TX107, and TX108. Among these mAbs, TX94 is a unique neutralizing mAb that most efficiently blocked the interaction between DNAM-1 and CD155. Furthermore, TX94 inhibited NK cell-mediated cytotoxicity against a tumor cell line and suppressed CD8+ T cell proliferation mediated by allogeneic mixed lymphocyte reaction. Thus, TX94 may be useful for molecular therapy targeting DNAM-1.
Introduction
T
We also showed that an anti-mouse DNAM-1 mAb (TX42) inhibited the cytotoxicity against CD155-expressing tumor cells by CD8+ T cells and NK cells.(2,5) Furthermore, we demonstrated the critical role of DNAM-1 in the pathogenesis of acute graft versus host disease (GVHD) in a mouse model. The administration of TX42 to mice inhibited the development of GVHD.(10) Thus, DNAM-1 might be a molecular target for therapeutic approaches to GVHD in humans.
In this study, we have generated and characterized five clones of anti-human DNAM-1 mAbs, named TX94, TX95, TX96, TX107, and TX108. We show that TX94 is a unique mAb that is potentially useful for the molecular therapy targeting DNAM-1.
Materials and Methods
Mice
BALB/c nu/nu mice were purchased from CLEA (Tokyo, Japan) and housed under specific pathogen-free conditions at the Animal Resource Center of the University of Tsukuba. The animal protocol was approved by the Institutional Review Committee and performed according to the guidelines of the University of Tsukuba.
Antibodies
Five novel mouse anti-human DNAM-1 mAbs, TX94 (IgG1), TX95 (IgG2a), TX96 (IgG1), TX107 (IgG1), and TX108 (IgG2b) were generated in our laboratory. Briefly, lymphocytes were purified from BALB/c mice that were injected with BW5147 transfectant expressing human DNAM-1 (DNAM-1/BW5147) into the foot pads for immunization on day 0 and boosted with chimeric fusion protein consisting of the extracellular portion of human DNAM-1 and the Fc portion of human IgG (DNAM-1-Fc fusion protein) on day 7. The draining lymph node cells were then fused with the myeloma cells SP2/O, as described.(11) We obtained 31 clones specific to DNAM-1 and focused on the five clones that strongly bound to DNAM-1 for characterization. TX25 (IgG1) was previously generated in our laboratory. All of these antibodies were biotinylated by using sulfo-NHS-biotin (Funakoshi, Tokyo, Japan) and 0.1 M sodium bicarbonate (Sigma-Aldrich, St. Louis, MO). TX25 was conjugated by the Alexa Fluor® 647 Labeling Kit (Life Technologies, Carlsbad, CA). PE-conjugated antibodies against human CD4, CD8, CD19, CD56, CD14, FITC-conjugated antibody against CD3, V450-conjugated antibody against human CD56, and FITC-conjugated streptavidin and Golgi-Stop™ were purchased from BD Biosciences (San Jose, CA). PE-conjugated anti-human IgG (Fc) antibody was purchased from Beckman Coulter (Brea, CA). APC-conjugated antibody against human CD107a was purchased from BioLegend (San Diego, CA).
Generation of human DNAM-1-expressing transfectant and chimeric proteins of DNAM-1 and CD155 fused with IgG Fc portion
DNAM-1/BW5147 transfectant was established, as described.(12) Chimeric fusion proteins consisting of the extracellular portion of human DNAM-1 or CD155 and the Fc portion of human IgG (DNAM-1 or CD155-Fc fusion protein, respectively) were generated with chimeric cDNA of the entire extracellular domain of human DNAM-1 or CD155 with human IgG Fc portion as described.(4)
Purification of human peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMC) were isolated from healthy volunteers by Lymphoprep™ (STEMCELL Technologies, Vancouver, BC) density centrifugation after obtaining their informed consent.
Blocking assay
DNAM-1/BW5147 transfectants were pretreated with a saturation dose of CD155-Fc fusion protein, and then stained with each biotinylated mAb. CD155-Fc fusion protein and biotinylated mAbs were detected by PE-conjugated anti-human IgG (Fc) antibodies and APC-conjugated streptavidin, respectively.
Competitive binding assay
DNAM-1/BW5147 transfectants were pretreated with the saturation dose of Alexa Flour 647-conjugated TX25, and then stained with biotinylated TX94 or TX95, followed by FITC-conjugated streptavidin. Conversely, DNAM-1/BW5147 transfectants were pretreated with the saturation dose of biotinylated TX94 or TX95, and then stained with Alexa Flour 647-conjugated TX25.
Cytotoxicity assay
CD56+ cells were isolated from human PBMC by magnetic-activated cell sorting (MACS) (Miltenyi Biotec, Bergisch Gladbach, Germany) and cocultured with Colo205 in the presence of 1 μg of F(ab′)2 fragments of TX94 or control mouse IgG1 and 1 μg of APC-conjugated anti-CD107a antibody and 0.1 μL of Golgi-Stop (NK: Colo205 = 1: 1) for 6 hours. The expression level of CD107a in NK cells (CD3−CD56+) was analyzed by flow cytometry.
Mixed lymphocytes reaction
CD14+ cells were isolated from PBMC by MACS and cultured in the presence of granulocyte macrophage colony-stimulating factor (GM-CSF) (25 ng/mL) and IL-4 (20 ng/mL) for 7 days to induce differentiation into dendritic cells (DCs). Induced DCs were treated with Mitomycin C (50 μg) for 30 minutes. CD8+ T cells were isolated from PBMC of an allogeneic volunteer by MACS and cocultured with the induced DCs (CD8+ T: DC = 10: 1) in the presence of 1 μg/mL F(ab′)2 fragment of TX94 or control mouse IgG1. CD8+ T cell proliferation was analyzed by using Cell Proliferation ELISA, BrdU (Roche Life Science, Mannheim, Germany).
Results and Discussion
We generated 31 new clones of anti-DNAM-1 mAbs and focused on characterization of five mAbs, TX94, TX95, TX96, TX107, and TX108, which strongly bound to DNAM-1. Each mAb showed unique amino acid sequences that were different from the others in complementarity-determining regions, as determined by genomic analyses. These mAbs as well as the TX25 mAb specifically stained DNAM-1/BW5147 transfectant, but not parental BW5147 cells (Fig. 1A), indicating that these mAbs are specific to human DNAM-1. Since TX94 and TX25 most strongly bound to DNAM-1/BW5147 transfectant, we examined the reactivity of TX94 and TX25 to PBMC. Both clones showed comparable staining to CD4+ T cells and CD8+ T cells, B cells, NK cells, NKT cells, monocytes (Fig. 1B).
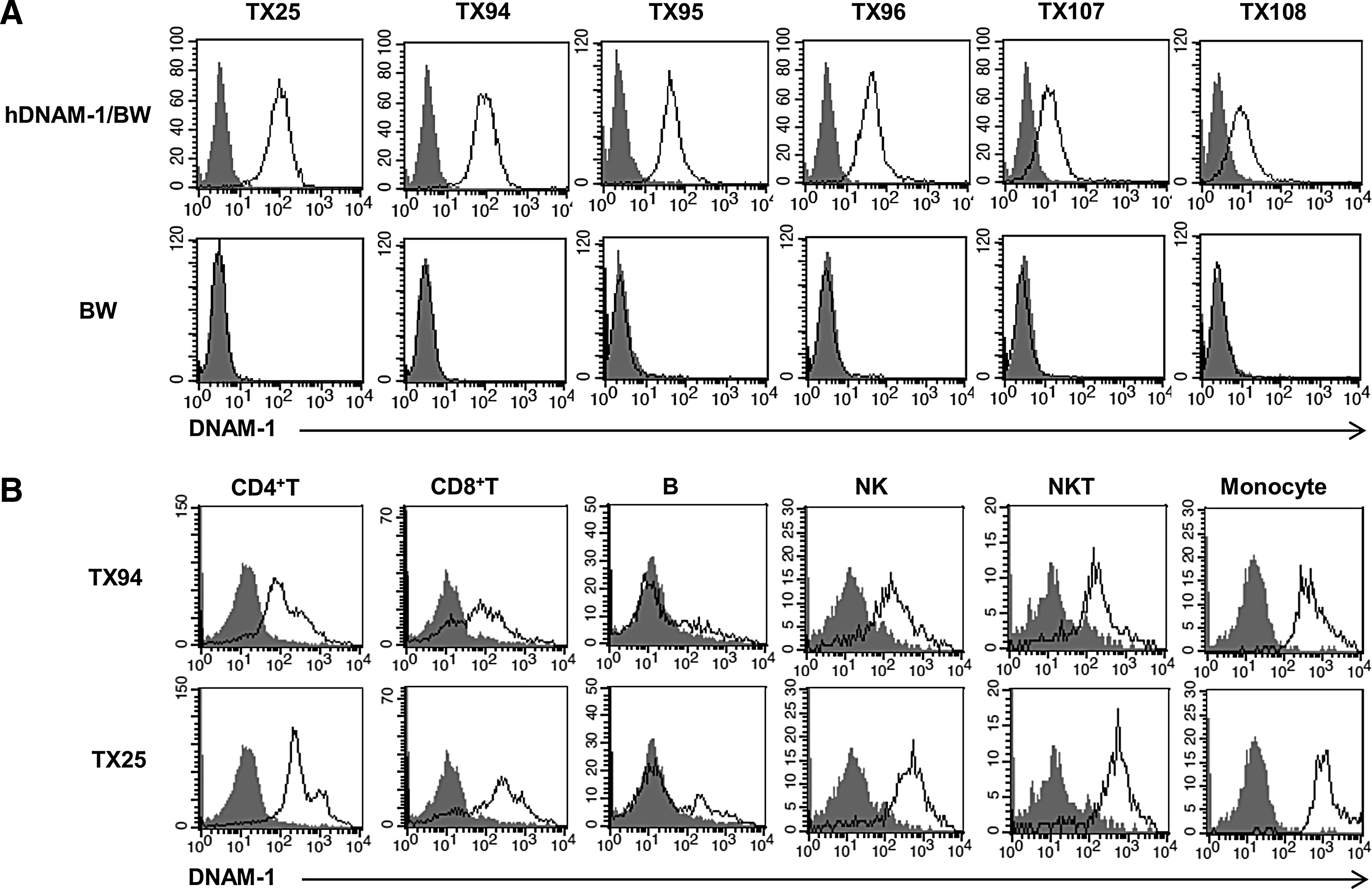
Specificity of mAbs to human DNAM-1.
To examine whether these mAbs interfere with the interaction of DNAM-1 and CD155, DNAM-1/BW5147 were pretreated with the CD155-Fc fusion protein at a saturated dose, and then treated with each biotinylated anti-human DNAM-1 mAb. Binding of anti-human DNAM-1 mAbs and human CD155-Fc fusion protein were detected by APC-conjugated streptavidin and PE-conjugated anti-human IgG, respectively. TX94 showed the most efficient binding to DNAM-1/BW5147 transfectant among these mAbs (Fig. 2A). TX94 also showed the strongest competitive ability among these mAbs for binding of DNAM-1/BW5147 to CD155-Fc fusion protein (Fig. 2B).
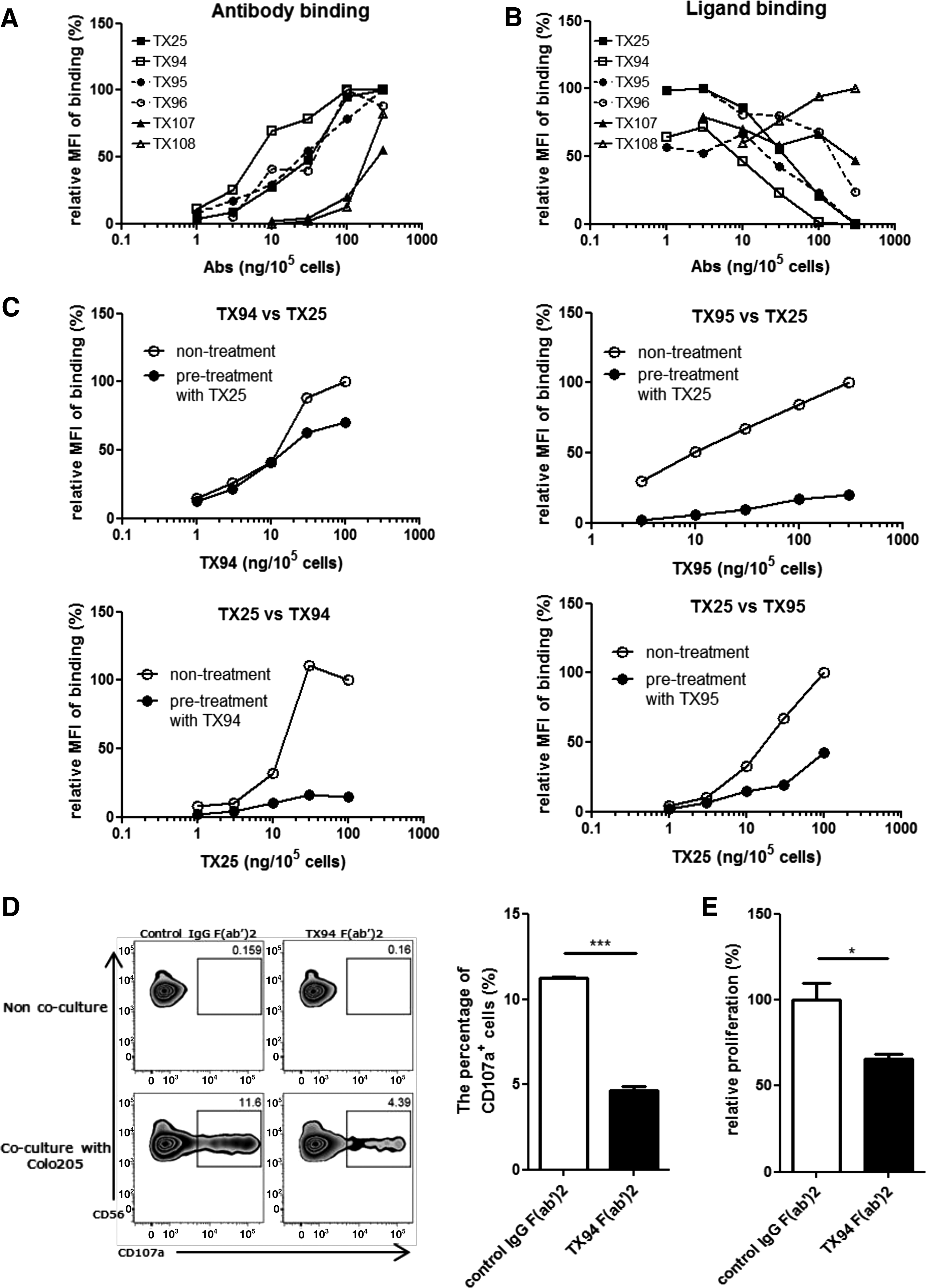
Blocking of the interaction of DNAM-1 and CD155.
To compare the affinity with DNAM-1 of newly established mAbs and TX25, we next performed competitive assay for binding of TX94, TX95, and TX25 to DNAM-1. DNAM-1/BW5147 transfectant was pretreated or not with a saturated dose of TX25 and then treated with biotinylated TX94 or TX95, followed by FITC-conjugated streptavidin. Whereas TX25 significantly inhibited the binding of TX95 to DNAM-1/BW5147 transfectant, the effect of TX25 pretreatment on the binding of TX94 was scarcely observed (Fig. 2C). Conversely, pretreatment with a saturated dose of TX94 and TX95 significantly and moderately inhibited the binding of TX25 to DNAM-1/BW5147 transfectant, respectively (Fig. 2C). These results suggest that the affinity of TX94 for DNAM-1 is higher than those of TX25 and TX95.
To examine whether TX94 inhibits the cytotoxicity of NK cells against tumor cells, we cocultured NK cells with Colo205, human colon cancer cell line, in the presence of F(ab′)2 fragments of TX94 or control IgG for 6 hours. TX94 significantly inhibited CD107a expression on NK cells (Fig. 2D). These results indicated that TX94 blocked the interaction of DNAM-1 on NK cells with CD155 on tumor cells, resulting in the inhibition of cytotoxicity.
We previously reported that the interaction between DNAM-1 and CD155 was involved in alloreactive T cell activation in mice.(10) To examine whether TX94 inhibited alloreactive T cell proliferation in humans, we performed an allogeneic mixed lymphocytes reaction (MLR). Human CD8+ T cells were cocultured with DCs derived from an allogeneic donor in the presence of F(ab′)2 fragments of TX94 or control IgG for 5 days. TX94 significantly inhibited CD8+ T cell proliferation compared with control IgG (Fig. 2E), indicating that TX94 inhibits alloreactive T cell proliferation.
In conclusion, TX94 is a high-affinity mAb against human DNAM-1, which efficiently blocked the ligand binding to DNAM-1 and inhibited the alloreactive proliferation of human CD8+ T cells. These results suggest that TX94 may be useful for molecular therapy targeting DNAM-1.
Footnotes
Acknowledgments
The authors thank S. Tochihara and Y. Nomura for their secretarial assistances and T. Nabekura for discussion. This study was supported in part by grants provided by the Ministry of Education, Culture, Sports, Science, and Technology of Japan (Grant number 16H06387 to A.S. and Grant number 16H05169 to K.S.).
Author Disclosure Statement
No competing financial interests exist.