
Abstract
Incubation of B10.MLM cells, a cell line of alveolar macrophages, with lycopene, a carotenoid, leads to an increase of lycopene content in their microsomal fraction. That increase was higher and developed faster when the cells were incubated with immune complexes formed by lycopene and mAb 6B9 (L–6B9 mAb), a monoclonal hapten-specific antibody raised against lycopene, as compared with dimethyl sulfoxide (DMSO)-dissolved lycopene (DMSO-L). Moreover, incubation of B10.MLM cells with L–6B9 mAb complexes was accompanied by more efficient accumulation of lipid droplets in the cultured cells and more significant inhibition of mRNA for 3-hydroxy-3-methylglutaryl-coenzyme (HMG-CoA) reductase, a rate-limiting enzyme of cholesterol biosynthesis known to be targeted by lycopene. Additionally, there was a better inhibition of Chlamydia trachomatis infection in B10.MLM cells infected with the pathogen and incubated thereafter with L–6B9 mAb complexes as compared with DMSO-L.
Altogether, the results suggest that association with monoclonal antibody promotes intracellular delivery of lycopene in cultured cells possibly through Fc-receptor mediated uptake.
Introduction
C
Lycopene (C40H56), a 40-carbon highly nonpolar acyclic mevalonate-derived carotenoid can be identified and isolated from tomatoes, watermelons, guavas, and papayas in the form of all-trans isomers.(5,7) Eleven conjugated double bonds in the lycopene structure govern its extraordinary antioxidant capacity, surpassing the antiradical properties of all known nonpolar carotenoids multifold.(5,8) The antioxidant properties of lycopene along with its effects on immunological parameters, inflammatory cytokine production, nuclear receptors, cell cycle progression and growth factor signaling explain the multiple favorable health effects of lycopene.(9) In particular, from the epidemiological point of view, lycopene consumption has been shown to be inversely associated with frequency of prostate cancer and the severity and outcomes of cardiovascular disease (CVD) and diabetes.(10–13) Interventional studies on the health effects of lycopene are less consistent and often encounter questionable reproducibility.(11,14) Blood lycopene and carotenoid concentrations have a better prognostic value for health outcomes than their dietary intake values.(5,15,16) This emphasizes the importance of carotenoid bioavailability status for lycopene biopharmacology. As we have shown recently,(5) a microencapsulation approach, in particular the use of lycosome technology, can efficiently overcome low intestinal bioavailability of lycopene and promote its effect on cardiovascular parameters and oxidative status in CVD patients.
In the present article, we address another important mechanism controlling lycopene bioavailability. As we report here, association with monoclonal antibody promotes intracellular delivery of lycopene in cultured cells.
Materials and Methods
Reagents and organisms
All reagents were from Sigma-Aldrich unless specified otherwise. Chlamydia trachomatis strain L2/Bu434 was kindly provided by Dr. P. Saikku (University of Oulu, Finland). Lycopene oleoresin all-trans isomers was purchased from LycoRed (LycoRed, Switzerland) and kept in oxygen-free containers at −80°C. Stock solutions dissolved in dimethyl sulfoxide (DMSO) at concentrations of 50 mM were made before each experiment.
Lycopene–mAb complex
mAb 6B9 was isolated from hybridoma ascites(17) and purified using an ammonium sulfate precipitation protocol(18) and the LigaTrap™ IgM Purification Kit (Ligatrap, LA). Briefly, between 1 and 10 mg/mL of the purified mAb in Dulbecco's phosphate-buffered saline (DPBS) was mixed with 8 molar equivalents of all-trans lycopene dissolved in DMSO under conditions of continuous microstirring and temperature shift from 25°C to 30°C. The mixture was allowed to react for 30 minutes at 30°C. Formation of the 6B9 mAb-lycopene complex was confirmed and the association constant determined by absorption spectroscopy at 472 nm.(19,20)
The resulting 6B9 mAb-lycopene complex was further purified using Amicon Ultra 30,000 molecular weight cutoff centrifugal filters (Millipore) and DPBS buffer exchange (3 × ) then used for cell culture studies within 1 hour of preparation. Lycopene concentration and protein content in the purified complex were measured with a spectrophotometer at 472 and 260 nm, respectively.
Cultured cells
C. trachomatis infection in B10.MLM cells
C. trachomatis was initially propagated in McCoy cells and elementary bodies (EB) were purified by Renografin gradient centrifugation as previously described.(21) Purified EB of known titer were suspended in sucrose–phosphate–glutamic acid buffer and used as inoculums for B10.MLM cells. Cells were grown in 24-well plates until a confluence rate of 80% was reached. B10.MLM plates were infected with C. trachomatis at multiplicity of infection of 30 in DMEM with 5% fetal bovine serum without cycloheximide and centrifuged for 0.5 hour at 1500 g. After 1 hour of incubation at 37°C, the cell monolayers were washed with DMEM. DMSO-dissolved lycopene, mAb 6B9, and lycopene-mAb complex were added as described above. IF staining was performed after overnight incubation.
Infected B10.MLM monolayers grown on coverslips in 24-well plates and subjected to overnight lycopene/mAb 6B9 additions were fixed with methanol. Permeabilized cells were stained for direct IF using FITC-conjugated monoclonal antibody against chlamydial lipopolysaccharide (Nearmedic Plus, RF). Inclusion-containing cells were visualized using a Nikon Eclipse 50i fluorescence microscope at ×200 and ×1000 magnification.
Lipid droplet staining
For neutral lipid staining, B10.MLM cells grown on coverslips were incubated with lycopene for 18 hours. Then cells were washed with PBS twice, fixed with 3% formaldehyde/0.025% glutaraldehyde at room temperature for 20 minutes, and stained with BODIPY 493/503 (Molecular Probes; Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. Cells were visualized using a Nikon Eclipse 50i fluorescence microscope at ×1000.
Microsome isolation from B10.MLM cells
The total microsomal fraction from the B10.MLM cells was prepared as published elsewhere.(22) Briefly, the cell monolayers were gently washed with ice-cold PBS containing proteinase inhibitors, scraped from the dish surface, and suspended in 1 mL of sucrose buffer (0.25 M sucrose, 1 mM EDTA, 10 mM HEPES-NaOH, pH 7.4). All steps were carried out at 4°C. After 20 cycles of homogenization with a syringe, the cell homogenate was centrifuged at 1000 g for 10 minutes to pellet the nuclear fraction. The supernatant was transferred to ultracentrifuge tubes and subjected to ultracentrifugation at 100,000 g for 1 hour. The resulting pellet was used for quantification of lycopene content.
Quantification of lycopene in microsomal fraction of B10.MLM cells
Quantification of lycopene was performed by spectrophotometric assay.(19) Briefly, B10.MLM cells incubated overnight with lycopene–6B9 mAb complex and appropriate controls were harvested. Total microsome fraction was isolated and mixed with 1.0 mL of extraction mixture (50:50 hexane-ethyl acetate and 1% butylated hydroxytoluene). After two rounds of homogenization with a glass syringe and extensive vortexing, the mixture was centrifuged at 1000 g for 10 minutes and the supernatant was assessed at 472 nm using a spectrophotometer. OD472 values were referenced to the protein content of the specimens measured using the Lowry Protein Assay Kit (Thermo Fisher, Wilmington).
RNA analysis
Total RNA extraction and cDNA preparation from B10.MLM cells cultured overnight with the lycopene–6B9 mAb complex were performed as described previously.(22) The mRNA levels for 3-hydroxy-3-methyglutaryl CoA reductase (HMG CoA reductase, human-specific primers: sense-5′-CAAGGAGCATGCAAAGATAATCC-3′ antisense -5′-GCCATTACGGTCC CACACA-3′) were measured by quantitative RT-PCR using an ANK 32 thermocycler (Syntol, RF) and referenced to the mRNA values of human beta actin (β-actin, primers: sense—5′-GCACCCAGCACAATGAAGAT-3′, antisense - 5′-GCCGATCCACACGGAGTAC-3′). The mRNA expression levels in the B10.MLM cells cultured overnight with the lycopene–6B9 mAb complex were referenced to the CT values in B10.MLM cells grown under the same conditions in the presence of DMSO alone. This reference value was taken as 1.00. Each cDNA sample was tested by PCR at least three times. All experiments were repeated at least twice. Representative sets of results obtained in the cell culture experiments with B10.MLM cells are shown below.
Results
Lycopene content in microsomal fraction of B10.MLM cells
First, we decided to investigate the changes in intracellular, microsome-associated lycopene content of B10.MLM cells after overnight incubation with lycopene–6B9 mAb complex. As can be seen from Figure 1A, overnight incubation with lycopene–6B9 mAb complex leads to a statistically significant increase (up to 116.20 ± 6.73 ng/μg of protein) of lycopene content in microsomal fraction of B10.MLM cells as compared with DMSO-dissolved lycopene (59.62 ± 5.55 ng/μg of protein). There was no detectable lycopene in microsomal fraction of B10.MLM cells incubated overnight with DMSO alone or mAb 6B9 alone, which were used as the two major control groups in the experiment (results not plotted, readings below assay sensitivity limit).
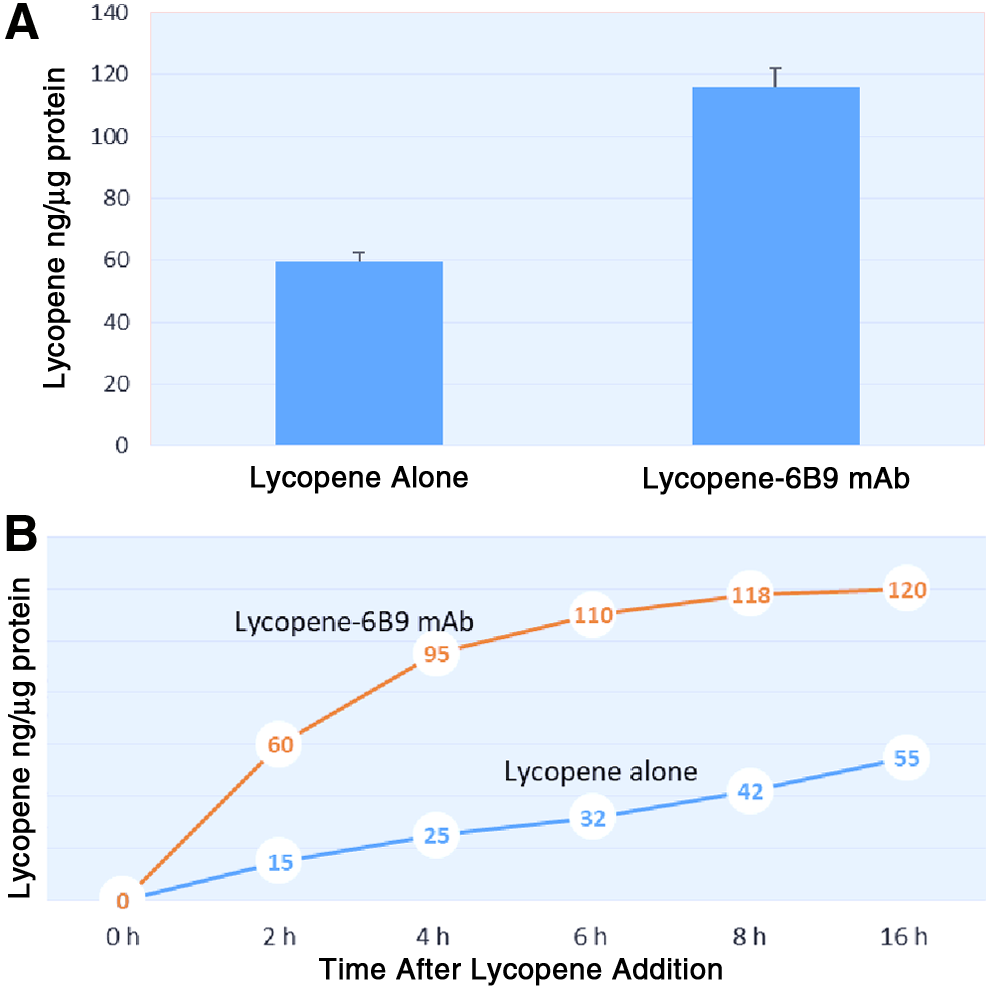
Lycopene content in microsomal fraction of B10.MLM cells incubated with lycopene–6B9 mAb complexes.
The time course study shown in Figure 1B revealed a different pattern of lycopene accumulation in B10.MLM cells treated with the two formulations of lycopene. Incubation with lycopene–6B9 mAb complex leads to a relatively steep and robust increase in the microsomal lycopene content as compared with incubation with DMSO-dissolved lycopene. In particular, lycopene content in the microsomal fraction of lycopene–6B9 mAb after a 2-hour incubation period reached 50% of the final value seen at the endpoint of the time course study. In contrast, incubation with DMSO-dissolved lycopene led to much slower dynamics of lycopene accumulation. Overall, lycopene content in the microsomal fraction of B10.MLM cells treated with lycopene–6B9 mAb complex was twice that in B10.MLM cells treated with DMSO-dissolved lycopene at the endpoint of the time course study (16 hours).
HMG-CoA reductase mRNA
The increased intracellular bioavailability of lycopene in the B10.MLM cells treated with lycopene–6B9 mAb complex reported above may have been accompanied by an increased effect of lycopene on cellular metabolism. Therefore, we decided to investigate changes in the mRNA of HMG-CoA reductase, which is known to be negatively regulated by lycopene.(24) Indeed, as can be seen in Figure 2, overnight incubation of the B10.MLM cells with DMSO-dissolved lycopene was accompanied by a measurable reduction of HMG-CoA reductase mRNA (15.0% of the control level). This decline was much more pronounced in the B10.MLM cells treated with lycopene–6B9 mAb complex (reduction of 29% as compared with the control level). Incubation with 6B9 mAb alone had no effect on HMG-CoA reductase mRNA.
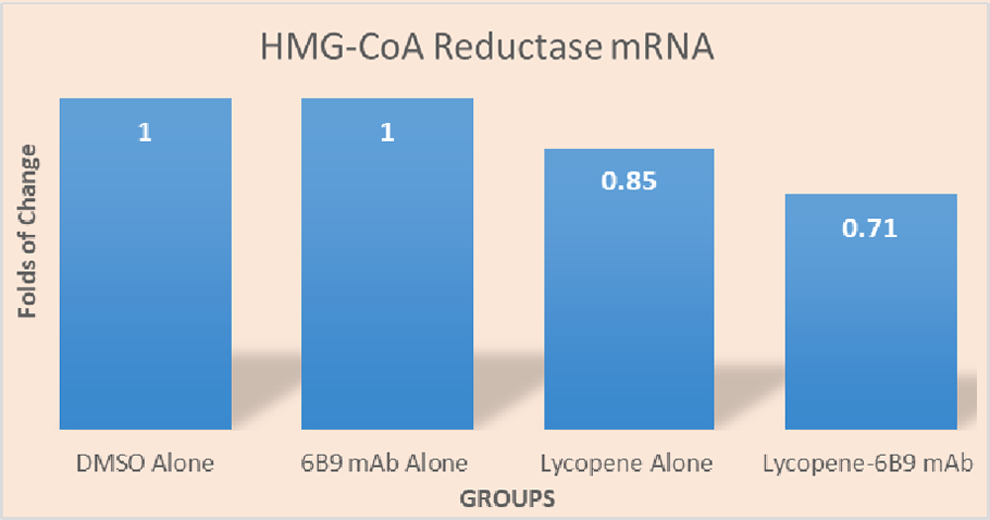
Changes in HMG-CoA reductase level in B10.MLM cells incubated with lycopene. B10.MLM cells were set, grown in duplicates, and incubated either with DMSO, 6B9 mAb, Lycopene–DMSO alone, or Lycopene–6B9 mAb complexes as described in the “Material and Methods” section. The cells were harvested, total RNA extracted, and real-time PCR analysis was performed after 16 hours of incubation. The results were normalized for human β-actin mRNA and plotted against HMG-CoA Reductase mRNA detectable in the cells incubated with DMSO alone (arbitrary value of 1.0). The study was conducted 3 times. The most representative set of results is shown above. HMG-CoA, 3-hydroxy-3-methylglutaryl-coenzyme.
Effect on C. trachomatis infection
As we have shown previously,(25) lycopene inhibits propagation of chlamydial pathogens in cultured cells. Therefore, we decided to investigate whether the increased cellular bioavailability of lycopene conferred by lycopene–6B9 mAb complex would be accompanied by increased inhibition of C. trachomatis infection in B10.MLM cells.
As can be seen from Figure 3, incubation with mAb 6B9 alone (Fig. 3B) did not affect the number of B10.MLM cells infected with C. trachomatis as compared with the control (Fig. 3A). However, incubation with DMSO-dissolved lycopene reduced the number of B10.MLM cells showing positive IF staining for chlamydial antigen (Fig. 3C). Furthermore, the greatest inhibition of C. trachomatis infection was seen in B10.MLM cells treated with lycopene–6B9 mAb complex (Fig. 3D) utilizing the same concentration of lycopene.

Lycopene inhibits propagation of Chlamydia trachomatis in B10.MLM cells. B10.MLM cells were set, grown in duplicates, infected with C. trachomatis and incubated with either DMSO
Lipid droplet formation
As we have shown in previous studies with B10.MLM cells, incubation with lycopene may lead to intracellular lipid droplet formation.(25) Therefore, we decided to investigate whether lycopene–6B9 mAb complex might affect lipid deposition in B10.MLM cells. As can be seen from Figure 4B, incubation with mAb 6B9 did not produce any detectable appearance of lipid droplets. However, overnight incubation with DMSO-dissolved lycopene caused formation of microvesicular lipid droplets (Fig. 4C). A greater number of lipid droplets (micro- and macrovesicular types) were observed in B10.MLM cells after overnight incubation with lycopene–6B9 mAb complex (Fig. 4D).
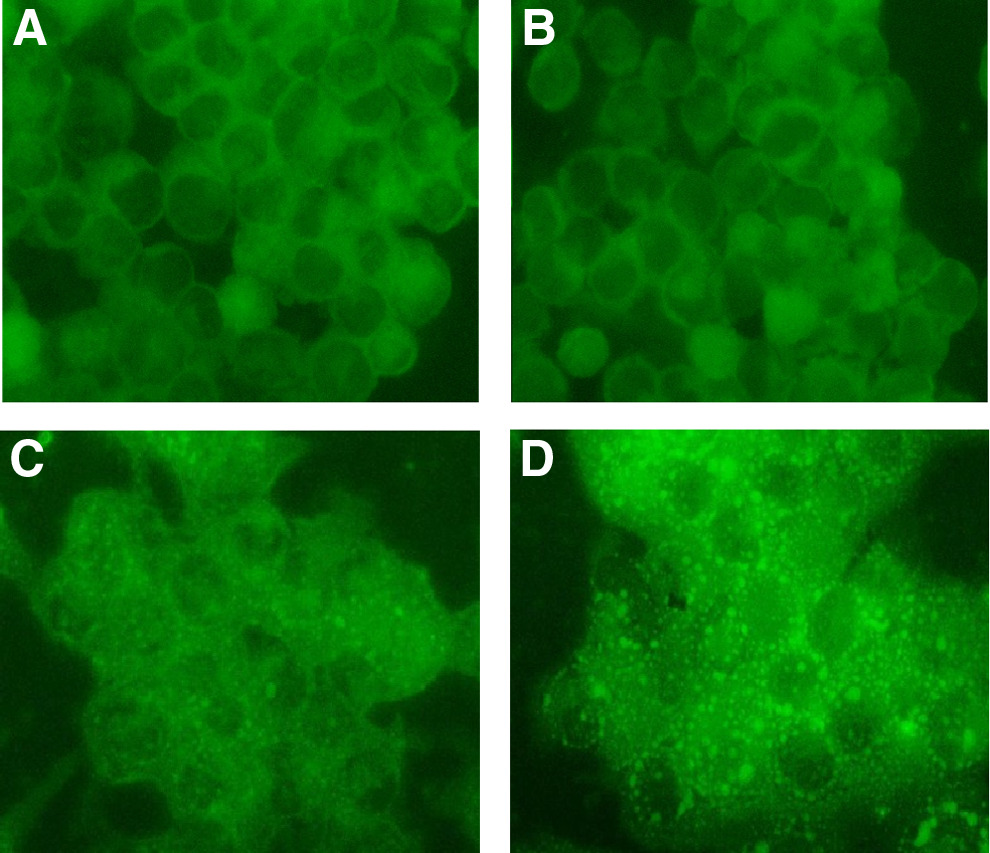
Lycopene-induced lipid droplet formation in B10.MLM cells. B10.MLM cells were set, grown in duplicates, and incubated with either DMSO
Discussion
There is an epidemiological link between circulating lycopene level and occurrence of CVD, prostate and breast cancers, and possibly other diseases.(4,5) However, low bioavailability of lycopene undermines its potential use as a therapeutic agent and confers poor reproducibility to clinical studies.(5) Intestinal absorption, as well as blood and transmembrane transport, represent three crucial levels controlling lycopene bioavailability within in vivo systems. First, during intestinal absorption a significant proportion of lycopene is lost due to irreversible enzymatic catabolism in the intestinal lumen and/or undergoes fecal excretion with indigestible plant matrix.(26) Second, to enter the systemic circulation and reach cells, lycopene needs to be incorporated into ApoB-containing lipoproteins and further cleared from the bloodstream by scavenger receptors, which are known to be impaired by aging, CVD, and cancer.(5,27) Low intestinal bioavailability and blood transport of lycopene can be at least partially improved by microencapsulation technologies, in particular lycosome technology.(28)
However, transmembrane transport of lycopene, the third mechanism limiting its bioavailability, remains poorly investigated. Passive transmembrane diffusion as well as receptor-mediated uptake by scavenger receptors is likely to be implemented in the mechanism of intracellular lycopene delivery.(9,14)
In the present article we show that overnight incubation of alveolar macrophages with a complex containing lycopene and complimentary monoclonal antibody (mAb 6B9) promotes accumulation of lycopene in the microsomal fraction of the cultured cells. Lycopene dissolved in an organic solvent (DMSO) was only half as effective in terms of increasing lycopene content in the microsomal fraction of B10.MLM cells when using the same lycopene concentration. These results allow us to suggest that lycopene can be delivered to the cultured cells using specific antibody–ligand complexes through receptor-mediated mechanisms with the possible participation of Fcγ-receptors. The possibility of receptor-mediated uptake involvement is supported by the results of the time course study conducted with lycopene–6B9 mAb complex. There was a steep and robust increase in microsomal lycopene content reaching a saturation plateau after 4 hours of incubation with lycopene–6B9 mAb complex, whereas incubation of B10.MLM cells with DMSO-dissolved lycopene led to a low magnitude and almost linear increase in microsomal lycopene content resembling the classic pattern of passive diffusion. However, deeper insight into the mechanism behind lycopene transmembrane transport in association with mAb is obviously required. Competition analysis and the possible use of mutant cell lines would be needed to gain a better understanding of the mechanisms mediating lycopene–mAb complex delivery.
Nevertheless, as can be seen from the results reported above, the increase in microsomal lycopene content seen in the B10.MLM cells incubated with lycopene–6B9 mAb complex was accompanied by important functional consequences revealing increased intracellular bioavailability of lycopene. First of all, there was substantial inhibition of mRNA for HMG-CoA reductase, a rate-limiting enzyme of cholesterol biosynthesis known to be negatively regulated by lycopene.(23,24) Second, there was improved inhibition of C. trachomatis infection in B10.MLM cells. Finally, incubation with lycopene–6B9 mAb complex promoted the formation of lipid-droplet phenotype in B10.MLM cells. In all three cases DMSO-dissolved lycopene demonstrated significantly less potency.
Special consideration needs to be given to the lycopene-induced lipid droplet formation observed in the B10.MLM cells. Generally, lipid droplet formation reflects impaired intracellular lipid transport as well as imbalance between lipid accumulation and lipid disposal in cultured cells.(29) A similar finding was reported previously in hepatic stellate cells treated with lycopene.(30) Lipid droplet formation is essential for lycopene storage in cells, since up to 51% of internalized lycopene is stored in lipid droplets.(31) Moreover, lycopene-induced lipid droplet formation may be very important for oxidative metabolism. As recently reported,(32) lipid droplet biogenesis protects mitochondrial function under conditions of nutrient deficiency. Ultrastructural studies conducted in our laboratory(25) and others(33) show that lipid droplets are often located in direct contact or close proximity to mitochondria and participate in oxidative metabolism by supplying fatty acid and possibly oxygen molecules to promote oxidative phosphorylation and counteract thereby the Warburg effect. Therefore, lycopene-induced lipid droplet formation may represent an important mechanism to decode the antineoplastic effects of lycopene in prostate and breast cancer cell lines reported elsewhere.(34,35) The effect of lycopene and lycopene–6B9 mAb complex on prostate cancer lines needs to be thoroughly addressed and is currently under our investigation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.