
Abstract
Ovarian tumor domain-containing protein 1 (OTUD1), an OTU-family deubiquitinating enzyme, has been reported to be involved in cancer progression through the regulation of p53 and SMAD7. However, the precise pathophysiological functions of OTUD1 remain elusive. Here, we report the establishment of OTUD1-specific monoclonal antibodies (mAbs), using the rat medial iliac lymph node method. The generated antibodies recognize the N-terminal portion (aa. 1–290) of human and mouse OTUD1 proteins. In addition, immunofluorescent staining and subcellular fractionation analyses using these antibodies indicated that OTUD1 is predominantly localized in the cytosol. Thus, these mAbs can be further used to elucidate cellular functions of OTUD1 and its involvement in processes such as cancer progression.
Introduction
T
Ovarian tumor domain-containing protein 1 (OTUD1; also known as DUBA7), a DUB of the OTU family, was identified as a biomarker of thyroid cancer.(6) OTUD1 deubiquitinates and stabilizes p53 tumor suppressor.(7) Moreover, OTUD1 upregulates expression of p21 and Mdm2, which accelerates apoptosis.(7) Recently, OTUD1 was shown to cleave Lys33-linked polyubiquitin chains from the transforming growth factor-β pathway inhibitor SMAD7, which is involved in the repression of breast cancer metastasis.(8) The results of these studies indicate that OTUD1 functions as a regulator of cancer progression. To further clarify the function of OTUD1, in this study, we established monoclonal antibodies (mAbs) against OTUD1 using the rat medial iliac lymph node method. These mAbs will be useful in immunoblotting and immunofluorescent staining of both human and mouse OTUD1 proteins.
Materials and Methods
Cell culture
HEK293T and HeLa cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS), 100 IU/mL penicillin G, and 100 μg/mL streptomycin under a humidified atmosphere with 5% carbon dioxide at 37°C. Subcellular fractionation of HEK293T cells was performed using NE-PER nuclear and cytoplasmic extraction reagents (Thermo Fisher Scientific).
Plasmid and transfection
For bacterial expression of glutathione S-transferase (GST)-tagged OTUD1, the complementary DNAs (cDNAs) encoding human OTUD1 (full length, aa. 1–290, and aa. 457–481) and mouse Otud1 (full length) were PCR amplified and cloned into pGEX-6P-1 (GE Healthcare). For ectopic expression of OTUD1 in HEK293T and HeLa cells, the cDNAs encoding human or mouse OTUD1 (full length) proteins were ligated to the appropriate epitope sequences and cloned into pcDNA3.1 (Invitrogen). Transfection to mammalian cells was performed with Lipofectamine 2000 (Thermo Fisher Scientific).
Expression and purification of recombinant proteins
Expression vectors of GST fusion proteins to hOTUD1 (full length, aa. 1–290, and aa. 457–481) and mOtud1 (full length) were transformed into Escherichia coli Rosetta2 (Novagen) or BL21(DE3) (Novagen). E. coli was grown in Luria-Bertani broth (LB) medium containing 50 μg/mL ampicillin at 37°C. Expression of the recombinant protein was induced by the addition of 0.4 mM isopropyl-1-thio-β-
Rat immunization and MAb production
The anti-OTUD1 rat MAb was generated based on the rat lymph node method established by Sado et al.(10) and Ohkawa et al.(11) A 10-week-old female WKY/Izm rat (SLC) was administered 200 μL of an emulsion containing 70 μg of mOtud1 (full length) and Freund's complete adjuvant in the hind footpads. After 19 days, the cells from the medial iliac lymph nodes of a rat immunized with an antigen were fused with mouse myeloma SP2 cells at a ratio of 5:1 in 50% polyethylene glycol (PEG 1500; Roche) solution. The resulting hybridoma cells were plated onto 96-well plates and cultured in HAT selection medium (Hybridoma-SFM [Invitrogen], 10% FBS, 5% BM-Condimed H1 [Roche], 100 mM hypoxanthine, 0.4 mM aminopterin, and 16 mM thymidine). At 8 days postfusion, the hybridoma supernatants were screened by enzyme-linked immunosorbent assay (ELISA) against GST-mOtud1. Positive clones were subcloned and rescreened by ELISA, immunoblotting, and immunofluorescent staining.
Enzyme-linked immunosorbent assay
GST-mOtud1 (full length) in ELISA buffer (20 mM sodium phosphate [pH 7.2]) was adsorbed on the surface of a Serocluster 96-well Nunc-Immuno™ Plate II through an overnight incubation at 4°C. To avoid nonspecific binding, the plates were blocked with 1% bovine serum albumin in TBS-T (20mM Tris-HCl buffer, pH 7.5, 150mM NaCl, and 0.05% Tween-20). Hybridoma supernatants were incubated overnight at 4°C, and then washed three times with TBS-T. The plates were incubated for 30 minutes at room temperature with an alkaline phosphatase-conjugated antirat IgG antibody (GE Healthcare) at a dilution of 1:5000. After washing with TBS-T, immunoreactivity was observed using a pNPP phosphatase substrate system (KPL).(12)
Immunoblotting
Cells were washed with phosphate-buffered saline (PBS) and lysed in lysis buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% Triton X-100, and complete protease inhibitor cocktail). The proteins were separated using 9% sodium dodecyl sulfate gels and electrophoretically transferred to polyvinylidene difluoride membranes. The membranes were blocked for 1 hour at room temperature with a blocking solution containing 5% skim milk in TBS-T, and then incubated overnight at 4°C with the 1/20-diluted anti-OTUD1 rat MAb containing hybridoma supernatants in the blocking solution. After washing with TBS-T, the membrane was incubated for 2 hours at room temperature with the horseradish peroxidase-conjugated antirat IgG antibody (GE Healthcare). After washing with TBS-T, the membrane was developed by treatment with SuperSignal West Pico (Thermo Fisher Scientific) or Luminata Forte Western HRP Substrate (Merck). For the detection of FLAG-tagged or GST-tagged proteins, anti-DYKDDDDK-tag (anti-FLAG) MAb peroxidase conjugated (Wako) or GST antibody (CST; No. 2622) was used. Antitubulin and anti-PARP antibodies were obtained from Cedarlane and Cell Signaling Technology, respectively.
Immunofluorescent staining
HEK293T and HeLa cells were washed with PBS and fixed with 4% paraformaldehyde for 15 minutes at room temperature. After washing with PBS, cells were permeabilized with 0.1% Triton X-100 containing PBS for 5 minutes, and then incubated in blocking buffer containing 10% FBS. The transiently expressed OTUD1 was detected with anti-FLAG, 7G4-A6, A8, and B12, followed by an Alexa 488-conjugated antirat IgG (Invitrogen) for 1 hour at room temperature. The nucleus was stained by 4′,6-diamidino-2-phenylindole (DAPI). The OTUD1-expressing cells were observed under a fluorescence microscope (BZ-9000; Keyence, Japan).
Results and Discussion
Human and mouse OTUD1 proteins contain 481 and 454 amino acid residues, respectively (Fig. 1A). These proteins show a high (∼75%) sequence similarity at the whole protein level. To establish mAbs specific for OTUD1, we utilized recombinant whole mOtud1 as an antigen. At 19 days postimmunization, lymphocytes were collected from the enlarged lymph nodes of the rat. The hybridomas, obtained after fusing the lymphocytes with mouse myeloma SP2 cells, were tested for the production of mAbs that react with GST-mOtud1, using ELISA. Some positive clones that were identified by ELISA were examined by immunoblotting to assess specificity to the lysates of HEK293T cells, which transiently expressed N-terminally FLAG-tagged hOTUD1 or mOtud1. mAbs 7G4 and 7B9 reacted to both human and mouse OTUD1 proteins, whereas 6C4 and 7F11 recognized only mOtud1 in immunoblotting (Fig. 1B). Compared with the immunoblot with the anti-FLAG antibody, these antibodies detected multiple lower molecular weight bands presumably due to the susceptibility of OTUD1 to intracellular protease cleavage.

Immunoblotting with anti-OTUD1 monoclonal antibodies.
Since 7G4 showed the strongest signals in immunoblotting, we subcloned and rescreened 7G4 by ELISA. Three positive clones, designated 7G4-A6, 7G4-A8, and 7G4-B12, were further examined by immunoblotting, and the results suggested that all the clones yielded strong signals against transiently expressed human and mouse OTUD1 proteins (Fig. 1C). The immunoglobulin class of these mAbs was determined as rat IgG2aκ. Moreover, immunoblotting using recombinant GST-fused hOTUD1 protein indicated that epitopes for these mAbs are present in the N-terminal portion (aa. 1–290) of hOTUD1 (Fig. 1D).
Next, we utilized these mAbs to analyze the intracellular localization of OTUD1. Immunofluorescence analyses and subcellular fractionation analysis using transiently expressed hOTUD1 in HEK293T cells indicated that hOTUD1 is predominantly localized in the cytosol, and small amounts of hOTUD1 are localized in the nucleus (Fig. 2A, B). Indeed, mAbs designed for OTUD1, 7G4-A6, 7G4-A8, and 7G4-B12 recognized the cytosolic localization of transiently expressed mOtud1 in HeLa cells (Fig. 2C).
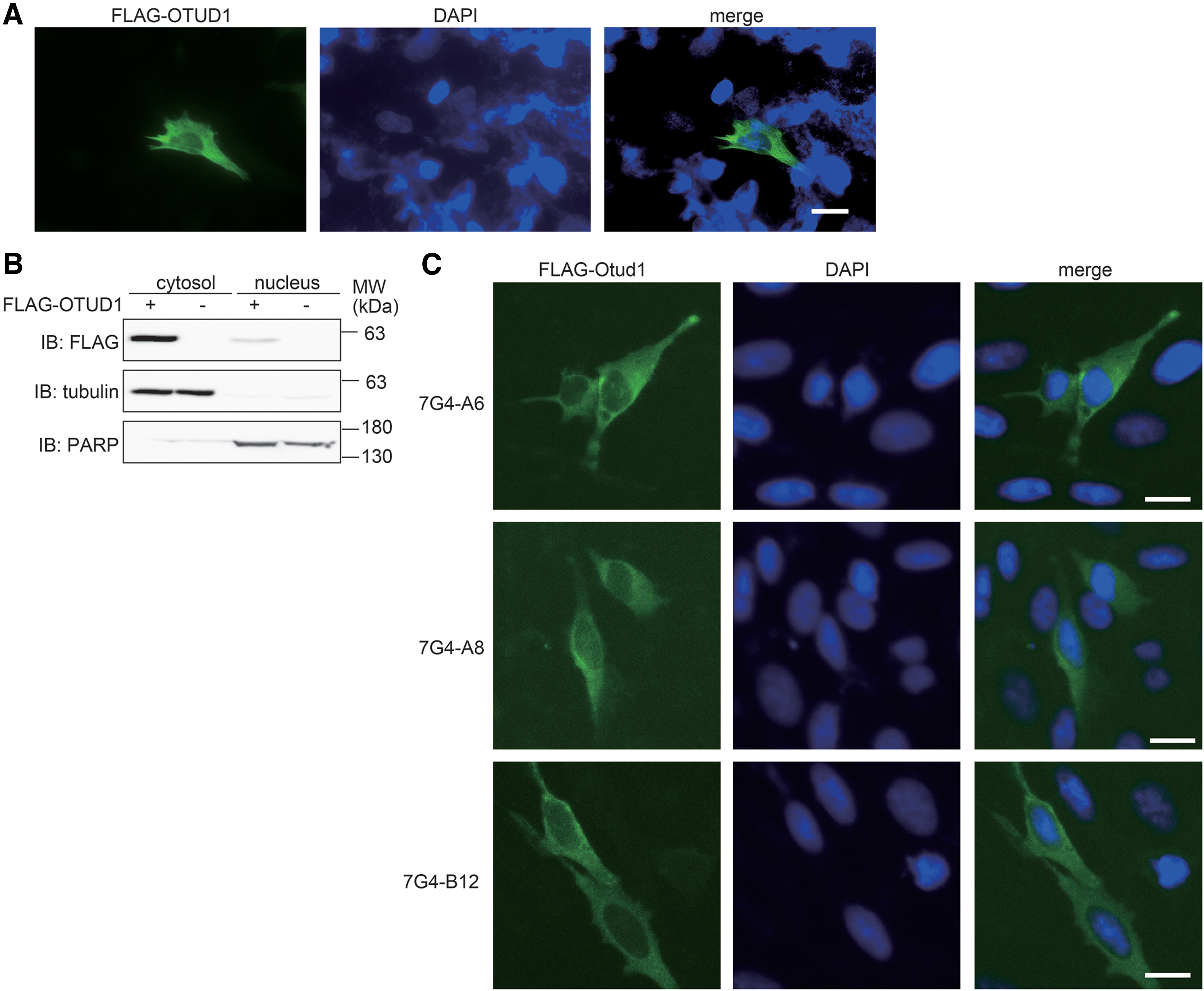
Cytosolic localization of OTUD1 in cells.
These findings demonstrate that we generated mAbs that specifically recognize the N-terminal portion of mouse and human OTUD1 proteins, and they could be useful tools for immunoblotting and immunofluorescence analyses. The use of these antibodies will allow further elucidation of the pathophysiological roles of OTUD1, such as in cancer progression and metastasis.
Footnotes
Acknowledgments
The authors thank Mr. Yuki Katayama (Gunma University) and Ms. Wakaba Koeda (Osaka City University) for technical assistance, and research support platform of Osaka City University Graduate School of Medicine for technical support. This work was partly supported by JSPS KAKENHI grants (18K06967 to D.O.; 16H06575 and 18H02619 to F.T.), Osaka City University Strategic Research Grant 2017 for top priority researches (F.T.), and Nakabayashi Trust for ALS Research (D.O.).
Author Disclosure Statement
No competing financial interests exist.