
Abstract
Type 1 diabetes mellitus is an autoimmune syndrome defined by the presence of autoreactive T and B cells, which results in destruction of insulin-producing beta cells. Autoantibodies against GAD65 (glutamic acid decarboxylase 65)—a membrane-bound enzyme on pancreatic beta cells, contribute to beta cells' destruction and the loss of pancreatic functions. Mouse FcγRIIb on B lymphocytes possesses an inhibitory effect on the activity of these cells. We hypothesized that it may be possible to suppress GAD65-specific B cells in mice with diabetes using chimeric molecules, containing an anti-FcγRIIb antibody, coupled to peptide B/T epitopes derived from the GAD65 protein. With these engineered chimeras, we expect to selectively co-cross-link the anti-GAD65-specific B cell receptor (BCR) and FcγRIIb, thus delivering a suppressive signal to the targeted B cells. An anti-FcγRIIb monoclonal antibody and two synthetic peptide epitopes derived from the GAD65 molecule were used for chimeras' construction. The suppressive activity of the engineered molecules was tested in vivo in mice with streptozotocin (STZ)-induced type 1 diabetes. These chimeric molecules exclusively bind disease-associated B cells by recognizing their GAD65-specific BCR and selectively deliver a strong inhibitory signal through their surface FcγRIIb receptors. A reduction in the number of anti-GAD65 IgG antibody-secreting plasmocytes and an increased percentage of apoptotic B lymphocytes were observed after treatment with protein-engineered antibodies of mice with STZ-induced type 1 diabetes.
Introduction
Type 1
Fc receptors are crucial for the interaction between effector cells and the antibody-bound antigen. Mouse myeloid cells, B cells, and natural killer cells are considered to have a vast array of Fc receptors on their surface, which are able to recognize Fc parts of immunoglobulins. Multiple receptor aggregation initiates tyrosine phosphorylation of signaling elements, which provides an intracellular signal. Immunoreceptor tyrosine-based activation motifs or immunoreceptor tyrosine-based inhibitory motifs (ITIMs) in the structure of receptors are responsible for transduction of activating or inhibitory signals inside the cell. So far, the only mouse FcγR possessing an ITIM is the FcγRIIb.(3,4)
The role of B cells in autoimmune diseases has been investigated and has recently attracted attention. Self-specific B cells play a key pathogenetic role in development of T1DM as producers of autoreactive antibodies and effective antigen-presenting cells.(5–9) Autoantibodies against membrane GAD65 molecules on pancreatic beta cells lead to their destruction and decline of pancreatic functions.(10,11) Therefore, selective elimination or suppression of anti-GAD65 B lymphocytes is a legitimate therapeutic goal.
Despite their autoreactivity, GAD65-specific B lymphocytes are also antigen driven and can be controlled in the same manner that controls proliferation and production of IgG antibodies to foreign antigens. FcγRIIb (CD32) is the only Fc-gamma receptor that B lymphocytes express, and cross-linking of the B cell receptor (BCR) with inhibitory FcγRIIb by IgG-containing immune complexes provides a mechanism for control of their activity.(12,13)
The 2.4G2 hybridoma produces IgG antibodies specific for mouse FcγRIIb. In previous studies, we have constructed several chimeric molecules that consist of self-epitopes (DNA mimotope or histone H1 peptide) conjugated to the 2.4G2 antibody. We have used the engineered protein chimeras for treatment of an experimental, lupus-prone, MRL/lpr mouse model. The specific binding to disease-associated B lymphocytes by cross-linking BCRs with inhibitory FcγRIIb receptors suppressed autoantibody production and lupus progression, prevented the appearance of proteinuria, and significantly prolonged survival.(14–16)
Several disease-prone or disease-induced animal models have been developed to study autoimmunity. A number of agents such as silicone, alum, many drugs, and different mineral oils are able to induce autoimmune symptoms in experimental mice.(17) Streptozotocin (STZ) has a preferential toxicity toward pancreatic beta cells and, at low doses, induces a diabetes-like syndrome in intact mice after multiple exposures. The ongoing immune inflammation and release of GAD65 autoantigens after STZ challenge result in beta cell destruction, followed by hyperglycemia, and inflammatory cells infiltrate the pancreatic islets.(18,19)
In the present research, we have investigated the possibility of suppressing anti-GAD65 IgG antibody-producing B lymphocytes in an STZ-induced C57BL/6J murine model of T1D by chimeric molecules comprising GAD65-derived peptide sequences and the 2.4G2 monoclonal antibody. These protein-engineered molecules should target GAD65-reactive B cells by co-cross-linking their GAD65-specific BCR and the inhibitory FcγRIIb receptor to selectively silence them.
Materials and Methods
Prediction of Major Histocompatibility Complex binders by EpiDOCK
EpiDOCK is a server for prediction of peptides binding to HLA class II proteins.(20) It uses quantitative matrices derived from docking-based analyses of peptide-HLA class II complexes. EpiDOCK predicts binding to 23 most frequent HLA class II proteins: 12 HLA-DR, 6 HLA-DQ, and 5 HLA-DP. In the present study, EpiDOCK was used to predict binders to the type 1 diabetes-susceptible alleles: DQ8 (DQA1*03:01/DQB1*03:02), DRB1*04:01, and DRB1*04:05. Default thresholds were used: 0.1 for DQ8 and 0.3 for DRB1*04:01 and DRB1*04:05. EpiDOCK was used to determine two epitopes of the GAD65 molecule—p121–140 and p270–289.
Monoclonal antibodies
Purified rat anti-mouse 2.4G2 monoclonal antibody specific to mouse FcγRIIb was prepared as described.(14) Briefly, the rat 2.4G2 hybridoma (ATCC HB-197) producing the IgG2b antibody was adapted to grow in the serum-free CHO medium. Antibodies from the supernatant were isolated by ammonium sulfate precipitation and subsequent dialysis. Their purity was determined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) using a PageBlue™ protein staining solution (Thermo Fisher Scientific, Waltham, MA). The 2.4G2 antibody was used for construction of chimeric molecules.
Anti-mouse phycoerythrin (PE) or eFlour450-conjugated CD19 (eBioscience, Frankfurt, Germany) and anti-mouse CD32 (Abcam, Cambridge, United Kingdom) antibodies were used for fluorescence-activated cell sorting (FACS) experiments.
Construction of chimeric molecules
Two designed peptides from the human GAD65 molecule—p121–140 and p270–289—were synthesized using Fmoc chemistry on resin with >96% purity from Caslo Laboratory (Lyngby, Denmark). The amino acid sequences were p(121–140) Ac-YVVKSFDRSTKVIDFHYPNE-Ahx-K-CONH2 and p(270–289) Ac-LPRLIAFTSEHSHFSLKKGA-Ahx-K-CONH2. For initiation of protein conjugation, a lysine carrying the Ahx linker was introduced to the C-end of peptides during synthesis. The chimeric molecules were constructed by separate conjugation of peptides to anti-FcγRIIb antibodies using the EDC [1-ethyl-3(3′-dimethylaminopropyl) carbodiimide.HCl] (Sigma-Aldrich, Taufkirchen, Germany) cross-linking procedure, as described.(14,21) The resultant engineered protein molecules, Chimera1 and Chimera2 (consisting of the p121–140 or p270–289 peptides, respectively), were used in the next experiments.
Mice
Eight-week-old, female C57BL/6J mice were obtained from the Jackson Laboratory (Bar Harbor, ME). The animals were kept under specific pathogen-free conditions, and manipulations were approved by the Animal Care Commission at the Institute of Microbiology in accordance with national regulations (EU Directive 2010/63/EU).
STZ-induced type 1 diabetes
STZ (Sigma-Aldrich) was dissolved in 25 mM sodium citrate/citric acid buffer (pH 4.0) and injected into the mice intraperitoneally (i.p., 40 mg/kg) within 15 minutes of preparation. Mice were treated with five consecutive daily injections and with a single boost (150 mg/kg) 45 days after the first injection. The blood glucose levels of experimental animals were tested weekly using a blood glucose meter Accu-Chek (Roche Diabetes Care). Mice with a blood glucose concentration exceeding 250 mg/dL (13.9 mmol/L) were considered diabetic.
Treatment schedule
A group of 8-week-old, female C57BL/6J mice (n = 10) with STZ-induced diabetes was immunized i.p. weekly with 50 μg/mouse of chimeric molecules (25 μg Chimera1 + 25 μg Chimera2), starting on day 52 after the first STZ challenge (Fig. 1A). A control animal group (n = 10) with STZ-induced diabetes was injected with phosphate-buffered saline (PBS) only under the same schedule. Another control group (n = 10) with STZ-untreated mice was injected weekly with PBS. Every week, the animals from all groups were bled and blood glucose levels were measured.
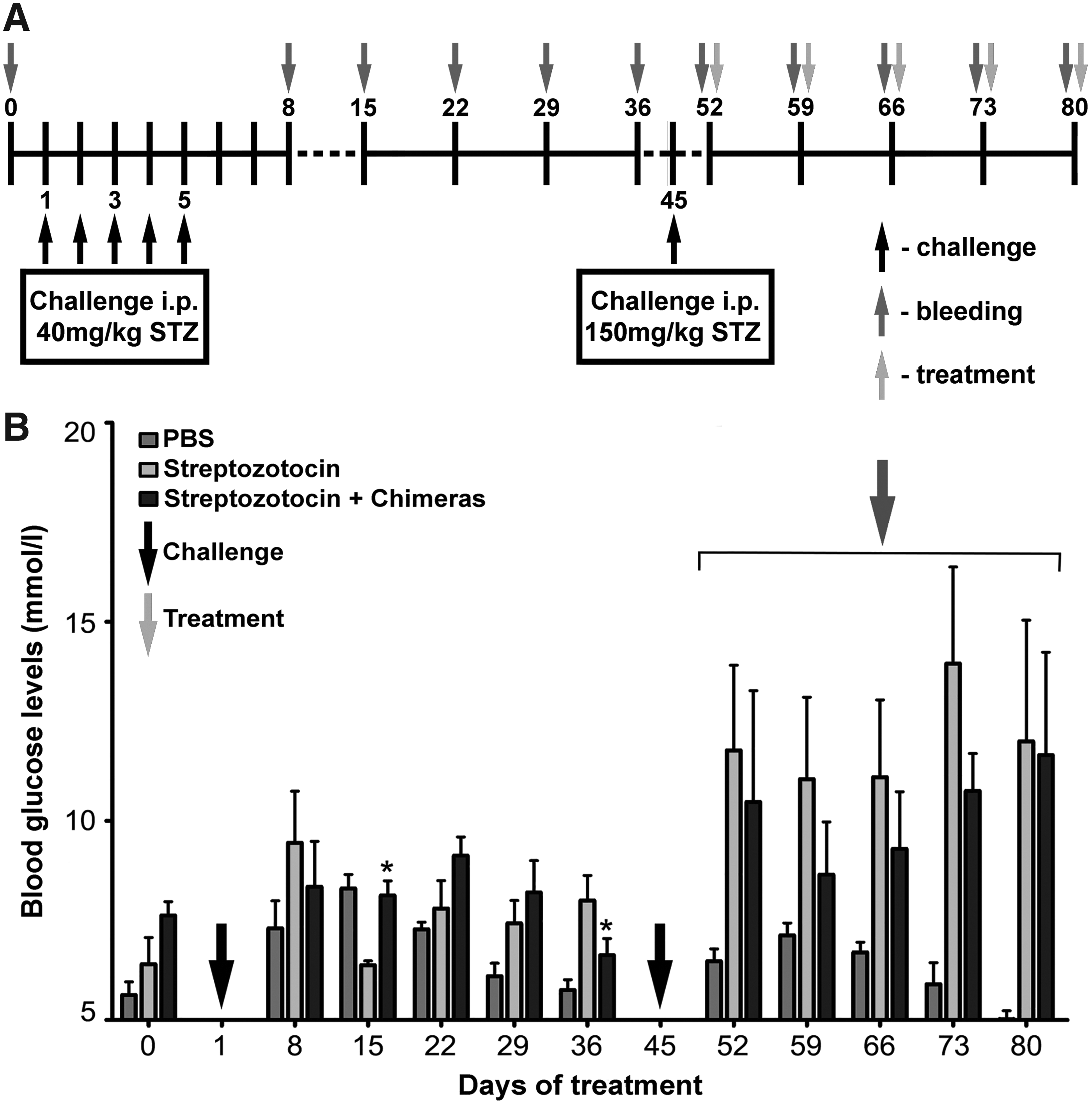
Development of an STZ-induced model of diabetes in C57BL/6J mice.
Flow cytometric analysis
Spleens from STZ-treated mice were taken and the monocellular suspension was isolated by cell strainers (BD Biosciences, Erembodegem, Belgium). The cells were washed with PBS (containing 2.5% fetal calf serum [FCS] and 0.05% sodium azide) and incubated with PBS only or with Chimera1 or Chimera2 (1 μg/106 cells), followed by two washes. Next, the cells were incubated with the fluorescein isothiocyanate (FITC)-conjugated anti-FcγRIIb antibody combined with anti-CD19-PE antibody. Each incubation step was performed for 20 minutes on ice. Ten thousand cells were analyzed from each sample with a BD LSR II flow cytometer using the Diva 6.1.1. software (BD Biosciences, San Jose, CA).
Detection of apoptosis
For detection of apoptosis, splenocytes from STZ-treated sick animals or intact untreated controls (day 80) were isolated and cultured for 3 days (2 × 106 cells/mL) in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma-Aldrich) supplemented with 10% FCS and 4 mM
Enzyme-linked immunospot assay for counting specific anti-GAD65 IgG antibody-secreting B cells
Isolated splenocytes from the STZ-treated mice or untreated controls were cultured (2 × 106 cells/mL) for 3 days in complete RPMI 1640 medium with different concentrations of protein chimeric molecules, as described above. Cells stimulated with a 10-μg mix of both peptides (5 μg p121–140 peptide +5 μg p270–289 peptide) or with 5 μg/mL lipopolysaccharide (LPS) (from Escherichia coli; Sigma) or cultured in medium only were used as controls.
Enzyme-linked immunospot (ELISpot) 96-well plates (Millipore, Bedford, MA) were activated with 35% ethanol, coated with 10 μg/mL GAD65 peptides (5 μg p121–140 + 5 μg p270–289) for overnight/4°C, washed with PBS, and blocked with 1% gelatin in PBS. Preincubated cells from the culture plates were transferred to ELISpot plates with peptide-coated membranes, and the cells were further cultured for an additional 4 hours in a humidified 5% CO2 atmosphere at 37°C. After washing, the ELISpot plates were incubated with an alkaline phosphatase-conjugated anti-mouse IgG antibody for 1 hour and developed by NBT/BCIP substrate (Sigma). The number of plasmocytes producing IgG anti-GAD65 antibodies was (detected as colored spots on the membrane) counted by the CTL Immunospot S5 Versa Analyzer (Bonn, Germany).
Statistical analyses
All statistical analyses were performed with Prism software from GraphPad (San Diego, CA). The two-way ANOVA (analysis of variance) test was used to determine differences between the two groups. Values in the figures correspond to mean ± standard deviation. A value of p < 0.05 was considered as statistically significant.
Results
Generation of protein-engineered chimeric molecules
Using EpiDOCK, a GAD65-derived peptide 121–140 (containing 7 DQ8 and 1 DRB1*04:05 binders) and a peptide 270–289 (containing 6 DQ8 and 2 DRB1*04:05 binders) were selected for generation of protein chimeric molecules. Several copies of both synthesized peptides were chemically conjugated separately to the monoclonal anti-mouse FcγRIIb antibody 2.4G2.
The protein chimeric molecules decreased blood glucose levels in C57BL/6J mice with STZ-induced diabetes
Administration of the chimeric molecules to animals challenged with STZ resulted in insignificant delay of the sharp increase of blood glucose levels during the next 5 weeks, compared with the PBS-treated STZ-challenged group (Fig. 1B). After 5 weeks of treatment, the appearance of anti-rat immunoglobulin antibodies was observed, thus the period of chimeras' administration was limited to that point.(15) A rapid increase of blood glucose levels was observed in the PBS-treated STZ-challenged controls during the same period after the second STZ challenge.
The chimeric molecules retain their binding capacity to the targeted receptors
Further on, we tested the target receptor specificity of engineered chimeric molecules (Chimera1 and Chimera2) by competitive FACS analysis with commercial FITC-conjugated anti-FcγRIIb antibody to the gated CD19+ mouse cells from STZ-treated animals. Both chimeras retained their receptor-binding capacity and inhibited the binding of an FITC-conjugated antibody with the same specificity (Fig. 2A).

The GAD65 chimeras 1/2 retain their ability to bind mouse B lymphocytes and to induce apoptosis.
Chimeric molecules increased the percentage of apoptotic B lymphocytes from STZ-challenged mice
Isolated splenocytes from STZ-treated animals or healthy controls were used to investigate the proapoptotic effect of GAD65 chimeras. The level of phosphatidylserine surface expression on CD19-gated lymphocytes was analyzed by flow cytometry after incubation for 3 days with different concentrations of protein chimeric molecules. Both GAD65 chimeras induced increased dose-dependent apoptosis levels in B cells from STZ-treated mice; however, the results obtained after Chimera2 treatment were more prominent (Fig. 2B). Lymphocytes isolated from STZ-untreated mice were not affected after chimeric molecule treatment (data not shown).
The constructed chimeric molecules suppressed the GAD65-specific B cell differentiation to autoantibody-secreting plasmocytes
The effects of chimeric molecules on the number of IgG anti-GAD65 antibody-producing plasma cells were assessed by ELISpot. STZ-treated C57BL/6J mice have large numbers of GAD65-specific B and plasma cells. The isolated splenocytes from all groups were cultured for 3 days, and numbers of anti-GAD65 IgG antibody-producing cells were counted by the ELISpot technique (Fig. 3). Increasing concentrations of the constructed protein chimeras were added to splenocytes and cells were cocultured in complete RPMI 1640 medium without B and T cell stimulation. The treatment of splenocytes with both chimeras caused a dose-independent decrease in the numbers of in vitro-differentiated anti-GAD65 IgG antibody-producing cells in STZ-challenged groups either treated or not in vivo with the chimeric molecules.
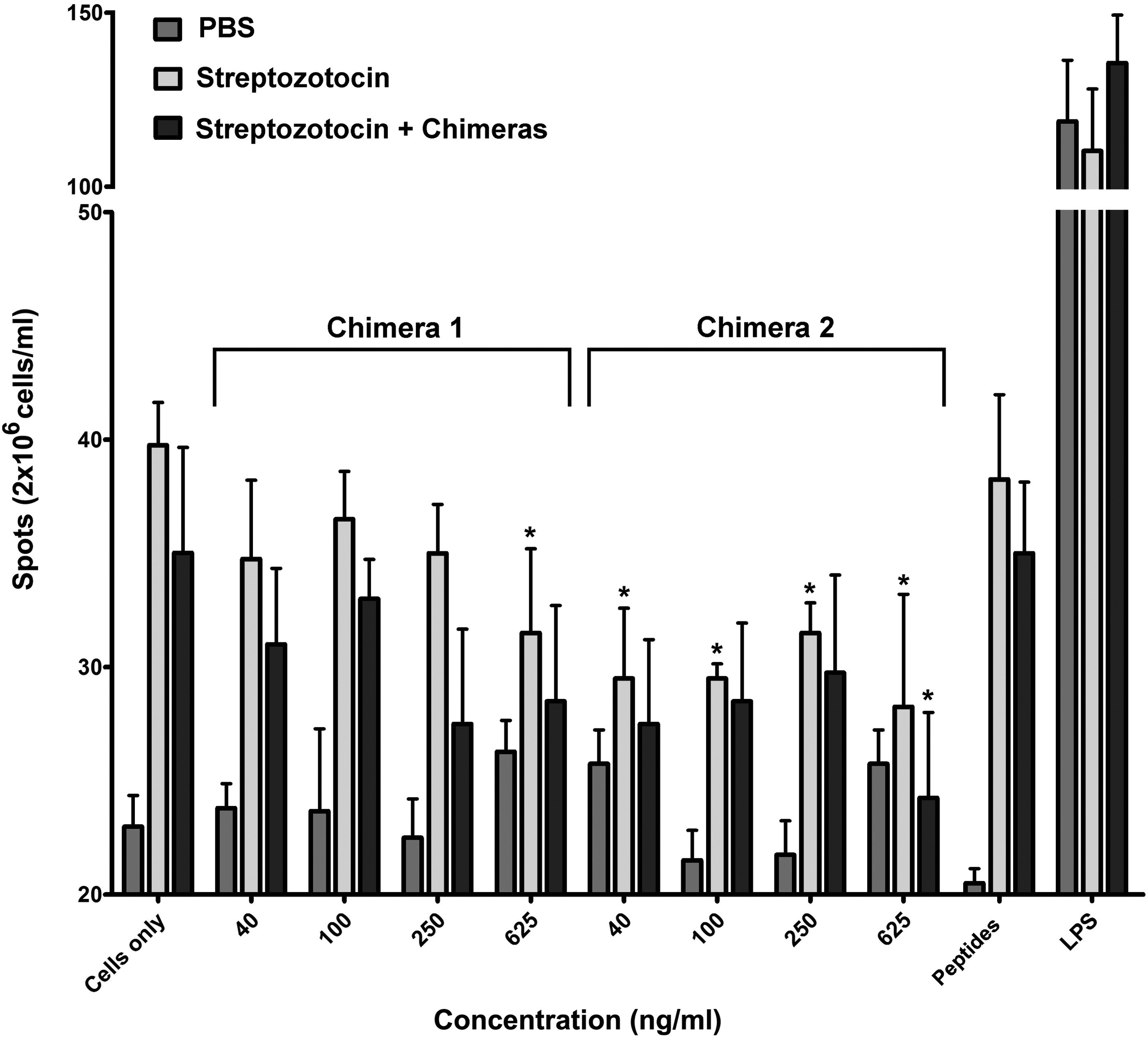
The GAD65-specific chimeras affect the differentiation of targeted GAD65-specific B cells. Spleen cells from all groups of C57BL/6J mice with diabetes were cultured in medium alone or cocultured with the Chimera1 or with Chimera2 for 3 days. Control cells were stimulated with combined GAD65-derived peptides or with LPS. The number of spots corresponding to the number of anti-GAD65 IgG antibody-secreting plasma cells was determined by ELISpot assay and compared with untreated cells (medium only). Results are expressed as the mean value ± SD of triplicated assays. p-Values were calculated using the two-way ANOVA test (*p < 0.05). Data are representative of four independent experiments. ELISpot, enzyme-linked immunospot; LPS, lipopolysaccharide.
Addition of bacterial LPS further increased the numbers of antibody producers with the studied specificities. In contrast, addition of both GAD65-derived peptides did not further increase the numbers of antibody producers, showing that most B cells with the studied specificities were already activated.
The same experiments were performed using splenocytes isolated from STZ-untreated mice as controls. The background of the lower number of GAD65-specific plasmocytes was weakly affected by chimeric molecule treatment (data not shown).
Discussion
The ligands of mouse Fc receptors can be secreted antibodies (FcμR, FcγR, and FcɛR) or intracellular immunoglobulins (FcRn and TRIM21).(22) Based on the overall effect of the interaction, FcRs are divided into activating, inhibitory, and routing/transport Fc receptors(4)—a varied cohort of Fc receptors. Fc gamma receptors recognize different subclasses of IgG molecules and (based on that) are shared into four groups—FcγRI, FcγRIIb, FcγRIII, and FcγRIV. Most Fcγ receptors are responsible for cell activation and cytokine secretion and provide proliferative stimuli, being affected by the antigen–antibody complex. These evidences are in contrast to the inhibitory activity triggered with FcγRII on B lymphocytes.(23) Although there have been researches showing that FcγRIIb can be expressed on T cells, Fc gamma RIIB is considered to be expressed mainly on macrophages, FDCs, and B cells.(23–26) Downregulation through FcγRIIb is able to affect not only the cell itself but other cells as well through intercellular mechanisms.(27,28)
The present therapy of autoimmune diseases is nonspecific and it is mainly based on administration of immunosuppressive and anti-inflammatory drugs, which have severe side effects. Monoclonal antibodies are another option and they have been widely used for the treatment of various autoimmune conditions. The therapy is cell targeted and affects T and B lymphocytes, but the mechanisms are still not well understood. Several attempts have been made by using monoclonal antibodies recognizing B cell-specific surface molecules, such as CD20, CD22, BAFF (B cell-activating factor), and APRIL (a proliferation-inducing ligand).(29–32) Despite some beneficial results, these approaches are not specific as they deplete all the B cells regardless of their antigen-binding specificity, affecting the protective function of the immune system.
The difference in presentation of GAD65 CD4 T cell epitopes depending on their localization into the protein molecule has been shown before. The autoreactive B cell processing of T cell epitopes built in the frame of B cell epitopes results in ineffective antigen presentation and blocking or inhibition of the response from respective autoreactive T cells.(33,34) In contrast with this phenomenon, T cell epitopes without antibody overlaying proceed to normal antigen presentation, followed by a developing autoimmune response. The authors speculated that suppression of T cell epitope's presentation in the antibody-binding region is covered by epitope-masking GAD65-specific autoantibodies, a process that stabilizes and sets apart these epitopes.
In the present study, we take advantage of the opportunity to use two peptides comprising T cell epitopes that will bind self-reactive B lymphocytes. We aimed to prove that the specific cell-silencing approach based on engaging inhibitory receptors on targeted pathological B lymphocytes has therapeutic potential. A possible mechanism for control on the magnitude and duration of the IgG antibody response to self-antigens is to co-cross-link the inhibitory FcγRIIb and BCR on the surface of an autoreactive B lymphocyte by bifunctional protein chimeric molecules. This co-cross-linking leads to specific B cell downregulation and prevention of B cell differentiation to autoantibody-producing plasmocytes. Our data from previous studies support the suggestion that bispecific chimeric molecules that contain copies of the dsDNA-mimotope peptide coupled to an anti-mouse FcγRIIb antibody suppressed the self-specific B cell development and disease progression in lupus-prone MRL/lpr mice.(14–16) A similar approach was used here for generation of GAD65-specific chimeric molecules consisting of GAD65-derived peptide epitopes coupled to the same anti-mouse FcγRIIb antibody. The number of potentially coupled GAD65 peptides (with a mass of 2400 Da) is expected to be between 12 and 15 copies for a single immunoglobulin molecule. The process has been determined by mass spectrometry analysis of similar peptide–antibody chimeras constructed by us in the same way.(15) During chemical conjugation, the existing free carboxyl groups on the surface of the antibody molecule are potentially available for interaction with the reactive H2N group of the Ahx linker bound to the C-end of synthetic peptides. Some of these carboxyl groups are within the antigen-binding pockets of the antibody and may have a potential effect on antibody affinity if bound. We proved the retained, intact antigen-binding capability of generated protein chimeras by competitive FACS analysis against FITC-conjugated anti-FcγRIIb antibody.
The aggregate binding avidity of bispecific molecules should be equal to the product of the two values of individual receptor affinities—that of the GAD65 epitope/GAD65-specific BCR and 2.4G2 antibody/FcγIIb. Even if individual receptor values are low, the overall avidity will be high and biologically significant. Under in vivo administration, the preferred targets of protein chimeric molecules will be disease-associated GAD65-specific B lymphocytes. Nevertheless, it is inevitable that some of the GAD65-specific chimeric molecules will bind to nontargeted cells that express surface FcγIIb only, but we cannot expect to provide a single-signal biological effect.
Previously, we have shown that chimeric molecule treatment leads to the development of anti-rat immunoglobulin antibodies in all treated mice between the sixth and eighth week of the treatment.(14) This evidence was a limiting factor, which restricts the term of treatment of STZ-challenged mice with GAD65-specific engineered molecules. This span of chimeras' administration was able to block the replenishment of the pool of disease-associated plasma cells suppressing GAD65-specific B cell differentiation by induction of anergy, followed by apoptosis. Similar results were obtained using epitope-specific, protein chimeric molecules in murine models of lupus and house dust mite allergy.(14,14,35)
The STZ model of induced diabetes presented a wide range of disease symptoms in the challenged group of animals. In silico generated two GAD65 epitopes represent part of potential targets among the anti-GAD BCR repertoire. In our experiments, the treatment of mice with STZ-induced diabetes with GAD65 chimeric molecules delayed disease progression and suppressed disease-associated B lymphocytes. This selective approach for elimination of the existing low number of epitope-specific B lymphocytes cannot suspend diabetes development, but demonstrates a possible mechanism for specific therapy of autoimmune diabetes.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The work was supported by a grant NFDP 53/27.04.16 from the Program for Support of Young Researchers and PhD Students, Bulgarian Academy of Sciences.