
Abstract
The classic method for identifying the epitope that monoclonal antibodies (mAbs) bind uses deletion mutants and point mutants of the target protein. However, determining the epitope of mAbs-reactive membrane proteins is often challenging. We recently developed the RIEDL insertion for epitope mapping (REMAP) method to identify mAb-binding epitopes. Herein, we first checked the reactivity of an anti-epidermal growth factor receptor (EGFR) mAb (EMab-51) to several EGFR deletion mutants such as EGFR/dN152, EGFR/dN313, EGFR/dN370, EGFR/dN375, EGFR/dN380, and EGFR/dN482. We found the N-terminus of the EMab-51-binding epitope between residues 375 and 380 of EGFR. We next produced EGFR/dN313 mutants with the RIEDL peptide tag inserted at each possible position of 375-AFRGDSFTHTPPLDP-389. EMab-51 lost its reactivity with the mutants having a RIEDL tag inserted at each position of 377-RGDSFTHTPP-386, whereas LpMab-7 (an anti-RIEDL mAb) detected every mutant. Thus, using the REMAP method, we identified the EMab-51-binding epitope of EGFR as 377-RGDSFTHTPP-386.
Introduction
The epidermal growth factor receptor (EGFR) belongs to the human epidermal growth factor receptor (HER) family.(1) The HER family is composed of four different receptors with a common structure, including EGFR (HER1/ErbB1), HER2 (ErbB2/neu), HER3 (ErbB3), and HER4 (ErbB4).(2) When the ligand (EGF) binds the EGFR, it stabilizes the receptor dimer. The two subunits phosphorylate each other in the C-terminal region of the cytoplasmic side, which transmits signals downstream.(3,4) Besides, EGFR can form homodimers or heterodimers with the other HER family members (HER2, HER3, and HER4).(5,6) The downstream effectors of EGFR include PI3K/AKT/mTOR, RAS/RAF/MAPK, and JAK/STAT, which act on division, migration, anti-apoptosis, and cell cycle progression.(7) Many cancers overexpress EGFR, including squamous cell carcinoma of the esophagus,(8) brain tumor,(9,10) lung cancer,(11) head and neck cancer,(12) colorectal cancer,(13,14) breast cancer,(15) bladder cancer,(16) clear cell renal cell carcinoma,(17) ovarian cancer,(18) prostate cancer,(19) pancreatic cancer,(20) and melanoma.(21) Therefore, monoclonal antibodies (mAbs) that can detect EGFR with high sensitivity are essential.
Because mAbs usually recognize epitopes consisting of several amino acids, they often might cross-react with unexpected proteins. Therefore, identifying the epitopes that mAbs recognize is important to avoid unexpected cross-reactivity, and is helpful for the development of antibody drugs. Epitope identification methods include site-directed mutagenesis mapping, array-based oligopeptide scanning, and X-ray co-crystallography.(22) Because X-ray co-crystallography allows to directly visualize the mAb-antigen interaction, it gives the clearest epitope identification. However, crystallizing the antigen-mAb complex is usually costly and time-consuming. Although the array-based oligopeptide scanning and site-directed mutagenesis mapping can easily identify linear epitopes, they are not appropriate for conformational epitopes.
We previously developed a novel anti-EGFR mAb, EMab-51 (IgG1, kappa),(23) by immunizing mice with the purified recombinant ectodomain of EGFR (EGFRec) from culture supernatants of LN229/EGFRec cells. Importantly, this mAb is useful for flow cytometry, Western blotting, and immunohistochemical analyses against EGFR. Accordingly, a practical use of EMab-51 will likely be EGFR detection in various tumors. However, we failed to identify the EMab-51-binding epitope using conventional epitope-mapping methods such as alanine scanning. In this study, we aimed to identify the EGFR epitope recognized by EMab-51 using the RIEDL insertion for epitope mapping (REMAP) method.(24)
Materials and Methods
Cell lines
We obtained Chinese hamster ovary (CHO)-K1 cells from the America Type Culture Collection (ATCC, Manassas, VA). We transfected the EGFR mutation plasmids into CHO-K1 cells using Neon Transfection System (Thermo Fisher Scientific, Inc., Waltham, MA) and sorted the stable transfectants using an anti-PA tag mAb (NZ-1) with a cell sorter (SH800; Sony Corp., Tokyo, Japan). We cultured the CHO-K1 cells and transfectants in RPMI 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan) supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL of penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified atmosphere containing 5% CO2. We cultivated the transfectants in a medium containing 0.5 mg/mL Zeocin (InvivoGen, San Diego, CA).
Plasmid preparation
We amplified the EGFR open reading frame as described previously.(25) We produced the EGFR deletion mutants (EGFR/dN152, EGFR/dN313, EGFR/dN370, EGFR/dN375, EGFR/dN380, and EGFR/dN482) using HotStar HiFidelity Polymerase Kit (Qiagen, Inc., Hilden, Germany) with oligonucleotides containing the desired mutations. At the N-terminus, we added the PA tag (GVAMPGAEDDVV),(26) which is recognized by NZ-1.(27) We inserted the RIEDL tag(24) in the EGFR sequence using a HotStar HiFidelity Polymerase Kit with oligonucleotides containing the RIEDL tag insertions at the desired position in 375-AFRGDSFTHTPPLDP-389 of EGFR/dN313. For example, we produced Ala375_RIEDL_Phe376 (A375_R*_F376) by inserting the RIEDL sequence between Ala375 and Phe376 of EGFR/dN313. We inserted the polymerase chain reaction fragments bearing the desired mutations into the pCAG-Ble vector using an In-Fusion HD Cloning Kit (Takara Bio, Inc., Shiga, Japan). The RIEDL tag insertion mutants produced are Ala375_RIEDL_Phe376 (A375_R*_F376), Phe376_RIEDL_Arg377 (F376_R*_R377), Arg377_RIEDL_Gly378 (R377_R*_G378), Gly378_RIEDL_Asp379 (G378_R*_D379), Asp379_RIEDL_Ser380 (D379_R*_S380), Ser380_RIEDL_Phe381 (S380_R*_F381), Phe381_RIEDL_Thr382 (F381_R*_T382), Thr382_RIEDL_His383 (T382_R*_H383), His383_RIEDL_Thr384 (H383_R*_T384), Thr384_RIEDL_Pro385 (T384_R*_P385), Pro385_RIEDL_Pro386 (P385_R*_P386), Pro386_RIEDL_Leu387 (P386_R*_L387), Leu387_RIEDL_Asp388 (L387_R*_D388), and Asp388_RIEDL_Pro389 (D388_R*_P389).
Flow cytometry
We harvested the cells by brief exposure to 0.25% trypsin/1 mM ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.). After washing in 0.1% bovine serum albumin in phosphate-buffered saline (blocking buffer), the cells were treated with primary mAbs, such as EMab-51 (mouse IgG1, kappa), NZ-1 (rat IgG2a, lambda), or LpMab-7 (mouse IgG1, kappa) at a concentration of 1 μg/mL for 30 minutes at 4℃; subsequently, with Alexa Fluor 488-conjugated anti-mouse IgG or Alexa Fluor 488-conjugated anti-rat IgG (1:1000; Cell Signaling Technology, Inc., Danvers, MA). We collected fluorescence data using an EC800 Cell Analyzer (Sony Corp.).
Results
Determination of the EMab-51-recognized epitope using EGFR deletion mutants
EMab-51 might recognize an epitope located at the extracellular region of EGFR because we obtained it by immunizing mice with EGFRec from culture supernatants of LN229/EGFRec cells.(23) First, we produced N-terminal deletion mutants (EGFR/dN152, EGFR/dN313, and EGFR/dN482) with a PA tag at their N-terminus and investigated the reactivity between EMab-51 and each deletion mutant by flow cytometry analysis. EMab-51 recognized EGFR/wild type, EGFR/dN152, and EGFR/dN313, but not EGFR/dN482 (Fig. 1A). In contrast, NZ-1 detected all the deletion mutants (EGFR/dN152, EGFR/dN313, and EGFR/dN482) (Fig. 1B). These results show that the N-terminus of the EMab-51 epitope exists between residues 313 and 482 (Fig. 1C).

Epitope mapping using EGFR deletion mutants.
Next, we produced additional N-terminal deletion mutants (EGFR/dN370, EGFR/dN375, and EGFR/dN380) with a PA tag at their N-terminus and investigated the reactivity between EMab-51 and each deletion mutant by flow cytometry analysis. EMab-51 recognized EGFR/dN370 and EGFR/dN375, but not EGFR/dN380 (Fig. 2A). In contrast, NZ-1 detected all the deletion mutants (EGFR/dN370, EGFR/dN375, and EGFR/dN380) (Fig. 2B). These results show that the N-terminus of the EMab-51 epitope exists between residues 375 and 380 (Fig. 2C).
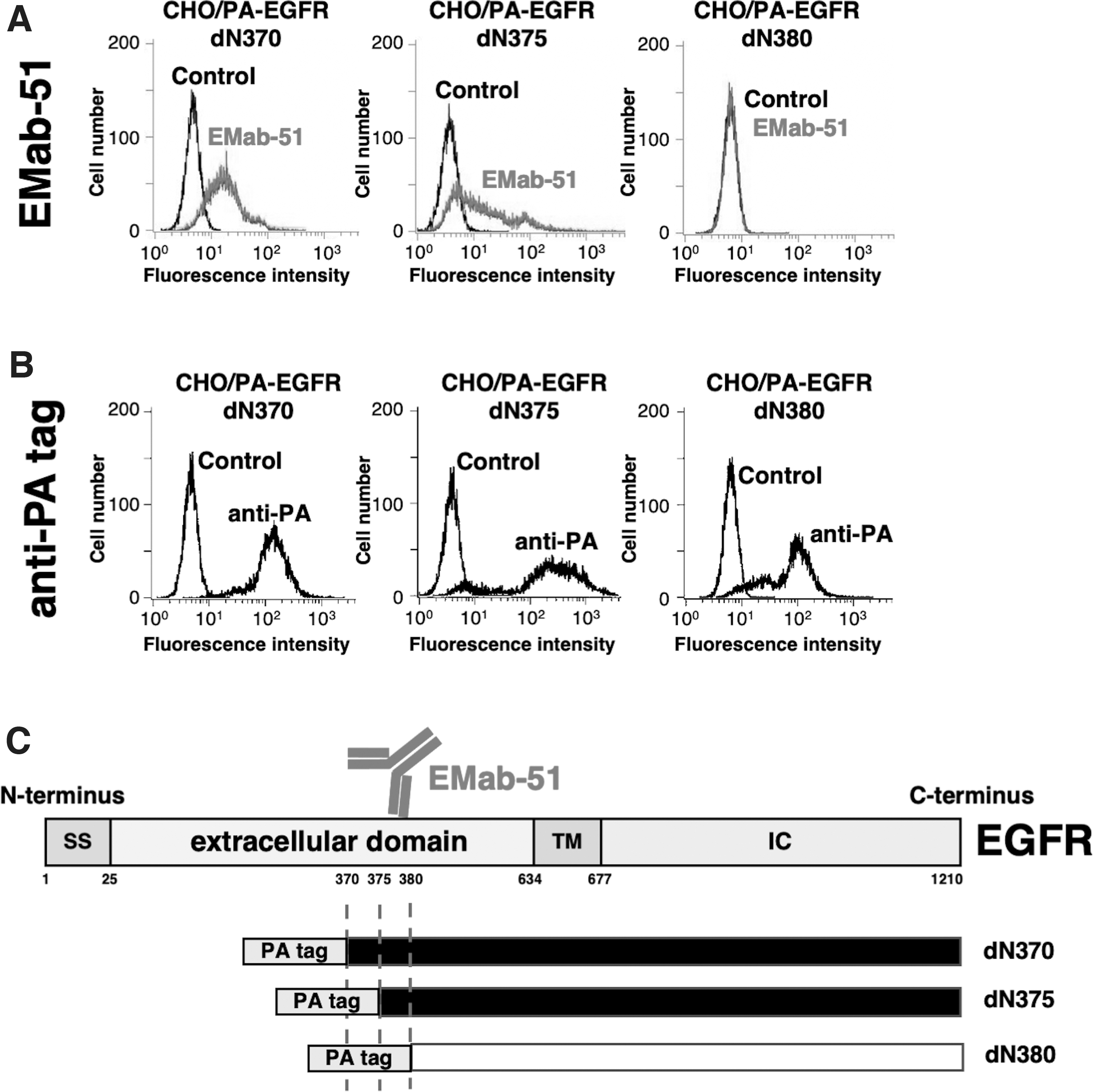
Epitope mapping using EGFR deletion mutants.
Determination of the EMab-51-recognized epitope using the REMAP method
To identify the amino acids composing the conformational epitope binding EMab-51, we used the REMAP method.(24) We constructed 14 EGFR/dN313 mutants, in which we inserted a RIEDL tag into the expected epitope region at each possible position of 375-AFRGDSFTHTPPLDP-389. The anti-RIEDL tag mAb (clone LpMab-7) recognized the five amino acid-long RIEDL tag.(24) For example, we produced Ala375_RIEDL_Phe376 (A375_R*_F376) by inserting the RIEDL sequence between Ala375 and Phe376 of EGFR/dN313.
Flow cytometry analysis showed that EMab-51 did not react with nine mutants, such as Arg377_RIEDL_Gly378 (R377_R*_G378), Gly378_RIEDL_Asp379 (G378_R*_D379), Asp379_RIEDL_Ser380 (D379_R*_S380), Ser380_RIEDL_Phe381 (S380_R*_F381), Phe381_RIEDL_Thr382 (F381_R*_T382), Thr382_RIEDL_His383 (T382_R*_H383), His383_RIEDL_Thr384 (H383_R*_T384), Thr384_RIEDL_Pro385 (T384_R*_P385), and Pro385_RIEDL_Pro386 (P385_R*_P386) although it strongly detected four mutants, such as Ala375_RIEDL_Phe376 (A375_R*_F376), Phe376_RIEDL_Arg377 (F376_R*_R377), Leu387_RIEDL_Asp388 (L387_R*_D388), and Asp388_RIEDL_Pro389 (D388_R*_P389), and weakly detected Pro386_RIEDL_Leu387 (P386_R*_L387) (Fig. 3A), indicating that EMab-51 might bind to EGFR through 10 amino acids (377-RGDSFTHTPP-386). The positive controls NZ-1 and LpMab-7 detected all 14 mutants (Fig. 3B, C). Thus, using the REMAP method, we determined that EMab-51 binds to the EGFR epitope 377-RGDSFTHTPP-386 (Fig. 4).

Epitope mapping using RIEDL tag insertion mutants of EGFR/dN313. RIEDL tag was inserted into EGFR/dN313. The RIEDL tag insertion mutants are as follows: Ala375_RIEDL_Phe376 (A375_R*_F376), Phe376_RIEDL_Arg377 (F376_R*_R377), Arg377_RIEDL_Gly378 (R377_R*_G378), Gly378_RIEDL_Asp379 (G378_R*_D379), Asp379_RIEDL_Ser380 (D379_R*_S380), Ser380_RIEDL_Phe381 (S380_R*_F381), Phe381_RIEDL_Thr382 (F381_R*_T382), Thr382_RIEDL_His383 (T382_R*_H383), His383_RIEDL_Thr384 (H383_R*_T384), Thr384_RIEDL_Pro385 (T384_R*_P385), Pro385_RIEDL_Pro386 (P385_R*_P386), Pro386_RIEDL_Leu387 (P386_R*_L387), Leu387_RIEDL_Asp388 (L387_R*_D388), and Asp388_RIEDL_Pro389 (D388_R*_P389). We analyzed the RIEDL tag insertion mutants using flow cytometry. We expressed the RIEDL tag insertion mutants in CHO-K1 cells and incubated them with
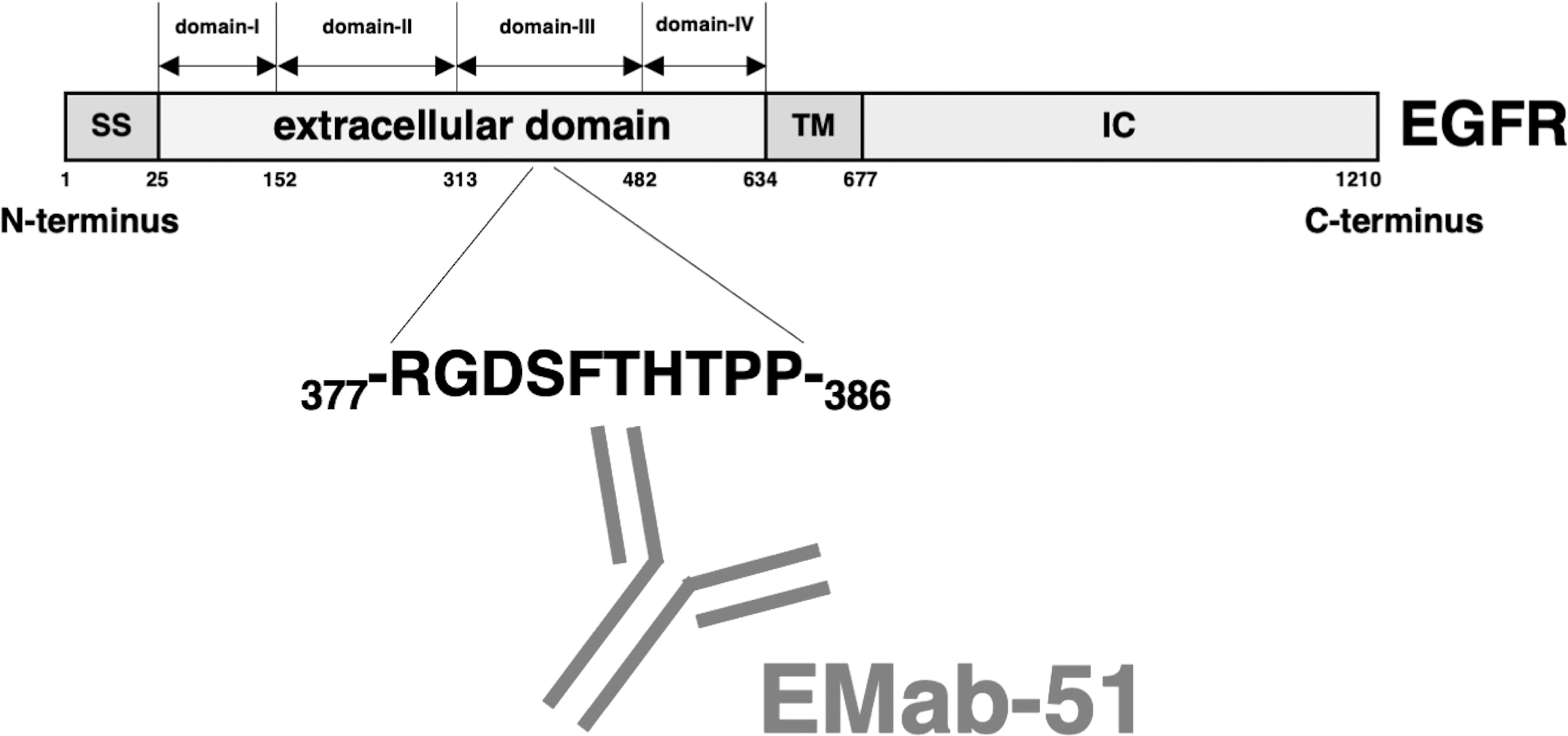
Schematic illustration of EMab-51 epitope. The EMab-51 epitope is 377-RGDSFTHTPP-386, which is located in the extracellular domain III of EGFR.
Discussion
mAb-binding epitope investigations often use alanine-scanning mutagenesis and peptide screening.(22,28–41) Although these methods are effective for identifying linear epitopes, they are inapplicable to conformational epitopes. By immunizing mice with EGFRec, we previously developed a novel anti-EGFR mAb (clone EMab-51) (23). EMab-51 has applications not only in flow cytometry and Western blotting but also in immunohistochemical analysis to detect EGFR in various cancers. Unfortunately, we could not determine the EMab-51-binding epitope using conventional epitope-mapping methods such as alanine scanning. This is because EMab-51 recognizes a conformational epitope and a single amino acid substitution may insufficiently disrupt the epitope conformation region to inhibit EMab-51 binding.
In this study, N-terminal deletion mutant analyses showed that the N-terminus of the EMab-51-binding epitope is located between EGFR residues 375 and 380 (Fig. 2). To identify the critical epitope amino acids, we employed the REMAP method.(24) The flow cytometry analysis showed that EMab-51 lost its reactivity to some RIEDL tag insertion mutants (Fig. 3). Thus, the RIEDL tag insertion may have partially disrupted the EGFR conformation and the binding of EMab-51 to EGFR. Using the REMAP method, we successfully determined that the EMab-51 epitope is 377-RGDSFTHTPP-386, which is located in the extracellular domain III of EGFR (Fig. 4). These amino acids are continuous, but may not form a linear structure. The epitope identification of EMab-51 will be helpful for the development of EGFR-targeting therapeutic antibodies in the future.
Footnotes
Authors' Contributions
R.N.,
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers JP21am0401013 (to Y.K.) and JP21am0101078 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) Grant Numbers 21K15523 (to T.A.), 21K07168 (to M.K.K.), 20K16322 (to