
Abstract
The C-C motif chemokine receptor 8 (CCR8) is highly expressed in regulatory T cells. CCR8 is also expressed in many cancers and is associated with those progression. The development of monoclonal antibodies (mAbs) for CCR8 leads to cancer immunotherapy and elucidation of unknown mechanisms of CCR8-dependent cancer progression. In this study, we have developed an anti-mouse CCR8 (mCCR8) mAb (clone C8Mab-3, rat IgG1, kappa) using the Cell-Based Immunization and Screening (CBIS) method. We revealed that C8Mab-3 and its recombinant antibody (recC8Mab-3) bind to mCCR8-overexpressed Chinese hamster ovary (CHO)-K1 cells (CHO/mCCR8), but not to the parental CHO-K1 cells, in flow cytometry. In addition, C8Mab-3 and recC8Mab-3 reacted to P388 (a mouse lymphocyte-like cell) and J774-1 (a mouse macrophage-like cell), which express endogenous mCCR8. C8Mab-3 also detected exogenous and endogenous mCCR8 using immunocytochemistry. These results suggest that C8Mab-3, developed using the CBIS method, is useful for immunocytochemistry against exogenous and endogenous mCCR8.
Introduction
The C-C motif chemokine receptor 8 (CCR8) is a family of the seven transmembrane domain G protein-coupled receptors (GPCRs) mainly expressed by regulatory T cells (Tregs).(1–5) Human CCR8 has four ligands: C-C motif chemokine ligand 1 (CCL1), CCL8, CCL16, and CCL18,(6) whereas only three of them, CCL1, CCL8, and CCL16, are expressed in mouse.(7–9) CCR8 is the only known receptor for CCL1,(5) whereas the other CCR8 ligands bind several chemokine receptors.(7–9) The CCR8+ Tregs are master drivers of immune regulation. In the mice model of encephalomyelitis, a T cell-mediated autoimmune disease of the central nervous system (CNS), the number of CCR8+ Tregs is reduced in the periphery. Moreover, CNS CCR8+ Tregs are potentiated by CCL1, which restricts the progression of the disease.(10)
Therapeutic monoclonal antibodies (mAbs) targeting immune checkpoints have changed the therapeutic strategy of tumors.(11,12) Tumors described as “hot” are signs of inflammation, particularly massive infiltration and enrichment with CD8+ effector T cells. The immune checkpoint inhibitors are effective in hot tumors. However, immune checkpoint inhibitors' response rate is still low owing to the lack of effector CD8+ T cell infiltration or massive accumulation of Tregs that suppress their activities, which is characterized as “cold tumors.”(13,14) Anti-mouse CCR8 (mCCR8) mAbs could be used to limit cancer growth in several cancer models.(15,16) Therefore, anti-CCR8 mAbs, mostly Tregs depleting antibodies, are currently developed for clinical use.
We have developed anti-GPCR mAbs including anti-mouse CCR3 mAb (clone C3Mab-2),(17) anti-human CCR9 mAb (clone C9Mab-1),(18) and anti-mCCR8 mAb (clone C8Mab-2)(19) using the Cell-Based Immunization and Screening (CBIS) method. This study developed another anti-mCCR8 mAb (clone C8Mab-3), and showed that C8Mab-3 recognizes both exogenous and endogenous mCCR8 in flow cytometry and immunofluorescence.
Materials and Methods
Cell lines and animals
P388 and J774-1 were obtained from the Cell Resource Center for Biomedical Research Institute of Development, Aging, and Cancer, Tohoku University (Miyagi, Japan). In addition, Chinese hamster ovary (CHO)-K1 and P3X63Ag8U.1 (P3U1) were obtained from the American Type Culture Collection (Manassas, VA).
Stable transfectants of CHO/mCCR8 were established in our previous study.(19) In brief, synthesized DNA (Eurofins Genomics KK, Tokyo, Japan) encoding mCCR8 (Accession No.: NM_007720.2) was subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). mCCR8 plasmid was transfected using a Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA). CHO/mCCR8 cells were sorted using a cell sorter (SH800; Sony Corp., Tokyo, Japan) and cultivated in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA).
P388, J774-1, CHO-K1, P3U1, and CHO/mCCR8 were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc., Kyoto, Japan), supplemented with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Inc.), 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 0.25 μg/mL of amphotericin B (Nacalai Tesque, Inc.). Cells were grown in a humidified incubator at 37°C with 5% CO2 and 95% air.
A female Sprague-Dawley (SD) rat (6 weeks old) was purchased from CLEA Japan (Tokyo, Japan). The animal was housed under specific pathogen-free conditions. All animal experiments were conducted following the relevant guidelines and regulations to minimize animal suffering and distress in the laboratory.
Animal experiments were approved by the animal care and use committee of Tohoku University (Permit No.: 2019NiA-001). The rat was monitored daily for health during the 4 full weeks duration of the experiment. A reduction of >25% of the total body weight was defined as a humane endpoint. The rat was euthanized by cervical dislocation, and death was verified by respiratory and cardiac arrest.
Hybridoma production
The CBIS method was used to develop mAbs against mCCR8.(20–26) One SD rat was immunized with CHO/mCCR8 cells (1 × 109) using intraperitoneal injection together with the Imject Alum (Thermo Fisher Scientific, Inc.). The procedure included three weekly immunization followed by a final booster intraperitoneal injection administered 2 days before harvesting spleen cells. Harvested spleen cells were subsequently fused with P3U1 cells using PEG1500 (Roche Diagnostics, Indianapolis, IN). The hybridomas were grown in RPMI 1640 medium supplemented with hypoxanthine, aminopterin, and thymidine for selection (Thermo Fisher Scientific, Inc.). Culture supernatants were screened using flow cytometry.
Production of the recombinant antibody
Variable (VH) and constant (CH) regions of heavy chain cDNAs of C8Mab-3 were subcloned into the pCAG-Neo vector along with variable (VL) and constant (CL) regions of light chain cDNAs of C8Mab-3 into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation) to produce recombinant C8Mab-3 (recC8Mab-3). C8Mab-3 vectors were transfected into ExpiCHO-S cells using the ExpiCHO Expression System (Thermo Fisher Scientific, Inc.). The resulting mAb (recC8Mab-3) was purified using Ab-Capcher ExTra (ProteNova Co., Ltd, Kagawa, Japan).
Flow cytometry
We harvested cells after brief exposure to 0.25% trypsin and 1 mM of ethylenediaminetetraacetic acid (Nacalai Tesque, Inc.), washed them with 0.1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS), treated them with primary mAbs (10 μg/mL) for 30 minutes at 4°C, and treated with Alexa Fluor 488-conjugated anti-rat IgG (1:2000; Cell Signaling Technology, Inc., Danvers, MA). In addition, we collected fluorescence data using the EC800 Cell Analyzer (Sony Corp.).
Determination of binding affinity by flow cytometry
We suspended CHO/mCCR8 cells in 100 μL of serially diluted anti-mCCR8 mAbs, and then in 100 μL of Alexa Fluor 488-conjugated anti-rat IgG (1:200; Cell Signaling Technology, Inc.). Fluorescence data were collected using the EC800 Cell Analyzer. The dissociation constant (KD) was calculated by fitting binding isotherms to built-in one-site binding models in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA).
Immunocytochemistry
CHO-K1, CHO/mCCR8, P388, and J774-1 cells were cultured on acid-wash coverslips. They were treated with 4% paraformaldehyde (in PBS) for 10 minutes and in the additional 50 mM of NH4Cl (in PBS containing 0.2 mM of Ca2+ and 2 mM of Mg2+ [PBSc/m]) for 10 minutes. Subsequently, they were incubated with a blocking buffer (PBSc/m containing 0.5% BSA) for 30 minutes, C8Mab-3 or recC8Mab-3 (10 μg/mL; in a blocking buffer) for 1 hour, and Alexa Fluor 488-conjugated anti-rat IgG (1:400; in a blocking buffer) for 45 minutes. 4′,6-Diamidino-2-phenylindole (DAPI; Thermo Fisher Scientific, Inc.) was used for the cell nuclei staining. The fluorescence images were acquired on a digital microscope BZ-X800 (Keyence, Osaka, Japan) using a 40 × objective.
Results
Establishment of anti-mCCR8 mAbs
We employed the CBIS method to develop anti-mCCR8 mAbs. In the CBIS method, we used stable transfectant immunization and flow cytometry-mediated hybridoma screening.(20–26) In this study, a rat was immunized with CHO/mCCR8 cells. Hybridomas were seeded into 96-well plates, and CHO/mCCR8-positive and CHO-K1-negative wells were selected. After limiting dilution, C8Mab-3 (rat IgG1, kappa) was finally established. We further produced its recombinant antibody (recC8Mab-3).
Flow cytometry
We conducted flow cytometry using C8Mab-3 and recC8Mab-3 against CHO/mCCR8 and CHO-K1. Both C8Mab-3 and recC8Mab-3 reacted with CHO/mCCR8, but not with CHO-K1 (Fig. 1). In addition, both C8Mab-3 and recC8Mab-3 also recognized P388 and J774-1, which express endogenous mCCR8 in a dose-dependent manner (Fig. 1).
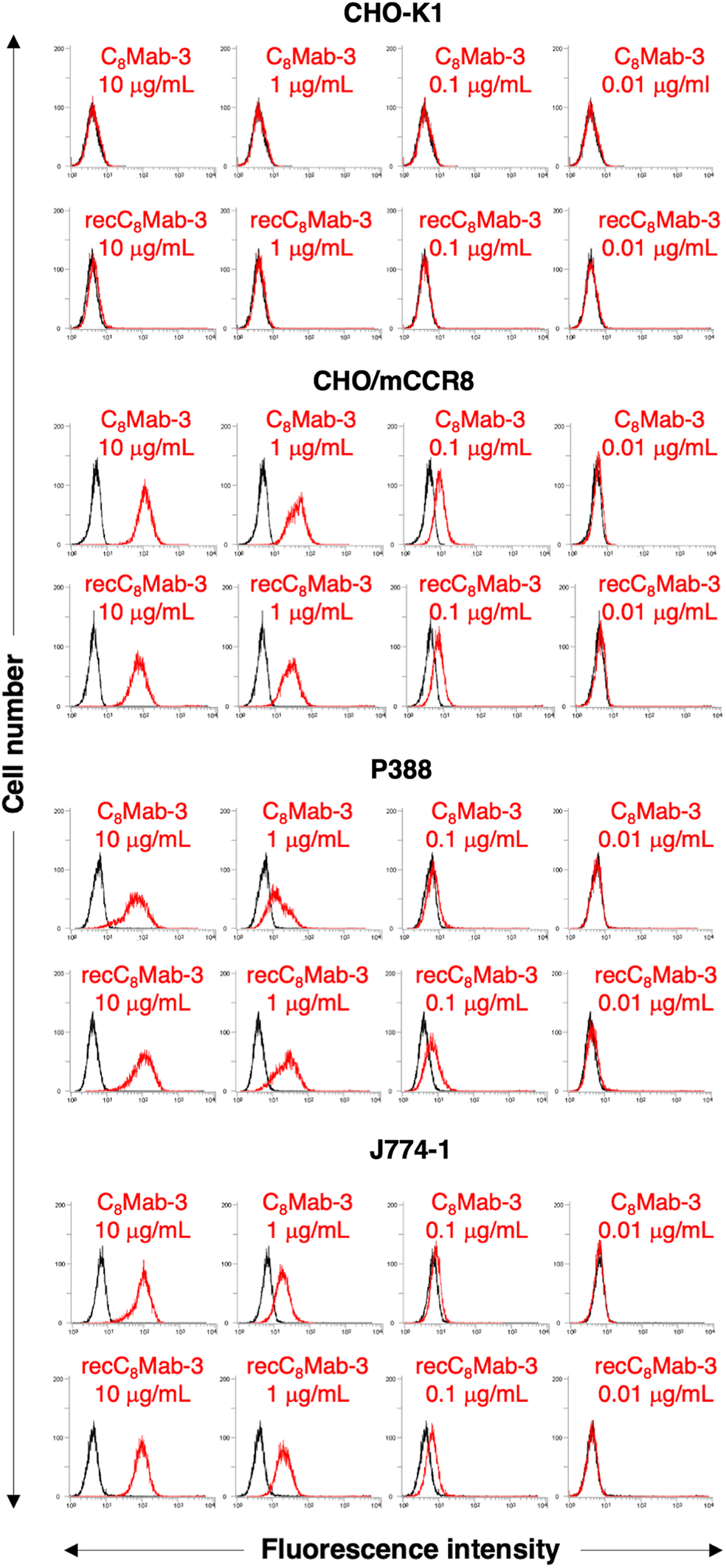
Flow cytometry using anti-mCCR8 mAbs, C8Mab-3 and recC8Mab-3. CHO-K1, CHO/mCCR8, P388, and J774-1 were treated with 0.01–10 μg/mL C8Mab-3 or recC8Mab-3, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG; black line, negative control. CHO, Chinese hamster ovary; CCR8, CC motif chemokine receptor 8; mAbs, monoclonal antibodies.
Determination of the binding affinity of C8Mab-3 and recC8Mab-3
The binding affinity of C8Mab-3 and recC8Mab-3 with CHO/mCCR8 was evaluated using flow cytometry. The KD of C8Mab-3 and recC8Mab-3 for CHO/mCCR8 was 1.1 × 10−7 and 1.7 × 10−7 M, respectively (Supplementary Fig. S1), indicating that C8Mab-3 possesses a low affinity for CHO/mCCR8 cells.
Immunocytochemical analysis using C8Mab-3 and recC8Mab-3
We explored the availability of C8Mab-3 and recC8Mab-3 in immunocytochemical analysis. First, C8Mab-3 (10 μg/mL) and recC8Mab-3 (10 μg/mL), but not buffer control, bound to CHO/mCCR8 (Fig. 2), whereas both antibodies did not bind to CHO-K1 (Fig. 2), suggesting that C8Mab-3 and recC8Mab-3 specifically detect exogenous mCCR8. Next, C8Mab-3 and recC8Mab-3, but not buffer control, are also bound to P388 and J774-1 cells, suggesting that both antibodies recognize endogenous mCCR8 (Fig. 2). These results show that C8Mab-3 and recC8Mab-3 are available in immunocytochemical analysis against exogenous and endogenous mCCR8.
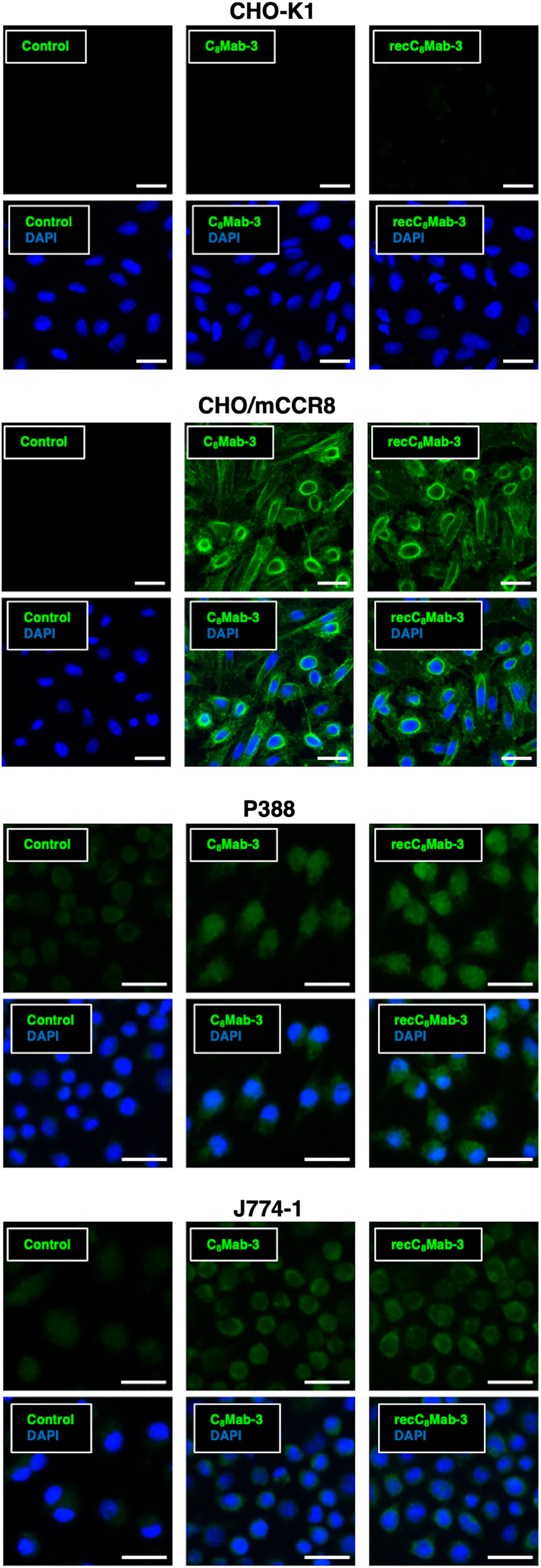
Immunocytochemistry using C8Mab-3 and recC8Mab-3. CHO-K1, CHO/mCCR8, P388, and J774-1 were treated with buffer control, 10 μg/mL C8Mab-3, or recC8Mab-3, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG. DAPI was used for nuclear staining. Scale bars, 20 μm. DAPI, 4′,6-diamidino-2-phenylindole.
Discussion
We developed a novel mAb for mCCR8, C8Mab-3, using the CBIS method. Both C8Mab-3 and recC8Mab-3 bind to mCCR8 with low affinity (Supplementary Fig. S1), but they specifically recognize exogenous and endogenous mCCR8 in both flow cytometry (Fig. 1) and immunocytochemistry (Fig. 2). The confirmation to other applications, including Western blotting, immunoprecipitation, and immunohistochemistry, is required for basic study in the future.
The secretion of CCL1, a ligand of CCR8, by CD11b+CD14+ myeloid cells is involved in Treg cell infiltration.(27) Increased expression of CCR8 is observed in Treg cells and natural killer T cells, especially in cancer patients.(28) Upon binding of CCL1 to CCR8, the expression of FoxP3 is elevated through the STAT3 pathway, and the activated CCR8+ Treg cells strongly suppress antitumor immunity by secretion of IL-10 and granzyme B.(10) The overall survival of breast cancer patients with high infiltration of CCR8+FoxP3+ Tregs is significantly shorter than that of patients with low infiltration.(27) Therefore, therapies that target CCL1–CCR8 molecules are desired in the clinic. Currently, we have developed three mAbs for mCCR8 (C8Mab-1 to 3). The determination of their epitopes and neutralizing activity for CCL1–CCR8 interaction is required for in vivo experiments.
Tregs express several cell surface markers, including CD25, CTLA-4, PD-1, ICOS, GITR, OX40, CD15s, CCR4, and CCR8, and these markers can be used to deplete Tregs. Tregs are also indispensable for preventing autoimmunity. The removal of Tregs improves antitumor immunity and may also trigger autoimmunity.(29) Therefore, we should specifically deplete Tregs in tumors for cancer immunotherapy. Since CCR8 expression is increased in tumor-infiltrated Tregs,(28) CCR8 is one of the promising targets depleting Tregs selectively in tumors.
In our previous studies, we changed the isotype of mAbs into mouse IgG2a to retain antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC), which caused high antitumor activities in mouse xenograft models.(30–32) Since the subclass of C8Mab-3 is rat IgG1, it does not possess ADCC and CDC. Therefore, in further studies, the subclass of C8Mab-3 will be converted into mouse IgG2a to assess the effect of depletion of CCR8+ Treg cells in cancer immunotherapy.
In summary, we developed a novel mAb for mCCR8, C8Mab-3, in this study. C8Mab-3 and recC8Mab-3 apply to flow cytometry and immunofluorescence. These mAbs would be valuable for elucidating the roles of CCR8 in cancer.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.