
Abstract
During the past decades, tremendous advances have occurred in manufacturing recombinant therapeutic proteins in Chinese hamster ovary (CHO) cells. Nevertheless, the production of stable high-producing cell lines has remained a major obstacle in the development process of the CHO cell line. It has been shown that genomic regulatory elements can promote cell line development efficiency by improving transgenes' productivity and stability. Such elements include insulators, ubiquitous chromatin opening elements, scaffold/matrix attachment regions, and antirepressors. In addition, tDNA elements are shown to act as insulators in mammalian cells. This study examines the effect of the tDNA insulator on stable expression of a vascular endothelial growth factor receptor-Fc fusion protein.
Introduction
Today, recombinant proteins play vital roles in biomedical research as well as therapeutics and diagnostics settings. Recombinant proteins of clinical significance, including growth factors, hormones, clotting factors, Fc-fusion proteins, and monoclonal antibodies, have provided novel therapeutic options for human diseases. 1 In this regard, antiangiogenic therapeutic recombinant proteins have been successfully employed to treat ocular diseases such as diabetic retinopathy and age-related macular degeneration.2,3 Examples of these matters include Fc-fusion proteins and vascular endothelial growth factor (VEGF)-neutralizing monoclonal antibodies. 4
Fc-based fusion proteins include an immunoglobulin Fc domain attached to a peptide or protein-based fusion partner. 5 The fusion moiety is usually responsible for the therapeutic effect, and the Fc domain improves the plasma half-life. 6
Mammalian cell expression systems (e.g., Chinese hamster ovary [CHO] cells) have been extensively employed to produce recombinant therapeutic proteins. 7 Productivity and stability of a recombinant protein producing CHO cell line are highly important in industrial manufacturing of therapeutic proteins. 8 As a result, researchers have examined numerous approaches to improve recombinant protein expression in CHO cells. 9 Several studies have focused on vector engineering strategies considering the significance of expression vector design in recombinant protein expression. 10 In this regard, scaffold/matrix attachment regions (S/MARs), ubiquitous chromatin opening elements, antirepressors, and insulators have been employed.11,12
Insulators are regulatory elements that can exert two main activities: the enhancer blocking and position effects blocking activities. 13 Researchers have evaluated several insulators to enhance the expression and stability of the recombinant protein in mammalian cells using plasmid and viral-based vectors. The best-studied example includes the cHS4 insulator; however, this element has only shown little efficiency in CHO cells. 14
Identification and characterization of novel insulator elements are an attractive field of study. In this respect, Raab et al. 15 showed that human tRNA genes can act as insulators with both enhancer blocking and barrier activities in mammalian cells. Also, Lee et al. reported a tDNA insulator consisting of two copies of tRNA genes that showed barrier activity in the centrochromatin region of a human artificial chromosome (HAC) vector. 16
We have previously shown the positive effects of human Beta-Interferon Scaffold Attachment Region on stable expression of vascular endothelial growth factor receptor-Fc (VEGFR-Fc) fusion protein in CHO-K1 cells. 17 The present study aimed to examine the tDNA insulator impacts on the stable expression of VEFGR-Fc fusion protein to further deduce optimal vector systems for the expression this fusion protein.
Materials and Methods
Vector design
The pFU-puro expression vector containing VEGFR-Fc fusion protein coding sequence under human cytomegalovirus (CMV) promoter was described previously. 17 Also, the pTD-EGFP expression vector, 18 which contains 500 bp tDNA elements at the 5′- and 3′-flanking sites of the EGFP coding sequence, was employed to construct the VEGR-Fc expression vector harboring the insulator element. The 1430 bp VEGFR-Fc coding sequence was subcloned from pFU-puro into the XhoI-NotI restriction enzyme site of the pTD-EGFP vector to replace the EGFP coding sequence to obtain the pTD-FU-puro expression vector.
Cell culture and transfection
An adherent CHO-K1 (ATCC CCL-61) cell line was employed as a host cell. Cells were cultured in DMEM/F12 medium complemented with
Development of stable cell pool
CHO-K1 cells were transfected with pTD-FU-puro and pFU-puro expression vectors. About 48 hours after the transfection, cell dilution was done in a fresh medium in a 1:10 ratio, and the cells were subjected to antibiotic selection in a medium containing puromycin at 5 μg/mL concentration for 2 weeks. Surviving cells were kept in the selected medium for another 3–5 days and then were expanded in T25 flasks for further analysis.
Evaluation of VEGF-Fc expression
VEGFR-Fc expression was evaluated by culturing pTD-FU-puro and pFU-puro pools for 72 hours in six-well plates. Cell culture supernatants were then collected and diluted in phosphate-buffered saline (PBS) in 1:5 and 1:2.5 ratios. In this study, a human IgG1 antibody was used as the standard with concentrations at 0, 6.25, 12.5, 25, 50, 100, and 200 ng/mL. The standards and diluted supernatants were coated in 96-well microtiter plates for 2 hours. After three washing steps and 1 hour blocking in 3% skim milk solution, the process was followed by adding an horseradish peroxidase-conjugated antihuman IgG1 Fc antibody (diluted 1:10,000 in PBS with 1% bovine serum albumin) to the wells and incubating it for 1 hour.
The wells were then rinsed. After adding the tetramethylbenzidine (TMB) substrate (100 μL), the product was incubated for 15 minutes. Lastly, the reaction was stopped by adding 1 M HCl solution, and the optical density was calculated at 450 nm. In this research, supernatants of CHO-K1 cells were used as a negative control.
Quantitative real-time reverse transcription PCR
Total RNA was extracted from pTD-FU-puro and pFU-puro stable cell pools using a total RNA Isolation Kit (Favorgen Biotech). The extracted RNA was then treated with RNase-free DNase (Thermofisher Scientific, USA), and cDNA synthesis was conducted using a first-strand cDNA Synthesis Kit (Vivantis, Malaysia). Quantitative real-time reverse transcription PCR (qRT-PCR) was used in a Real-Time PCR system (Applied Biosystems, USA) to measure VEGFR-Fc mRNA expression based on these parameters: 20 seconds at 95°C, 40 cycles of 15 seconds at 95°C, and 30 seconds at 60°C by SYBR Green PCR Master Mix (Ampliqon, Denmark). The b-actin housekeeping gene served as an internal control.
The primers included 5′- TCCTCAGTAATTGCATCAG-3′ (reverse) and 5′-AGCCAGAATTGACATCTTC-3′ (forward) for Fc region; 5′-AGGTGTGGTGCCAGATCTTC-3′ (reverse) and 5′ ACTCCTACGTGGGTGACGAG-3′ (forward) for b-actin.
Single-cell cloning
The serial dilution cloning approach was applied to perform single-cell cloning. Cells were diluted to 5 cell/mL concentration in Ham's F-12 Nutrient Mixture with 15% FBS, and 100 μL of the obtained cell suspensions was seeded in 96-well plates. After 7 days, single-cell clones were recognized and incubated for another week. The clones were expanded in 12-well plates and subsequently cultured in T25 flasks.
This study was approved by ethical committee of Jiroft University of Medical Sciences, Ethical ID is IR.JMU.REC.1395.009.
Results
Expression vector construction
Restriction digestion analysis was used to confirm the VEGFR-Fc fragment cloning in XhoI-NotI sites. Observation of a 1430 bp fragment indicated successful cloning of the fusion protein in pTD-FU-puro and pFU-puro vectors (Fig. 1b). Figure 1c shows the schematic map of the expression vectors used in this study.

Development of cell pools
FU and TD-FU pools were generated after selecting pFU-puro and pTD-FU-puro transfectants in a puromycin-containing medium. The presence of VEGFR-Fc in stable cell pools was evaluated by PCR analysis on genomic DNA of pools using CMV For and Poly A Rev primers. As indicated in Figure 2b, a 2200 bp fragment corresponding to CMV-VEGFR-Fc-Poly A was observed in both pools.
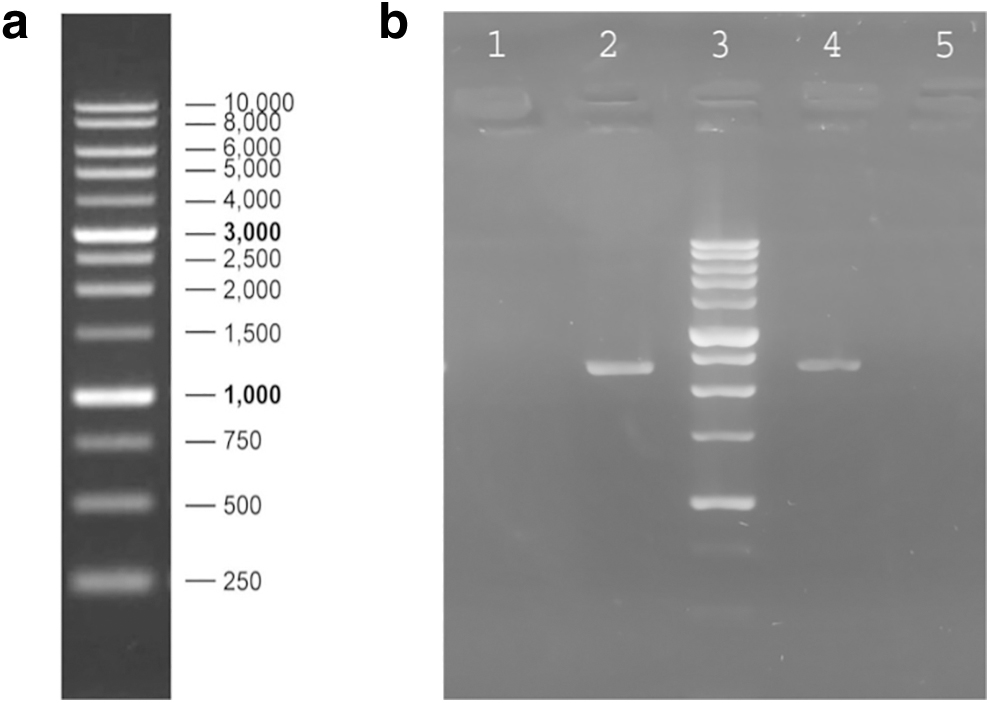
PCR analysis on genomic DNA of FU and TD-FU stable cell pools using CMV For and Poly A Rev primer.
VEGFR-Fc expression analysis in cell pools
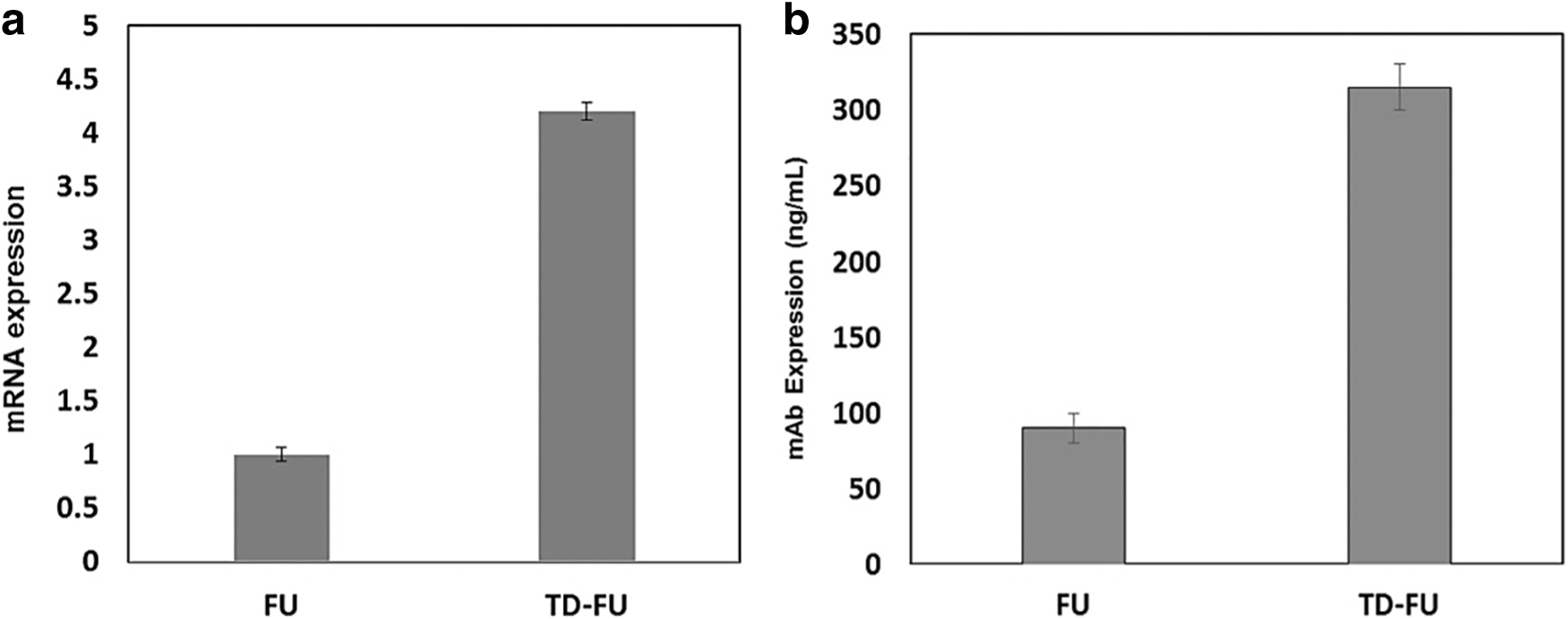
VEGFR-Fc expression in cell pools was evaluated at mRNA and protein levels. In this study, the qRT-PCR analysis indicated up to 4.2-fold enhancement in mRNA expression in pools generated using the pTD-FU-puro vector (Fig. 3a). Enzyme-linked immunosorbent assay (ELISA) analysis on cell culture supernatants of stable cell pools during 3 days of culture also indicated a 3.5-fold enhancement in secretory VEGFR-Fc expression (Fig. 3b).
VEGFR-Fc expression analysis in single-cell clones
Although cell pools are a heterogeneous mixture of different cells, clonal cells provide a homogeneous cell population. Therefore, VEGFR-Fc expression was analyzed in single-cell clones using ELISA. The mean expression of the fusion protein in pTD-FU-puro-derived clones was 3.39-fold higher (67 ng/mL vs. 227.64 ng/mL) than in pFU-puro clones. In addition, the two highest producing clones belonged to the TD-FU population with titers of 330 and 288 ng/mL compared with 100 and 85 ng/mL in FU high producing clones (Fig. 4).

Analysis of antibody production in five single-cell clones derived from stable a CHO cell pool transfected with pFU-puro and pTD-FU-puro vector by the ELISA. Error bars demonstrate standard deviation.
Discussion
Monoclonal antibodies and their derivatives have revolutionized modern medicine in both diagnostic and therapeutic settings. Fc-fusion proteins increase the half-life of the therapeutic proteins.6,19 VEGFR-Fc fusions were considered promising antiangiogenesis therapeutics, acting as VEGF neutralizing agents. 4 Effective expression of Fc-fusions depends on mammalian expression systems, like CHO cells. 20 Such systems suffer from low productivity despite their numerous advantages. 21
The effects of insulator elements on heterologous gene expression in mammalian cells have been examined in different studies. The positive effects of Ccnb1ip1 22 and gamma-satellite 23 insulators on monoclonal antibody expression in CHO cells have been reported. Increased expression level of an anti-CD52 monoclonal antibody in CHO cells has also been reported using tDNA insulator. 18 In line with previous reports, this study evaluated the effect of tDNA insulator on VEGFR-Fc stable expression in CHO cells.
After gene cloning, stable cell pools containing tDNA and control vectors were developed. Analysis of VEGFR-Fc expression in mRNA and protein levels indicated up to 4.2- and 3.5-fold enhancement, respectively. Although analysis of stable pools (i.e., a mixture of clonal cell populations) gives an initial understanding of the heterologous gene expression, evaluation of single clonal cells can be much more informative and yields more reproducible results. Therefore, in the next step, single-cell clones were separated from cell pools and were subjected to productivity analysis. This evaluation indicated up to fourfold increase in mean VEGFR-Fc expression in clones derived from tDNA containing pool.
Our results are consistent with those obtained by Naderi et al., 18 who reported 7.9- and 6.1-fold increases in mAb titer using tDNA insulator in cell pools and clonal cells, respectively. The tDNA insulator has also increased EGFP expression from a human artificial vector (HAC) in CHO and HeLa cells. 16 This study confirms that this element could be beneficial for the expression of both intracellular and secretory proteins.
In another study, Naghneh et al. 17 evaluated the VEGFR-Fc fusion protein expression using human beta-interferon S/MAR, which resulted in a 2.2-fold enhancement in mAb expression in cell pools and clonal cells. The results obtained in this study are comparable with those reported before, indicating that both tDNA insulator and beta-interferon S/MAR positively impact VEGFR-Fc expression in CHO cells.
Insulators protect a given gene from the negative effects of its surrounding chromatin. S/MARs protect genes by creating independent loops in chromatin and activating transcription. Increased expression of EGFP from lentiviral and plasmid-based vectors has been reported using combination of HS4 insulator and SAR element (HS4-SAR).24,25 Whether using tDNA insulator in addition to human beta-interferon S/MARs can elevate the expression of VEGFR-Fc fusion protein even further can be examined in future studies.
Footnotes
Acknowledgments
The authors thank School of Advanced Technologies in Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran, and Department of Medical Genetics, Semnan University of Medical Sciences, Semnan, Iran.
Authors' Contributions
M.S. and E.P. came with the idea of this study; A.R.., M.M., and S.H.T. did the study experiments and drafted the article; F.P. and R.H. revised the article; all of the authors read the final article and approved it for submission.
Author Disclosure Statement
No competing financial interests exist..
Funding Information
No funding was received for this article.