
Abstract
The acquisition of resistance to amoxicillin, tetracycline, and enrofloxacin by Escherichia coli MG 1655 was examined by exposing growing cells to constant or stepwise increasing concentrations of these compounds. The minimal inhibitory concentration (MIC) of E. coli for amoxicillin increased from 4–8 to 32 μg/ml after growth in the presence of 1.25 or 2.5 μg/ml. By stepwise increasing the exposure, an MIC of 512 μg/ml was reached. This high MIC was maintained after removal of the antibiotics, whereas the lesser increase after exposure to low levels was reversed, indicating that the high MIC was due to a genetic change, but the lower one to phenotypic adaptation only. The MIC for tetracycline increased from 2 μg/ml to maximally 32 μg/ml. The MIC decreased to control levels in the absence of tetracycline, so no genetic changes seem to have occurred. The MIC for enrofloxacin increased from 0.25 μg/ml to maximally 512 μg/ml depending on the concentration during growth. These data mostly support the “radical-based” theory that bactericidal antibiotics induce a common mechanism that contributes to cell killing. Our findings indicate that exposure to low levels of antibiotics causes an increase in MIC above the concentration that the cells were exposed to. The implication is that exposure to low levels of antibiotics should be prevented as much as possible, because this causes resistance far more than high concentrations that inhibit growth or kill the cell and thus prevent acquisition of resistance.
Introduction
Resistance is not only selected for by therapeutic use on humans. The extensive use of antibiotics in the agricultural sector has turned farms into sources of resistant microbes.5,9,10,43 Even in soil a considerable increase in resistance genes has been demonstrated. 20 The resistance selected for in the agricultural setting may be a direct threat as zoonotic agents become resistant or it can be indirect as it is eventually transferred from animal commensals to human pathogens. 5 In response, attempts are being made to reduce usage. For example, the use of antibiotics as growth promoters is prohibited in Europe and many other parts of the world. Often the recommendation is made to abandon the use of specific classes that are important for human therapy, such as fluoroquinolones for agricultural use. 31 A sound policy for reduction of usage can only be set if the effect of dosage and length of exposure are known. Therefore, this study explores the acquisition of resistance due to different antibiotic regimes and the subsequent reduction during growth in the absence of the antibiotics. Special attention is given to the effect of low concentrations, as these often occur in the agricultural sector because of subtherapeutic use and carryover of antibiotics after production medicinated feed to a following batch of regular feed.
Antibiotic resistance can be acquired through three main mechanisms: (1) transfer of resistance genes from resistant to susceptible microorganisms; (2) genetic adaptation, for example, changing the drug target; and (3) phenotypic adaptation, primarily increase of expression, of existing cellular machinery, such as efflux pumps. The latter two mechanisms, which yield resistance de novo, may be induced by a single mechanism in the case of bactericidal antibiotics, according to the “radical-based mechanism” theory.21,22 This theory proposes that even though the various bactericidal antibiotics have different targets, they all induce hyperactivation of the electron transport chain, which in the end leads to the formation of hydroxyl radicals. These radicals damage DNA, proteins, and lipids, contributing to cell death.
If the radical-based mechanism theory is correct, the build-up of resistance should follow a similar pattern for bactericidal antibiotics, such as beta-lactams or fluoroquinolones, but a different one in the case of their bacteriostatic equivalent, for example, tetracycline. Hence, the effects of exposure are likely to vary between different types of antibiotics. Resistance of Escherichia coli to multiple antibiotics is controlled by the multiple antibiotic resistance gene, which acts in response to a variety of chemicals that contain aromatic rings. 2 Antibiotic resistance carries a metabolic cost4,29 and different mechanisms of resistance can be expected to have varying costs.
In this study, we describe the de novo acquisition of resistance by E. coli against three antibiotics, the bactericidals amoxicillin and enrofloxacin and the bacteriostatic tetracycline. The metabolic costs of the resistance in each of these cases are compared. All data considered together suggest that the greatest risk of acquired resistance results from exposure to subminimal inhibitory concentrations (sub-MICs) of antibiotics.
Materials and Methods
Bacterial strains, growth media, and growth conditions
Throughout this study, the often used laboratory strain E. coli MG1655 was utilized. Cells were grown at 37°C in a defined minimal mineral medium containing 55 mM glucose 11 with a pH of 7.0 and a buffer of 15.6 g/L Na2H2PO4. Media were autoclaved for 20 minutes at 120°C, with the exception of glucose (10 minutes, 110°C). Precultures for inoculation of 96-well plates and batch fermentors were grown in shake-flasks continuously shaken at 200 rpm.
Adaptation to antibiotics by prolonged growth at increasing levels
Wild-type E. coli was grown overnight in the presence of sublethal concentrations of amoxicillin, initially 1.25 and 2.5 μg/ml, tetracycline, 0.5 and 1.0 μg/ml, or enrofloxacin, 0.125 and 0.25 μg/ml. When normal or almost normal growth occurred, a small aliquot of the culture, calculated to yield an initial OD600 of 0.1, was used to start two more incubations, one at the same level of antibiotic and the other at double concentration. This way the concentration of the antibiotic was increased in twofold increments each time that the cells were able to grow at the control rate, or at least at >75% of that rate. After 15 daily cycles of exposure to fixed or, when possible, increasing amounts of antibiotic, cells were inoculated in fresh medium without antibiotic every day for another 15 days.
MIC readings
MIC values were determined as previously described. 38 Duplicate serial dilutions, ranging from 0 to 4,096 μg/ml of each antibiotic, were made in 96-well plates using Evans medium. Tetracycline and amoxicillin stock solutions of 10 mg/ml were freshly made for each experiment in 1 N HCl. Enrofloxacin (10 mg/ml) was dissolved in 0.1 N HCl and, after dissolution, brought to pH 10. All stock solutions were 0.2-μm filter-sterilized and stored at 4°C prior to use. Tetracycline stock solutions were wrapped in tinfoil to prevent exposure to light. E. coli was inoculated into each well to an approximate starting OD600 of 0.05. Growth was followed over time in a microtiter plate reader, measuring OD600 every 10 minutes, with shaking in between. For the 96-well measurements, a Thermo Scientific Multiskan FC with SkanIt software or a BMG Fluorostar with Optima MARS software was used. The MIC was defined as the lowest concentration of antibiotic that reduced the growth to an OD of 0.2 or less after 24 hours. The MIC measurements were performed daily in the beginning of an experiment and later at regular intervals and when growth at a higher concentration occurred.
Physiological characterization of adapted strains
For determination of growth rate and specific metabolic rates, cells were grown in batch fermentors. Cells were inoculated from an overnight culture to an OD600 of 0.1. Cultures were well stirred and well aerated (1 v/v per minute) in 0.5 L working volume. pH was measured continuously and kept at pH 7.0 by adding 1 M NaOH. Cell growth was followed by measuring OD600, cell counts, and biomass (dry weight). Samples for measurement of glucose and metabolic end-product concentrations were analyzed by high-performance liquid chromatography (HPLC). Using this setup, wild-type E. coli MG1655 and the adapted strains obtained as described above were grown in the absence or presence of various levels of antibiotics. The adapted strains were also grown in the presence of a high concentration of the antibiotic to which they had been adapted.
In one experiment, cells were grown in a batch fermentor for 4 days, in medium with 10 μg/ml amoxicillin. Every day, cells were diluted in fresh medium to OD600 0.1 and samples were taken for HPLC analysis.
HPLC characterization was used to determine concentrations of fermentation products and remaining glucose. For this, 1 ml of culture was mixed with 100 μl of 35% (v/v) perchloric acid. After neutralization with 55 μl of 7 M KOH, samples were centrifuged. Subsequently, the supernatant was filtered and analyzed by HPLC using RI detection (Phenomenex type Rezex Organic Acid column; eluent, 7.2 mM H2SO4 at 45°C) and Azur HPLC analysis software (Datalys, France).
Results
The MIC of E. coli for amoxicillin increased rapidly upon exposure to sub-MICs. Cells growing for 15 days with daily transfers in the presence of 1.25 or 2.5 μg/ml reached a maximum MIC of 32 μg/ml (Fig. 1). There was no systematic difference between cultures grown at 1.25 or 2.5 μg/ml. The MIC of the control varied between 4 and 8 μg/ml. When the concentration was increased in twofold increments every time the cells grew at a certain concentration at rates equal to or at least 75% of the control rate, a maximum MIC of 512 μg/ml was reached. When the cultures were subsequently grown in antibiotic-free medium, the MIC of E. coli previously grown at 1.25 or 2.5 μg/ml amoxicillin decreased within 2 days to that of the control. In contrast, the MIC of the cells exposed to incrementally increasing levels remained at 256 μg/ml throughout the following 2 weeks, indicating the long-lasting nature of the resistance.

Increase of the MIC during growth of Escherichia coli at various amoxicillin regimes. MIC, minimal inhibitory concentration.
Resistance against tetracycline increased by maximally 3 or 4 twofold increments from 2–4 to 32 μg/ml (Fig. 2). The MIC of the control seemed to increase slightly as well, possibly as an effect of growth in minimal medium. Cells exposed to 1 μg/ml tetracycline had an MIC that exceeded that of cultures exposed to 0.5 μg/ml by a factor of 2. Steadily increasing the concentration whenever the cells grew well yielded an MIC that was only twice as high as that of cells grown at 1 μg/ml constantly. After growth in the absence of tetracycline, the MICs of all cultures returned to the control value.

Increase of the MIC during growth of E. coli at various tetracycline regimes.
Resistance to enrofloxacin built up rapidly, eventually by 3 orders of magnitude (Fig. 3). Exposure to initially low concentrations was also in this case necessary. The levels of resistance reached varied between and within experiments, but the MICs remained approximately at the final levels when growing subsequently in the absence of enrofloxacin. Exposure to levels below the susceptible MIC resulted in a jump of the MIC by a factor of up to 100, which remained after the exposure was ended. This outcome can be explained by assuming that different genetic mutations have occurred in separate experiments. The higher levels of resistance could be the result of several mutations having an additional affect.
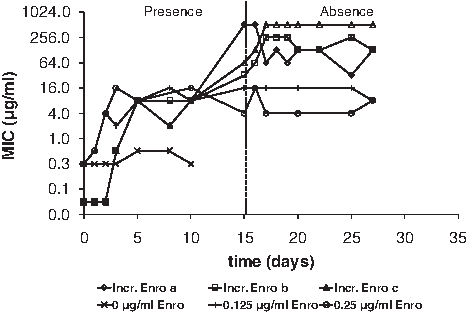
Increase of the MIC during growth of E. coli at various enrofloxacin regimes.
The growth rate of E. coli in the presence and absence of amoxicillin (Fig. 4) was the same under all conditions tested. The specific glucose consumption (q) of the cells adapted to low concentrations of amoxicillin was almost double at 2 μg/ml, but those adapted to high levels had a normal consumption rate at 64 μg/ml. This indicates that in the first case the adaptation required energy, but the adaptation in the latter case is of a different nature, demanding less maintenance energy. End products were measured as well, but the relative distribution did not change, and therefore, the results are not presented.
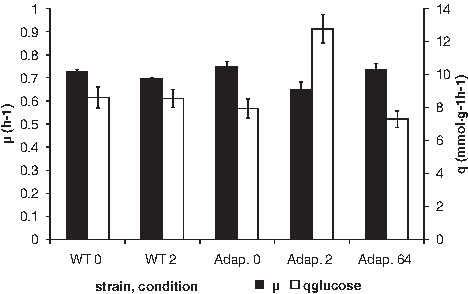
Growth rate and specific glucose consumption rates of E. coli in the presence or absence of amoxicillin. WT0: wild type, no antibiotics; WT2: wild-type strain growing in the presence of 2 μg/ml amoxicillin; Adap. 0: strain adapted to grow at 64 μg/ml amoxicillin growing in the absence of antibiotics; Adap. 2: strain adapted to grow at 64 μg/ml amoxicillin growing in the presence of 2 μg/ml; Adap. 64: strain adapted to grow at 64 μg/ml amoxicillin growing in the presence of 64 μg/ml.
The concept of a transient character of adaptation and the utilization of energy for that purpose is in agreement with the finding that, upon exposure to initially inhibiting concentrations of amoxicillin, growth at first stops, but resumes afterward (Fig. 5). When a normally growing culture of E. coli was exposed to 10 μg/ml, growth was completely inhibited, but glucose was still consumed at about two thirds of the normal rate. After 24 hours, growth resumed, albeit at a lower rate, which recovered to the control rate after another 48 hours. By that time, the specific glucose consumption had almost doubled, and thus, the yield was halved. These observations can be explained by assuming that the growth in the presence of these concentrations of amoxicillin is possible only at the expense of considerable energy.
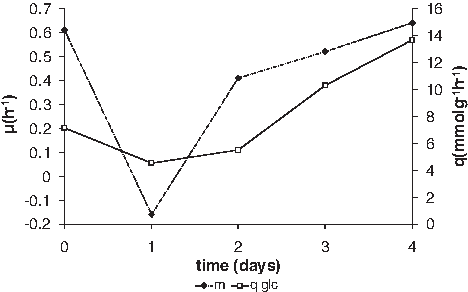
Growth rate and specific glucose consumption of E. coli in the presence of 10 μg/ml amoxicillin. Open squares: specific glucose consumption; closed diamonds: specific growth rate.
The growth rates of E. coli varied more under the influence of tetracycline than of amoxicillin. Tetracycline halved the growth rate of wild-type cells at a level of 1 μg/ml, whereas adapted cells grew slightly faster at 4 μg/ml and at a slightly reduced rate only at 1 μg/ml (Fig. 6). Nonadapted cells did not grow at 4 μg/ml. The adaptation did not come at the cost of maintenance energy. Instead, the specific glucose consumption decreased, indicating that the metabolism was hampered by the lower protein synthesis caused by exposure to tetracycline.
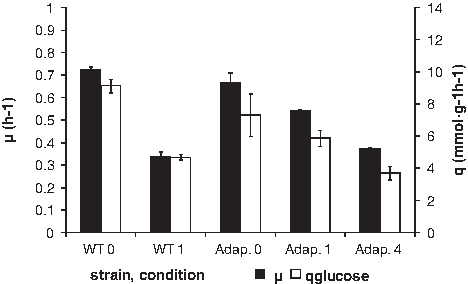
Growth rate and specific glucose consumption rates of E. coli in the presence or absence of tetracycline. WT0: wild type, no antibiotics; WT1: wild-type strain in the presence of 1 μg/ml tetracycline; Adap. 0: strain adapted to grow at 16 μg/ml tetracycline, no antibiotics; Adap 1: strain adapted to grow at 16 μg/ml tetracycline growing in the presence of 1 μg/ml; Adap. 4: strain adapted to grow at 16 μg/ml tetracycline growing in the presence of 4 μg/ml.
Enrofloxacin reduced both the growth rate and the specific glucose consumption of E. coli (Fig. 7). The resistant strain and wild-type cells had similar rates in the absence of enrofloxacin and the resistant strain was not influenced by exposure to 0.25 μg/ml. The same low concentration strongly reduced growth and glucose consumption rates of the wild-type cells, but 128 μg/ml was needed to affect the rates of the resistant strain.
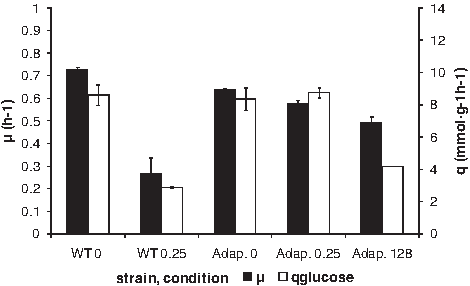
Growth rate and specific glucose consumption rates of E. coli in the presence or absence of enrofloxacin. WT0: wild-type strain growing in the absence of antibiotics; WT 0.25: wild-type strain in the presence of 0.25 μg/ml enrofloxacin; Adap. 0: strain adapted to grow at 256 μg/ml enrofloxacin, no antibiotics; Adap 0.25: strain adapted to grow at 256 μg/ml enrofloxacin growing in the presence of 0.25 μg/ml; Adap. 128: strain adapted to grow at 256 μg/ml enrofloxacin growing in the presence of 4 μg/ml.
Cross resistance of E. coli adapted to one of the three antibiotics used in this study for the two other was moderate (Table 1). The increase of the MIC for another antibiotic than the one to which the cells were adapted never exceeded a factor of 4 compared with the wild-type cells.
MIC, minimal inhibitory concentration.
Discussion
Antibiotic resistance can be brought about by phenotypic adaptation, by mutation of genes coding for proteins involved in regulatory processes, or by the acquisition of resistance genes from the environment. 14 In E. coli, the multiple antibiotic resistance gene confers multiple resistances upon exposure to aromatic compounds. 2 Still, the increase of the MIC, indicating de novo build-up of resistance, seems to occur in a modular fashion that differs for each of the antibiotics tested. Exposure to a twice higher concentration during growth did not change the MIC in the case of amoxicillin, but resulted in a twice higher MIC in the case of tetracycline, whereas the induced MIC for enrofloxacin far exceeded the concentration that induced it. In the cases of the bactericidal antibiotics enrofloxacin and amoxicillin, but not that of the bacteriostatic agent tetracycline, the higher MIC levels remained for the duration of the experiments and are thus probably based on mutations.
The different outcome for each antibiotic used in this study reflects the diversity of the mechanisms involved in the de novo acquisition of antibiotic resistance. 32 This line of reasoning may superficially seem to contradict the radical-based mechanism proposed by Kohanski and colleagues. 22 They suggest that the process of adaptation is independent of the antibiotics used, as long as it is bactericidal and not bacteriostatic. However, variation in the build-up of resistance did not only occur between antibiotics, but also in replicate experiments using the same antibiotic. Therefore, the outcome may point toward biological variation in the mutations that occur as a result of SOS-induced mutagenesis and not toward a different mechanism that initiates mutations. In this study, the main difference is that exposure to the bactericidals enrofloxacin and amoxicillin induces a permanent change of MIC, whereas acquired resistance against the bacteriostatic tetracycline seems to be rapidly reversible. This observation can very well be explained by the radical-based mechanism, 22 because, if the formation of hydroxyl radicals is indeed the main mechanism, bactericidals induce mutations rather than adaptation. As an additional support we found moderate cross-resistance, as would be expected if a common mechanism would cause adaptation, although the effect was too small to support major conclusions. Finally, the least resistance for other antibiotics was induced by adaptation to the bacteriostatic tetracycline, as predicted by the radical-based theory. 22
A large number of mechanisms that can contribute to antibiotic resistance through upregulation of existing cellular systems are known. 7 Several examples of electrochemical gradient-linked pumps that normally control, for example, pH but are converted to antimicrobial efflux pumps have been described. 23 Efflux pumps for a variety of compounds can be converted to export antibiotics. 36 Functional mutations introduced in the laboratory caused antibiotic resistance after about eight cycles of one or two mutations. 13 In general, rapid adaptation of proteins to new functions often occurs. 1 Resistance to quinolones is acquired stepwise by a series of mutations. 8 Therefore, the rapid and variable de novo development of antibiotic resistance by E. coli shown here is not surprising. The variation is probably caused by the different physiological roles of the adapted mechanism in the absence of antibiotics. Interaction between the different mechanisms is thought to cause accelerated development of resistance. 15
Steadily increasing enrofloxacin levels caused a sudden jump in MIC, far exceeding the level of exposure, in agreement with the conclusion of Hopkins and colleagues that fluoroquinolone resistance is multifactorial and due to one or more separate mutations. 16 The relationship between exposure and resistance is in agreement with the inverted U relationship found earlier by Tam and colleagues. 40 The mutations are often attributed to the GyrA or GyrB genes coding for the gyrases involved in DNA replication. 17 On the other hand, active efflux is the mechanism for fluoroquinolone resistance in Streptococcus pneumoniae.18,26 The variations in the MIC after removal of enrofloxacin suggest that both mechanisms, mutation of a gyrase and active efflux, can be operative at the same time, if it is assumed that one of the two adaptations is permanent and the other is reduced over time.
The different physiological reactions of E. coli to the three antibiotics used in this study are in line with what is known about these mechanisms. Low concentrations of tetracycline affect many genes, 44 for example, overexpressing several efflux pumps, 42 and therefore, both growth rate and specific glucose consumption are affected. Enrofloxacin interferes with DNA replication, reducing growth rate, and most likely, as a consequence, causes specific glucose consumption. Prolonged exposure to amoxicillin results in growth and metabolic rates that barely differ from those in the absence, as was observed in Mycobacterium tuberculosis. 12
The outcome of this study suggests that exposure to levels of antibiotics below the MIC and certainly below the mutant prevention concentration 24 causes the greatest risk for the development of resistance. The initial increase of the MIC may be just 1 order of magnitude, but if it is followed by a regular therapeutic dose, the target organism may become increasingly resistant. A comparable conclusion was drawn based on an in vivo and a pharmacodynamic study by Tam and colleagues.40,41 Our data support the notion that, because of the rapid development of resistance against fluoroquinolones, only high initial concentrations can be effective.33,34 The advice to use the highest recommended dose when treating patients 37 seems therefore very sensible.
An area of special concern is the use of antibiotics in agriculture. Antibiotics are often mixed into feed or drinking water. Especially in the first case, carry-over from medicated to supposedly antibiotic-free feed is known to occur. 19 When animals consume feed contaminated with low levels of antibiotics, for example, between 1% and 5% of the therapeutic dose, their microbiota might develop enough resistance to withstand therapy and become even more resistant. Antibacterial growth promoters also contribute to high levels of resistant bacteria in the feces. 3 The dung of these animals can then be a source of resistance genes continuously released into the environment. In a farm environment, abundant exchange with microbiota from natural environments can be expected. These microbiota contain large numbers of resistance genes.6,27 Transfer of resistance genes to pathogenic bacteria will occur most under selective pressures that allow some growth. When higher levels are introduced, resistant strains become dominant. 28 Exposure to low levels of antibiotics therefore clearly poses most risk. This contradicts one of the main assumptions made questioning the threat of usage of antibiotics in food animals, specifically that low levels of exposure would not be effective in selecting for resistance. 35 With time the resistance selected for in the agricultural sector will transfer to the human sector. In fact, the evidence is that this is already occurring for quite some time.5,9,10,43
Footnotes
Acknowledgments
This study was financed by the Netherlands Food and Consumer Product Safety Authority. The authors thank Drs. S. Brul and J.J. Collins for constructive criticism.
Disclosure Statement
No competing financial interests exist.