
Abstract
We analyzed the prevalence of resistance to extended-spectrum cephalosporins (ESCs) among clinical strains of Salmonella enterica collected by the Laboratory of Clinical Microbiology in the University Clinical Hospital Lozano Blesa in the region of Aragón (Spain), for which very few epidemiological information exists. A total of 2,092 strains of S. enterica were identified in stool samples from patients with gastroenteritis. Five isolates showed an extended-spectrum beta-lactamase (ESBL) phenotype: four isolates of S. enterica serotype Virchow harbored the ESBL-encoding blaCTX-M-9 gene and an isolate of serotype Enteritidis carried a blaCTX-M-1 gene, which, to the best of our knowledge, is described here for the first time in this serotype of S. enterica. The five ESC-resistant isolates were also resistant to spectinomycin, streptomycin, kanamycin, sulfonamides, tetracycline, and trimethoprim as well as to nalidixic acid. The ESBL isolate of serotype Enteritidis, however, remained susceptible to kanamycin and nalidixic acid. A class 1 integron of 1.5 kb was detected for the four serotype Virchow isolates with the gene cassette dfrA16–aadA2. The blaCTX-M-9 gene was carried by an ∼300-kb IncHI2 conjugative plasmid in the case of the S. enterica serotype Virchow isolates. The blaCTX-M-1 gene was carried by an ∼100-kb IncI1-N conjugative plasmid for the serotype Enteritidis ESC-resistant isolate. All the four ESC-resistant strains of S. enterica serotype Virchow clustered together in a XbaI pulsed-field gel electrophoresis, which also revealed a strong similarity between them and some pulsotypes of S. enterica serotype Virchow from France.
Introduction
In Spain, the first ESBL-producing Salmonella enterica isolate was described in 1996. 26 It was an S. enterica serotype Othmarschen producing TEM-27, which caused a nosocomial outbreak in Madrid. Thereafter, S. enterica isolates producing ESBLs CTX-M-9, CTX-M-27, TEM-52, or cephamycinase CMY-2 were reported sporadically in humans or animals.4,13,34 A recent study on randomly selected strains from different hospitals in Spain identified 27 (0.26%) human isolates of S. enterica producing ESBLs or cephamycinases between 2001 and 2005, 16 and in a specific study about S. enterica serotype Virchow, 79 of 504 (15%) isolates recovered from 14 of the 17 provinces in Spain were ESBL producers, 48 of them carrying a blaCTX-M-9 gene. 18
In 2000, in Aragón, a northwestern region of Spain (1,326,918 inhabitants), 129 foodborne outbreaks that affected 2,030 people (including 1,464 in Saragossa), resulting in 103 hospitalizations, were reported. S. enterica was found to be the causative agent in 62% of the cases. In 2005, a foodborne outbreak was reported, with 179 cases of salmonellosis from the whole region (109 occurring in Saragossa), 19 patients needing hospitalization. This outbreak was epidemiologically related to the consumption of locally farmed chicken (www.aragón.es). Very few studies have been published about the epidemiological situation of S. enterica or about the profile of antimicrobial resistances shown by them, both for the whole region of Aragón and for the Sanitary Area Saragossa 3. The last clinical study showed a continuous increase of isolations of S. enterica, from 1994 (118 isolates) to 2000 (287 isolates). 29
In this study, we assessed the frequency of different serotypes and the prevalence of ESC resistance among S. enterica isolates obtained in Saragossa during an 8-year period (2001–2008). The characterization of the beta-lactamase genes, their genetic support, and class 1 integron cassettes was done on ESC-resistant S. enterica isolates.
Materials and Methods
Strains
S. enterica isolates were recovered from stool samples at the Laboratory of Clinical Microbiology of the University Hospital H.C.U. “Lozano Blesa” between 2001 and 2008. The Hospital “Lozano Blesa” is a 900-bed teaching hospital located in Saragossa, the capital city of Aragón. It is the reference hospital of the Sanitary Area Saragossa 3 (268,624 inhabitants in 2005). During the study period, the Laboratory of Clinical Microbiology received 59,977 stool samples, and a pathogen was found in roughly 10% of the samples. S. enterica was the main agent with 2,092 isolates (only one isolate per patient per month was considered).
S. enterica strains were identified using the WIDER system (Soria Melguizo, Madrid, Spain) 36 and serotyped on the basis of somatic O and both phase 1 and phase 2 flagellar antigens by agglutination tests with antisera (Bio-Rad, Marnes la Coquette, France), as specified by the White-Kauffmann-Le Minor scheme. 17
Antimicrobial susceptibility
Antimicrobial susceptibility was first carried out for all the isolates using the WIDER system according to the recommendations of the Clinical and Laboratory Standards Institute guidelines. 9 For ESCs, the antibiotic concentration range was 0.12–8 μg/ml for cefotaxime (CTX) and 0.5–16 μg/ml for ceftazidime. All strains of Salmonella showing a decreased susceptibility to one or both of these antibiotics, meaning a minimal inhibitory concentration (MIC) ≥1 μg/ml, but remaining susceptible to cephamycins and to the association with clavulanic acid were selected for further analysis, in accordance with the classical definition of ESBLs. 31
ESBL phenotype was detected by the double-disk synergy method 20 and by measuring the MIC for ceftriaxone (CRO) ceftazidime and CTX with and without clavulanic acid using the ESBL detection E-test strips (AB Biodisk, Solna, Sweden).
Additional testing was carried out by the disk-diffusion method with 32 antimicrobial drugs (Bio-Rad), as previously described. 38
Polymerase chain reaction amplification of antimicrobial resistance genes and sequence analysis
Total DNA was extracted using the InstaGene matrix kit (Bio-Rad) according to the manufacturer's recommendations. The resistance genes blaTEM, blaSHV, blaOXA-1 group, blaCTX-M, and class 1 integron gene cassettes were amplified by polymerase chain reaction (PCR) as described previously. 38 Also qnr, qepA, and aac(6′)-Ib genes were investigated when resistance to nalidixic acid (NAL) was observed, as previously described.30,33
Sequencing was performed at the “Plateforme de Génotypage des Pathogènes et Santé Publique, PF8” (Institut Pasteur). The nucleotide sequences and the deduced protein sequences were analyzed with EditSeq and Megalign software (Dnastar, Madison, WI). The BLASTN program of NCBI (National Center for Biotechnology Information, U.S. National Library of Medicine, Bethesda, MD) was used for database searches.
Resistance transfer determination
A resistance transfer experiment was carried out on solid media, using either Escherichia coli K-12 BM14 resistant to sodium azide or E. coli C1a resistant to NAL as the recipient strain. 12 Transconjugants were selected on Drigalski agar (Bio-Rad) supplemented with CRO (20 μg/ml) and sodium azide (500 μg/ml) or NAL (64 μg/ml). Three E. coli transconjugants were arbitrarily selected for each experiment.
Plasmid analysis
Plasmid DNA from parental and transconjugant strains was analyzed by pulsed-field gel electrophoresis (PFGE) after linearization with the S1 nuclease enzyme, as described previously. 12 PCR-based replicon typing analysis was performed as described by Carattoli et al. 7 The 18 primer pairs targeting FIA, FIB, FIC, HI1, HI2, I1-Iγ, L/M, N, P, W, T, A/C, K, B/O, X, Y, F, and FII replicons were used in separate PCRs.
PFGE typing
The genetic diversity of the four S. enterica serotype Virchow isolates resistant to ESCs was assessed by PFGE of genomic DNA digested with XbaI (Roche, Mannheim, Germany), as previously described. 38 The running conditions and the molecular size marker were as described in the standardized PulseNet protocol (www.cdc.gov/pulsenet/). BioNumerics 4.0 (Applied Maths, Saint-Martens-Latem, Belgium) was used for image normalization and construction of similarity matrices. Bands were assigned manually. Clustering was carried out by the unweighted pair-group method with arithmetic averages based on the Dice similarity index, using a 1% optimization parameter and 1% band-position tolerance. The results were compared with PFGE profiles of S. enterica from the French National Reference Centre for Salmonella (FNRC-Salm) database.
Results
Between 2001 and 2008, 2,092 S. enterica isolates were identified (one per patient within a period of 30 days), with a continuous tendency to decrease from 2002 (394 isolates) to 2008 (138 isolates). The distribution of serotypes is presented in Table 1. Enteritidis was the predominant serotype, accounting for 52% of all the isolates.
The results of susceptibility testing are shown in Tables 2 and 3. Of the 2,092 isolates, 387 (18.5%) were susceptible to all antimicrobial agents tested. The most frequent types of resistance observed concerned ampicillin (increasing from 33.5% in 2001 to 59.4% in 2008), NAL (47.2% in 2005 but decreasing since then to a 21.7% in 2008), and trimethoprim/sulfamethoxazole (peaking at 15% in 2004). Although up to 17 isolates had an MIC for CRO of ≥1 μg/ml, only five isolates (0.24%) of serotypes Enteritidis (isolate 06-424914) and Virchow (isolates 02-236146, 02-214992, 03-1672608, and 04-1831083) showed an ESBL phenotype determined by double-disk diffusion test and ESBL-Etest® (Table 3). All five isolates were susceptible to cephamycins, association with clavulanic acid and carbapenems. The mechanisms of resistance to beta-lactam antibiotics of these five isolates were investigated and are shown below. The other 12 isolates with an MIC for CRO of ≥1 μg/ml were resistant to cefoxitin and gave a negative double-disk synergy test (despite the use of several ESC disks placed at distances of both 15 and 20 mm from the clavulanic acid disk); therefore, they were considered as cephamycinase-producing isolates. No further molecular characterization of these isolates has been possible, as they were accidentally discarded as no-ESBL producers.
All measurements are E-test minimal inhibitory concentrations (μg/ml) except for CRO and CIP, and values represent diameters on disk-diffusion test (mm).
ESBL, extended-spectrum beta-lactamase; TC, E. coli transconjugant; S, streptomycin; Sp, spectinomycin; K, kanamycin; Su, sulfonamides; Te, tetracycline; Tm, trimethoprim; Nal, nalidixic acid; CTX, cefotaxime; CTL, cefotaxime/clavulanic acid; CAZ, ceftazidime; CZL, ceftazidime/clavulanic acid; CPM, cefepime; CPL, cefepime/clavulanic acid; CRO, ceftriaxone; CIP, ciprofloxacin; Inc, plasmid incompatibility group.
The serotype Enteritidis isolate 06-424914 was also resistant to aminoglycosides (streptomycin, spectinomycin), trimethoprim–sulfamethoxazole, and tetracycline (Table 3). The serotype Virchow isolates 02-236146, 02-214992, 03-1672608, and 04-1831083 had a similar susceptibility profile, except an additional resistance to kanamycin and NAL, with a decreased susceptibility to ciprofloxacin (MIC of 0.25–0.5 mg/L).
PCR and sequence analysis detected in the five isolates both the penicillinase blaTEM-1 gene and an ESBL blaCTX-M gene. Serotype Enteritidis isolate 06-424914 contained the blaCTX-M-1 gene, whereas serotype Virchow isolates contained the blaCTX-M-9 gene. The serotype Virchow isolates harbored a 1.5-kb class 1 integron containing a dfrA16–aadA2 gene cassette known to confer resistances to trimethoprim and streptomycin and spectinomycin, respectively. No class 1 integrons were found for the Enteritidis isolate.
In serotype Enteritidis isolate 06-424914, the blaCTX-M-1 gene was carried by a ≈100-kb conjugative IncI1-IncN multireplicon plasmid. Other resistance determinants, affecting streptomycin, spectinomycin, sulfamides, trimethoprim–sulfamethoxazole, and tetracycline, were cotransferred to transconjugants with CRO resistance.
In serotype Virchow isolates, the blaCTX-M-9 gene was carried by a ≈300-kb conjugative plasmid of replicon IncHI2. The same resistance determinants as listed for the serotype Enteritidis isolate 06-424914 were cotransferred to transconjugants with CRO resistance. The resistance to NAL was due to a chromosomal mutation on the QRDR region of the gyrA, leading to a substitution of a serine in position 83 by a phenylalanine, as previously described. 22
In this study, all the CTX-M-9–producing S. enterica serotype Virchow isolates clustered together independently to their geographic area of origin, might it be Spain or France, as it was shown in a database comparison between the isolates from Saragossa and some pulsotypes of the FNRC-Salm (Fig. 1).
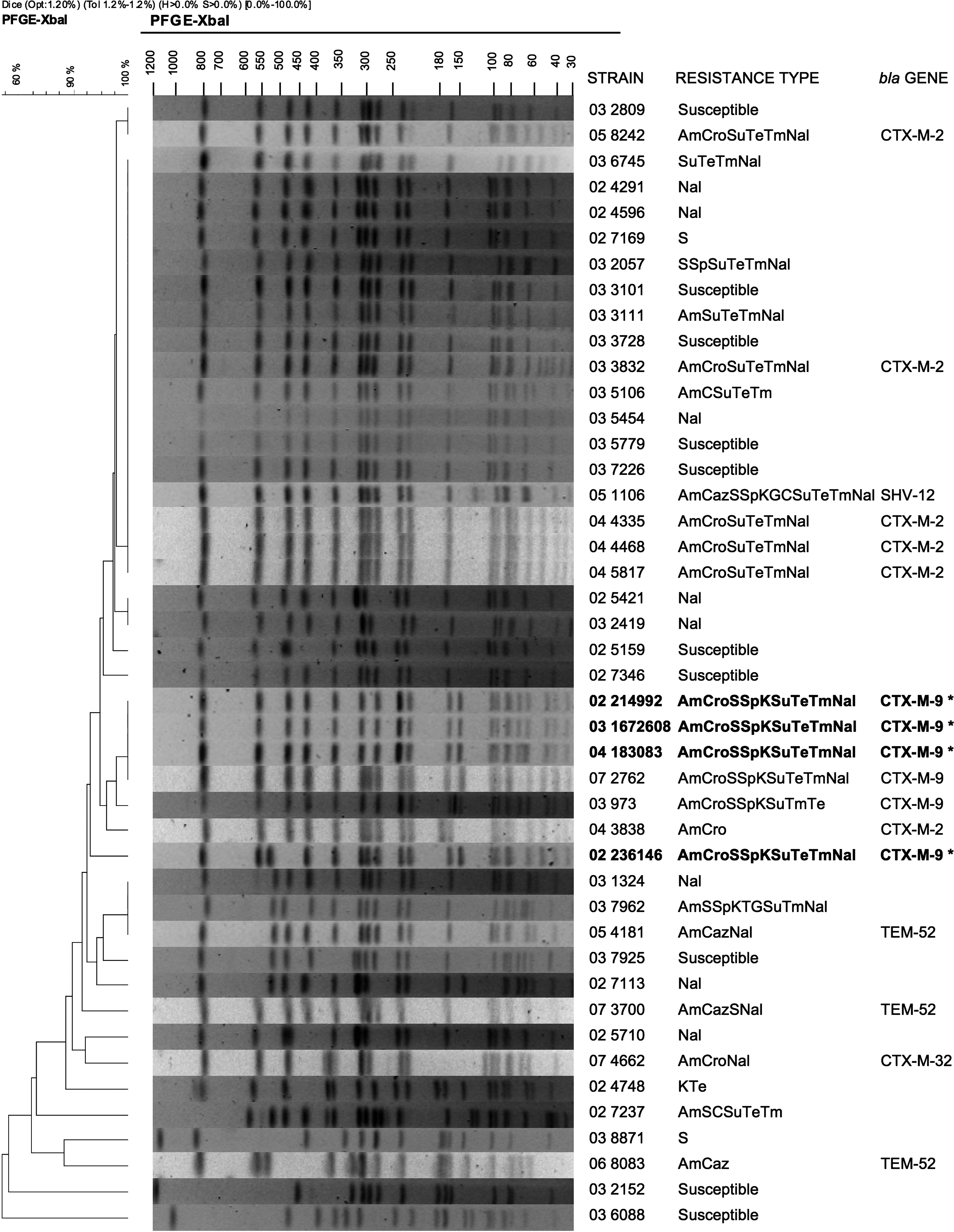
Phylogenetic analysis of XbaI-PFGE profiles obtained from S. enterica serotype Virchow isolates from Saragossa and France. The two first numbers indicate the year of isolation of the strain. The strains from Saragossa are outlined in bold and with asterisks (*). The remaining strains belong to the database of the French National Reference Centre for Salmonella. Am, amoxicillin; Cro, ceftriaxone; Caz, ceftazidime; S, streptomycin; Sp, spectinomycin; K, kanamycin; G, gentamicin; Su, sulfonamide; Te, tetracycline; Tm, trimethoprim; Nal, nalidixic acid.
Discussion
Since the first blaCTX-M genes were described in the early 1990s, S. enterica has been one of the first species of Enterobacteriaceae to be identified harboring this kind of resistances. 1 CTX-M-9, first reported in Spain in 1996, was produced by an E. coli human isolate, and S. enterica serotype Virchow carrying a blaCTX-M-9 appeared just a few years later. 34 Retrospective studies in the United Kingdom have found strains that were isolated in the 1990s from patients with a history of traveling abroad. 19 Strains isolated from both humans and poultry were reported in France, suggesting an interspecies transfer, which affected several serotypes (Virchow and Enteritidis among them) and different ESBL enzymes (CTX-M-2, TEM-52, CTX-M-9). 38 This is supported as well by the latest studies on poultry in Spain. 5
The ESBL gene blaCTX-M-1, first reported in Germany in 1996, was harbored by an E. coli strain, 2 and the first case of S. enterica producing CTX-M-1 was a strain of serotype Typhimurium isolated in France. 21 Recent studies found S. enterica serotypes Enteritidis carrying the blaCTX-M-32 gene and Litchfield carrying the blaCTX-M-1 gene in Spain, in relation to conjugative plasmids of IncN and IncI1, respectively. 16 The plasmid carrying the blaCTX-M-1 gene had also a similar size (110 kb) to the plasmid found for the Enteritidis isolate of our study, and both share a similar multidrug resistance profile and lack a class 1 integron. E. coli strains carrying this ESBL gene on a IncI1 plasmid have been isolated in Italy and France from both humans and animals (poultry and dogs).14,15 Another study on E. coli strains recovered from human samples in France showed that the blaCTX-M-1 gene was carried by either IncI1 or IncN plasmid. 24 Multireplicon plasmids do often occur, 6 but an IncI1-N has not yet been described. Previous findings in animals, mainly poultry, of both ESBL blaCTX-M-1 and blaCTX-M-9 harbored in IncI or IncHI2 plasmids suggest that poultry might play an important role as a reservoir for these bacteria.16,38
The first ESBL S. enterica strains were detected in our laboratory in 2002. 8 During this 8-year prospective study, we have found a total of four isolates of serotype Virchow harboring a blaCTX-M-9 gene and one S. enterica serotype Enteritidis with the blaCTX-M-1 gene, all of them during the period 2002–2006. ESBL strains were no longer recovered after 2006. Although the final rate of ESBL among the total figures was rather low (0.24%), we call out the fact that those four isolates of serotype Virchow producing a CTX-M-9 occurred in 26.6% of all isolates of that serotype during the time period of this study. The four strains of serotype Virchow appear to carry the same IncHI2 conjugative plasmid previously described in CTX-M-9–producing strains from Spain (in the region of La Rioja, adjacent to Aragón, 32 in Barcelona, 10 and in Madrid 28 ). These works describe plasmids of the same size, belonging to the same incompatibility group IncHI2, and carrying the same gene cassette dfrA16—aadA2 within the complex class 1 integron In60, altogether with the blaCTX-M-9 gene.3,10,27,32 This has finally become one of the most predominant combinations of S. enterica serotype and ESBLs in Spain and Portugal. 23
Clonal transmission of multidrug resistance has been proven in isolates from poultry and humans.32,38 The PFGE pulsotypes, with more than 96% of similarity among the four ESBL Virchow isolates, and the phenotype, gene cassettes, and plasmids found suggest that they could be associated with the clonal spread of S. enterica serotype Virchow PT19 previously described in Spain and France.18,38
Footnotes
Acknowledgments
The authors acknowledge Dr. Joseph Dragavon for the careful reading of this manuscript. M. Pardos de la Gándara was the recipient of the following grants during this work: Grant for Short Term Research in Clinical Centres Abroad by the S.E.I.M.C. (2007); Grant “Project Europe XXI” of the CAI-Social Action and the Government of Aragón for Short Term Research Abroad (2008); Postdoctoral/Postspecialization Fellowship for Research in Universities or Centres Abroad from the “Alfonso Martín Escudero” Foundation, Spain (2009); Student Travel Fellowship from the SEIMC (Spanish Society of Infectious Diseases and Clinical Microbiology) for the Meeting in Seville, Spain (2009).
Disclosure Statement
The authors declare that there are no commercial associations that might create a conflict of interest in connection with this article. M. Pardos de la Gándara has received research funding from the Sociedad Española de Enfermedades Infecciosas y Microbiología Clínica (Beca Martín Luengo 2007), the CAI-Gobierno de Aragón (Proyecto Europa XXI 2007–2008), and the Fundación Alfonso Martín Escudero (Becas para estudios en el extranjero 2008–2009). C. Seral García, F.J. Castillo García, C. Rubio Calvo, and F.X. Weill declare absence of any relationship or any degree of conflicting or dual interest (financial or of any other nature) that may affect professional judgment in relation to this article.