
Abstract
We isolated a clinical Escherichia coli strain with an antimicrobial resistance phenotype characteristic for the expression of an AmpC beta-lactamase. Molecular methods revealed a novel, plasmid-localized variant of CMY-2 with a substitution of valine 231 for serine (V231S), which was designated CMY-42. Like the CMY-2-like AmpC beta-lactamase CMY-30, carrying the substitution V231G, CMY-42 displayed increased activity toward expanded spectrum cephalosporins. This finding supports the hypothesis that a bulky side chain at position 231 (Ambler's position 211) may pose a steric clash with certain cephalosporins hindering the access of the AmpC beta-lactamase; however, additional phenomena may account for the observed hydrolytic properties.
Introduction
Materials and Methods
Bacterial isolates and antimicrobial susceptibility testing
The E. coli VA1171/10 strain was isolated from a surgical wound at the University hospital Hamburg-Eppendorf in Germany and was characterized biochemically (Vitek II; Biomerieux) and by MALDI-TOF mass spectrometry (Bruker Daltonics). The E. coli strain DH10B (Top 10; Invitrogen) was used for transformations and cloning.
Antimicrobial susceptibility testing
Minimum inhibitory concentrations (MIC) for the reported antibiotics were determined by Etest (AB Biodisk) and interpreted according to EUCAST criteria. Presence of AmpC and ESBL beta-lactamases was analyzed with the AmpC & ESBL ID quadruple disc test (MAST Diagnostics) in which susceptibility to cefpodoxime is determined alone, in combination with boronic acid and clavulanic acid, or both.
DNA manipulations
Molecular screening for the presence of plasmid-borne ampC genes was performed with a previously described multiplex polymerase chain reaction (PCR). 8 The full-length plasmid-localized ampC gene of VA1171/10 was amplified using the primers CIT-F: AACACACTGATTGCGTCTGAC and CIT-R: TGTTAAGTGTAGATGACAACA, and the PCR product was directly sequenced. Reference sequences were retrieved from www.lahey.org/Studies/other.asp#table1. The novel sequence of CMY-42 was deposited at GenBank (GenBank accession number: HM146927) and the Lahey clinic beta-lactamase databank.
PCR mapping was performed to characterize the genetic environment of cmy-42. A set of primers annealing to ISEcp1, cmy-2, blc, and sugE was used to produce overlapping amplicons. 12 The latter were sequenced on both strands and data were assembled using the DNASTAR software.
Plasmid DNA from VA1171/10 was isolated using the Nucleobond AX kit (Macherey und Nagel) according to the manufacturer's instructions. Plasmid DNA was subsequently transformed into E. coli DH10B cells by electroporation. AmpC-plasmid harboring transformants were selected on an ESBL screening agar (Oxoid).
The CMY-42 and the CMY-2 ampC genes were amplified with the primers CMY-F: AGGATCCATGATGAAAAAATCGTTATGCTG and CMY-R: AGTCGACTTATTGCAGCTTTTCAAGAATG from strain VA1171/10 and a CMY-2- carrying E. coli isolate from the strain collection of the institute. ampC genes were digested with BamHI and SalI, and inserted into the previously described pACYC184 vector harboring a plac promoter. 7 This resulted in plasmids pACYC-lac-CMY-42 and pACYC-lac-CMY-2. pACYC-lac-CMY-30 was generated by site-directed mutagenesis of pACYC-lac-CMY-2 with the primers CMY-30-mut-F: GAAGGGAAGCCCGTACACGGTTCTCCGGGACAACTTGAC and CMY-30-mut-R: GTCAAGTTGTCCCGGAGAACCGTGTACGGGCTTCCCTTC using the QuikChange-II-site directed mutagenesis kit (Stratagene). All constructs were verified by sequencing. The control vector contained a cDNA for red fluorescent protein (RFP) instead of a beta-lactamase (pACYC-lac-RFP). These constructs were then transformed into chemocompetent DH10B E. coli. Additionally, a pBC-SK +/− derivative plasmid carrying the cmy-42 gene that was under transcriptional control of the promoter provided by the ISEcp1 (pB-cmy-42) was constructed for purification purposes using the previously described pB-cmy-2 plasmid 12 as template, the CMY-V211S-F (5′-AAGGGAAGCCCGTACACAGTTCTCCGGGACAACTT-3′) and CMY-V211S-R (5′-AAGTTGTCCCGGAGAACTGTGTACGGGCTTCCCTT-3′) mutagenic primers and the QuikChange-II-site directed mutagenesis kit.
Isoelectric focusing
Beta-lactamase production from each strain was examined by analytical isoelectric focusing (IEF) of cell-free extracts derived by sonication. Beta-lactamase bands were observed in situ by nitrocefin staining.
Enzyme expression and purification
The pB-cmy-42 plasmid was used to transform E. coli MC4100 competent cells, the overproduced CMY-42 enzyme was released from the periplasmic space by mild sonication in 20 mM Tris/HCl pH 8, and the proteins were concentrated by ultrafiltration using Amicon 10 KDa cut-off filter. The preparation was then loaded on a Q-Sepharose column and the effluent that contained the beta-lactamase (as tested by a nitrocefin assay) was dialyzed against 20 mM NaPi pH 7 and loaded on a S-sepharose column. The bound CMY-42 was eluted with 150 to 250 mM NaCl, and the respective fractions were pooled dialyzed against 50 mM NaPi pH 7.6 and concentrated using Centricon 10 kDa cut-off. The purity of the final preparation was >90% as determined by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Protein concentration was determined by the Bradford method.
Determination of kinetic parameters
Hydrolysis of various substrates (i.e., benzylpenicillin, cephalothin, ceftazidime, cefotaxime, cefoxitin, and aztreonam) was monitored by UV spectrophotometry using a Hitachi U-2001 UV/Vis instrument. Wavelengths and extinction coefficients have been reported previously. 5 All reactions were performed in 50 mM NaCl and 50 mM NaPi (pH 7) at 25 C and steady state kinetics parameters were determined by estimating initial velocities at various substrate concentrations and then fitting the results in the Michaelis-Menten equation using nonlinear regression analysis with the aid of Prism software. In cases where Km was low the respective values were determined as inhibition constants (Ki's) by treating the substrate as competitive inhibitor and cephalothin as the reporter substrate. Inhibition by oxacillin, aztreonam, tazobactam, and RO 48-1220 was studied as described previously. 5
Conjugational transfer of ceftazidim resistance
To demonstrate the transferability of ceftazidime resistance equal amounts of late logarithmic cultures of E. coli K-12 recipient J53-1 resistant to sodium azide (MIC 512 μg/ml) but susceptible to ceftazidime (MIC <0.125 μg/ml) and E. coli donor VA1171/10 resistant to ceftazidime (MIC >32 μg/ml but susceptible to sodium azide (MIC 128 μg/ml) were mated on a 0.22 μm filter for 3 h at 37°C. The mixed culture was resuspended in sterile phosphate-buffered saline and serially diluted 10-fold, before aliquots were plated onto selective agar containing sodium azide (256 μg/ml) plus ceftazidime (0.5 μg/ml). While no cells of recipient or donor cells alone were able to grow under these conditions, cells from mixed culture grew. For individual clones of the mixed culture susceptibility testing yielded MICs of >32 and 512 μg/ml for ceftazidim and sodium azide, respectively. The frequency of transfer was calculated by dividing the viable cell count of the mixed culture by that of the donor cells.
Results
We report the identification of a novel acquired AmpC beta-lactamase from a clinical E. coli isolate (VA1171/10) that displayed phenotypical resistance to cefoxitin, cefotaxime, and ceftazidime and was, therefore, tested by multiplex-PCR for the presence of ampC genes. The isolate was recovered from an orbital wound of a patient from a maxillofacial surgery ward, who was antibiotically treated with clindamycin at that time. PCR confirmed presence of an AmpC beta-lactamase gene of the CMY-family. Analytical IEF confirmed the production of a cephalosporinase with a pI of 9.0, and also revealed an additional band at pH 5.4 probably corresponding to TEM-1. While most of the isolated plasmid-mediated AmpC beta-lactamases from E. coli at our institution are CMY-2 (DHA-1 from K. pneumoniae) amplification and sequencing of the entire ampC gene revealed a novel cmy variant that differed from cmy-2 by two nucleotides, a double frame shift (ins690_691A; del692T) that resulted in a Ser for Val-231 substitution (Ambler's position 211). The novel beta-lactamae variant was designated CMY-42.
Plasmidic localization of the cmy-42 gene was confirmed by isolating a plasmid (approximate size 67.5 kb) from VA1171/10 and transforming it into the DH10B E. coli strain resulting in DH10B-p1171. The transformed strain produced only the class C beta-lactamase as determined by IEF. Transferability of the ceftazidime resistance trait was demonstrated by a conjugation experiment using E. coli J53-1 (aziR) as the recipient. The transfer frequency was determined as 2.7 × 10–7. The ampC gene was then amplified and sequenced from DH10B-p1171. cmy-42 was part of a C. freundii–derived segment comprising blc and sugE (Fig. 1). A ISEcp1 was identified at a distance of 116-bp from the cmy-42 start codon. The tnpA gene of ISEcp1 was disrupted by an IS1 insertion. At the boundaries of the latter element 7-bp target site duplications (TTTGAAG) were detected indicating mobilization of IS1 into ISEcp1 via a transposition mechanism. Of note, IS1 was not previously identified in any of the cmy-encoding structures.
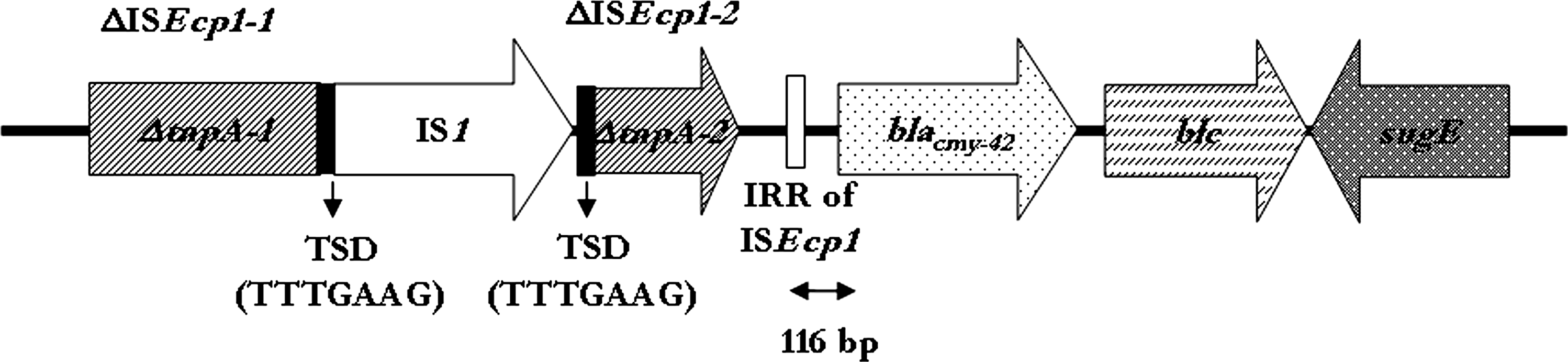
Schematic representation of the genetic environment of cmy-42. Arrows indicate the direction of transcription.
Susceptibility testing revealed similar resistance profiles for most beta-lactams in VA1171/10 and DH10B-p1171 except in the cases of cefoxitin, cefotaxime, and cefepime. Although the transformed strain exhibited lower MIC values for the above antibiotics than the wild type, it was still resistant (Table 1). As certain variants of the chromosomal ampC beta-lactamase gene of E. coli were shown to confer reduced susceptibility to cefepime, 6 the full ORF and the 5′-promoter region of the ampC gene of VA1171/10 was analyzed as described. 6 Except for one silent nucleotide exchange (nt354T > C; Asn > Asn) the sequence matched the reference sequence of E. coli ATCC 8379 (NC_010468.1). A commercially available ATCC 8379 strain (Oxoid) was fully susceptible resembling DH10B (data not shown). Further, presence of an OXA-30 beta-lactamase, which confers resistance to cefepime, 3 was excluded using previously described primers 2 (positive control described in Ref. 1 ). The concomitant presence of an ESBL was excluded by testing the strain with the AmpC & ESBL ID quadruple disc test. Thus, the reduced susceptibility to cefepime in VA1171/10 currently remains unexplained. Further, the MICs of ertapenem for both strains were slightly elevated compared to the untransformed DH10B strain. Susceptibilities to tobramycin, gentamicin, ciprofloxacin, and tetracycline were also tested. While the MICs of ciprofloxacin and tetracycline for VA1171/10 were ≥32 and ≥256 μg/ml, respectively, MICs of ciprofloxacin for both DH10B-p1171 and DH10B were 0.006 μg/ml and of tetracycline 4 μg/ml. All strains were fully susceptible to the tested aminoglycosides and exhibited MICs of 0.5–1 μg/ml.
ATM, aztreonam; FOX, cefoxitin; CTX, cefotaxime; CAZ, ceftazidime; FEP, cefepime; ERT, ertapenem; IMI, imipenem; MEM, meropenem; MIC, minimum inhibitory concentration.
Another previously published variant of CMY-2, CMY-30, also harbors an amino acid exchange at Ambler's position 211, yet with a substitution of valine 211 to glycine.5,10 CMY-30 was described to confer higher resistance to aztreonam and expanded spectrum cephalosporins compared to CMY-2. 5 To analyze whether CMY-42 also has an expanded substrate spectrum compared to CMY-2, we inserted the cmy-42, cmy-30, and the cmy-2 genes into the pACYC184 vector under the control of a plac promoter. The constructs and the control plasmid pACYC-lac-RFP were used to transform DH10B E. coli and the MICs of a number of beta-lactams for the transformants were determined and compared (Table 1). Transformation of all AmpC beta-lactamase plasmids resulted in a significant rise of the MICs of cefotaxime, ceftazidime, and aztreonam and a small increase of the MIC of cefepime as compared to the control plasmid. The isogenic CMY-42 and CMY-2 carrying strains consistently differed in their susceptibility profile for beta-lactam antibiotics with CMY-42 conferring twofold higher MICs of cefotaxime, ceftazidime, and aztreonam. Under our experimental conditions, CMY-30 also conferred higher resistance levels to cefotaxime (fourfold) and aztreonam (twofold) but not to ceftazidime as compared to the CMY-2-producing clone. MICs of carbapenems were unaffected by the presence of any of the AmpC beta-lactamases. The generally lower levels of resistance and lack of activity toward ertapenem conferred by CMY-42 expressed from the pACYC184 plasmid as compared to the natural plasmid may be due to lower activity of the artificial promoter.
Biochemical characterization of the CMY-42 enzyme confirmed its extended spectrum character denoted also by the MIC values. Compared to CMY-2 and CMY-30, CMY-42 exhibited a similar mode of interaction with benzylpenicillin, cephalothin, and cefoxitin, as the hydrolytic efficiency, kcat, and Km values were merely the same between the three enzymes (Table 2). The Ser211 enzyme exhibited increased activity toward oxyimino-cephalosporins (cefotaxime and ceftazidime) that was characterized by high turn-over rates (kcat) compared to CMY-2. Hydrolysis of ceftazidime was similar between CMY-42 and CMY-30 with the latter enzyme displaying a slightly decreased kcat value. In contrast, cefotaxime was hydrolyzed at lower rates by CMY-42 (eightfold decrease of kcat) as compared to CMY-30. Finally, aztreonam that possesses an oxyimino side chain identical to that of ceftazidime was hydrolyzed at measurable rates by CMY-42 though lower than those of CMY-30 (Table 2). Inhibition profile of CMY-42 was typical for a class C beta-lactamase 5 and followed the scheme: oxacillin (Ki = 0.2 nM) > aztreonam (6 nM) > RO 48–1220 (70 nM) > tazobactam (700 nM).
Values are the means of four independent measurements.
Values are those given at reference 2 except aztreonam measurements, which were obtained during the present study.
Or Ki values using cephalothin as the reporter substrate.
NH: No detectable hydrolysis after one hour incubation of 200 μM aztreonam with 0.1 μM of enzyme.
Concluding Remarks
We report the identification and genetic and biochemical characterization of CMY-42, a novel variant of the widespread AmpC beta-lactamase CMY-2, further expanding the AmpC beta-lactamase family. CMY-42, harboring the V211S substitution located in the Ω-loop, shows the typical profile of an AmpC beta-lactamase with activity toward cefoxitin, aztreonam, and third-generation cephalosporins. CMY-42 exhibits a consistent increase in activity against expanded spectrum cephalosporins and aztreonam as CMY-30, a V211G variant of CMY-2 cephalosporinase. This may point toward an important role of this residue in determining activity against third-generation cephalosporins. Recent studies suggested that removal of the side chain of Val211 of CMY-2 by replacement with glycine abolishes steric clashes with beta-lactam substrates, which might facilitate the deacylation process.5,11 The side chain of serine in CMY-42 is more bulky than that of glycine, which may explain the decreased hydrolysis of cefotaxime. Nevertheless, as ceftazidime hydrolysis is as high as in CMY-30, the role of position 211 is likely more complicated than previously hypothesized. 5 This will have to be further analyzed in future structural studies. The present findings underscore the evolutionary potential of the clinically important AmpC enzymes.
Footnotes
Acknowledgments
We thank Tim Lehmphul, Christin Kappert, Tatjana Claußen, and Alice Busch for technical assistance. We are grateful to Patricia Antunes for sending us the OXA-30-positive Salmonella Typhimurium strain as a positive control.
Disclosure Statement
No competing financial interests exist.