
Abstract
Abstract
In Brazil, the spread of an endemic clone of SPM-1-producing Pseudomonas aeruginosa has been reported. Recently, a higher genomic variety has been observed among the SPM-1-producing P. aeruginosa isolates. The principal aim of this study was to analyze through multilocus sequence typing (MLST) analysis whether the recently isolated SPM-1-producing P. aeruginosa descend or not from a common ancestor. A total of 50 SPM-1-producing P. aeruginosa exhibiting 11 distinct ribotyping genotypes collected from 11 different Brazilian cities were studied. Three IMP-1-producing P. aeruginosa and two non-metallo-beta-lactamase-producing P. aeruginosa isolates were included in the study as controls. For assignment of allelic numbers and subsequent determination of sequence type (ST), the obtained sequences were compared to existing sequences in the MLST database (www.pubmlst.org/paeruginosa). The eBURSTv3 software was used in this study for establishing the evolutionary relationship and phylogenetic analysis. A total of 5 different STs were identified among 55 P. aeruginosa isolates. All of the SPM-1-producing P. aeruginosa presented an identical allelic profile (ST277), except for one strain. The three IMP-1-producing P. aeruginosa strains were classified as belonging to the ST593, whereas the non-metallo-beta-lactamase-producing P. aeruginosa showed two new distinct STs, ST594 and ST595. Our study shows that SPM-1-producing P. aeruginosa isolates as well as the IMP producers evaluated in this study descend from a common ancestor.
Introduction
In 2002, when the first SPM-1-positive P. aeruginosa isolate (48-1997A) was reported, it was speculated that blaSPM-1 could be carried by a large plasmid. 25 However, the initial molecular characterization of 15 Brazilian SPM-1-producing P. aeruginosa isolates from multiple Brazilian hospitals indicated that a single epidemic clone was spread in several Brazilians regions. 7 Later, Poirel et al. described the genetic context of an SPM-1-producing strain and also showed that blaSPM-1 was flanked by two ISCR elements, designated ISCR4, which probably transposes via a mechanism called rolling-circle replication.18,26 Recently, Salabi et al. compared the genetic context of BH121 strain, which was isolated in Swiss patient, who was previously hospitalized in a Brazilian institution, to that 15 other SPM-1-positive P. aeruginosa isolated in Brazil, by pulsed-field gel electrophoresis (PFGE). 22 This analysis showed that BH121 had a DNA restriction pattern almost identical to that of the other Brazilian strains, differing by only three or four bands. Hybridization experiments performed by these authors showed that that the MBL gene was chromosomally located in all isolates. They also observed that three P. aeruginosa isolates, including P. aeruginosa BH121 and the index case strain, 48-1997, possessed two copies of blaSPM-1. The duplication of blaSPM-1 was likely to have arisen from ISCR4 transposition proceeded by homologous recombination and indicated that ISCR4 was likely to be active. 26
The aim of this study was to analyze whether the recently isolated SPM-1-producing P. aeruginosa descend or not from a common ancestor. In this study, three distinct typing methods (PFGE, automated ribotyping, and multilocus sequencing typing [MLST]) were employed for molecular characterization of an expanded set of SPM-1-producing P. aeruginosa isolates from a wider Brazilian geographic distribution. Further, we determined the ancestor relationship among SPM-1-producing P. aeruginosa using the international MLST database.
Materials And Methods
Bacterial isolates
Carbapenem-resistant P. aeruginosa isolates were submitted by 11 Brazilian hospitals spread across nine states between the years 2000 and 2004. Only one strain per patient was included in this study. All isolates received at Laboratório Especial de Microbiologia Clínica from Universidade Federal de São Paulo were cultivated on blood agar to assess purity. Species identification was confirmed using the API NF kit (BioMerieux Vitek). Three IMP-1-producing P. aeruginosa isolates and two non-MβL-producing P. aeruginosa isolates previously identified were included in this study as control strains.
blaSPM-1 genotypic detection
The presence of the blaSPM-1 gene was performed as previously described. 7 Briefly, cells were boiled to liberate the DNA and then PCR was performed using SPM-1 specific primers (forward 5′-CCTACAATCTAACGGCGACC-3′ reverse 5′-TCGCCGTGTCCAGGTATAAC-3′). Cycle parameters were 95°C for 5 minutes followed by 30 cycles of denaturation at 95°C for 1 minute, annealing at 40°C for 1 minute, and extension at 68°C for 1 minute. PCR products were observed by electrophoresis on 0.8% agarose gels stained with ethidium bromide.
Molecular typing of SPM-1-producing isolates
Each SPM-1-producing isolate and each control strain was characterized by three distinct molecular typing methods: PFGE, automated ribotyping, and MLST. The PFGE was accomplished for all the samples using the CHEF DRIII apparatus (Bio-Rad). Genomic DNA was digested with the SpeI restriction enzyme (New England BioLabs). The electrophoresis was run for 24 hr and the switch interval ramped from 5 to 90 sec for 24 h at 6 V. Photographs of ethidium bromide-stained gels were examined visually and banding patterns comparison were analyzed according to Tenover's criteria. 24 Ribotype patterns of all isolates were obtained by automated ribotyping using the RiboPrinter Microbial Characterization System (DuPont Qualicon) after digestion with the PvuII enzyme. Pattern analysis was carried out using the RiboPrinter software (version 2.2.012.0). MLST was performed according to the protocol developed by Curran et al. 3 The obtained sequences of seven house-keeping genes (acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE) were compared to existing sequences from the MLST database available at http://pubmlst.org/paeruginosa. Assignment of allelic numbers and sequence type (ST) were determined after comparison analysis. The eBURSTv3 software was used in this study for establishing the evolutionary relationship. The statistical significance of the relationships obtained was estimated by bootstrap resampling analysis (1,000 repetitions).
Results
Pulsed-field gel electrophoresis
Among the 50 SPM-1-producing P. aeruginosa isolates, 30 exhibited PFGE pattern C, 15 showed pattern C1, and 2 exhibited pattern C3. The patterns C2 and D were found each in single isolates. One out of the 50 isolates could not be typed by this technique. Five different PFGE patterns were identified among the five control strains (Table 1 and Fig. 1).

MβL, metallo-β-lactamase; MLST, multilocus sequence typing; PFGE, pulsed-field gel electrophoresis.
Automated ribotyping
Eleven distinct ribotype patterns were observed among the 50 Brazilian SPM-1-producing P. aeruginosa isolates. The majority (n = 29) of the isolates belonged to the ribotype 72-3. Ribotype 78-4 and 105-7 were observed in seven and three isolates, respectively. While the ribotypes 72-3 and 78-4 were found in distinct geographic regions, the ribotype 105-7 was found only in SPM-1-producing isolates collected from the city of Belo Horizonte (Brazil). Ribotypes 72-1 was encountered in three IMP-1-producing isolates and ribotype 69-5 was identified in two non-MβL producers.
MLST and database comparisons
Out of the 50 Brazilian SPM-1-producing P. aeruginosa strains evaluated in this study, 49 presented an identical allelic profile (acsA 39, aroE 5, guaA 9, mutL 11, nuoD 27, ppsA 5, and trpE 2) and were classified under sequence type 277 (ST277). One Brazilian isolate (P3303) collected from a Southeastern Brazilian Medical center exhibited distinct allelic profiles (acsA 38, aroE 11, guaA 3, mutL 13, nuoD 1, ppsA 2, and trpE 4) and was classified under a different ST, ST235. The three Brazilian IMP-1-producing isolates showed identical allelic profiles (acsA 38, aroE 89, guaA 3, mutL 13, nuoD 1, ppsA 2, and trpE 4) and were classified as ST593. The non-MβL-producing isolates showed different allelic profiles that were not previously reported in the MLST world database. After all, two new distinct STs, ST594 and ST595, were identified.
Comparison of Brazilian isolate MLST types to international database
The Brazilian endemic clone SPM-1-producing P. aeruginosa (ST277) showed an identical allelic profile to strain ID 275, strain Hietzing-3, which was isolated in Austria in September 2006. Compared to ST277, a single-locus variant in the nuoD allele (nucleotide 186, C-T) was observed in strain ID279, named Hietzing-11 (ST364), which was also isolated in Austria in January 2007. A similarity in five out of seven alleles (double-locus variants on guaA and nuoD alleles) was found between the SPM-1-producing P. aeruginosa isolates and the strain ID148 (ST206). This last strain was isolated in Canada in 1990 and denominated H6 sample. The eBURST displaying the relationships among the isolates from this study and the other isolates from the MLST data base is shown in Fig. 2. Among all isolates typed in this study, automated ribotyping detected 13 distinct patterns, PFGE detected 10 different patterns, and MLST detected 5 distinct patterns.
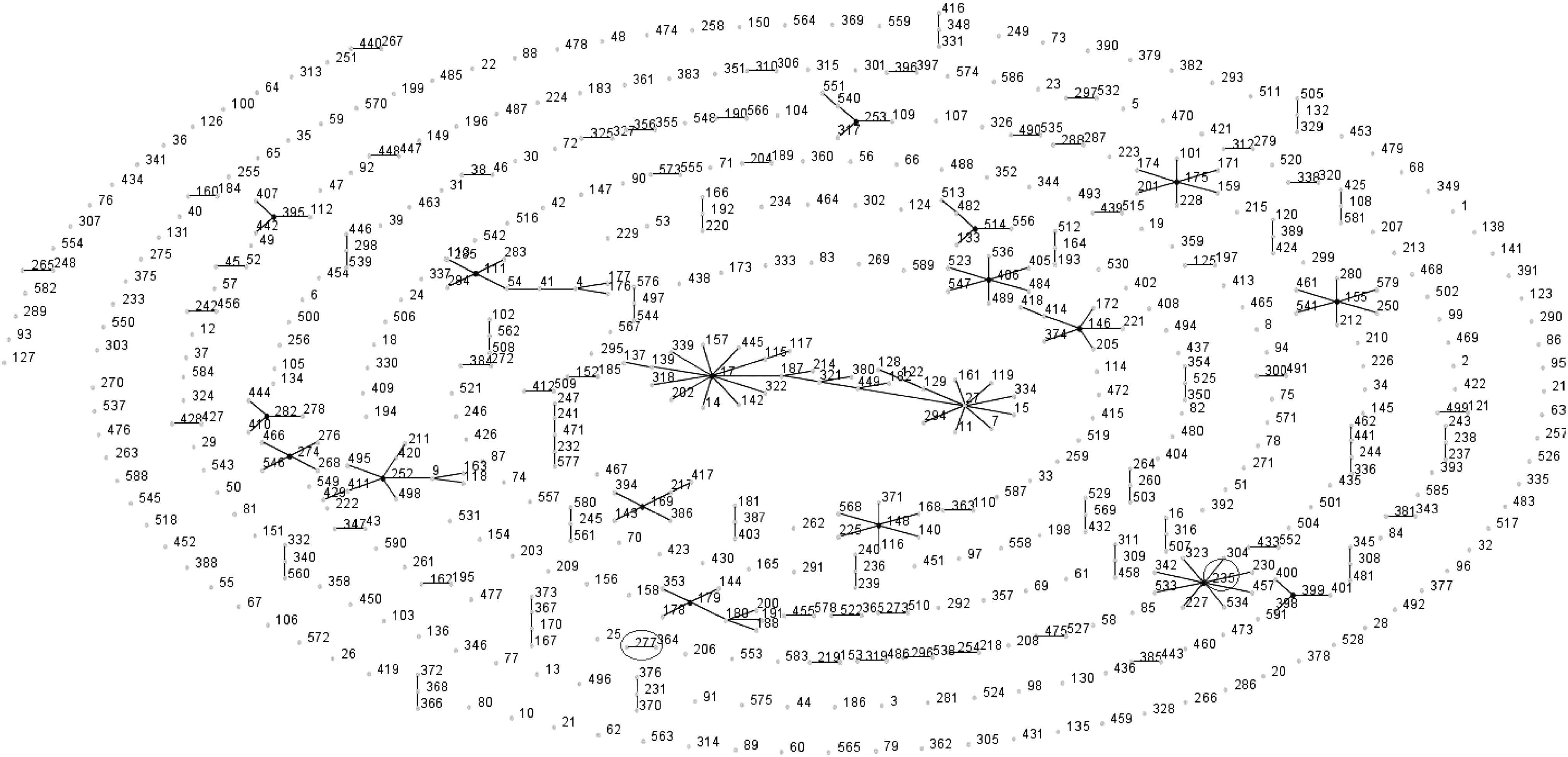
eBURST generated (population snapshot) showing the clusters of linked sequence types (STs) and unlinked STs in the whole Pseudomonas aeruginosa multilocus sequence typing database. ST277 and ST235 found in this study were marked with a circle. The black lines indicate the conection between the sequence types.
Discussion
The molecular typing methods most commonly employed for epidemiological purposes are based on band profile such as PFGE and ribotyping, and are not suitable for studying bacterial population genetics. This is one of the reasons that make sequence-based methods increasingly attractive and potentially effective for this purpose. MLST represents the application of nucleotide sequencing based on the principles previously established by multilocus enzyme electrophoresis. When used for investigation of outbreaks, the MLST shows lower discriminatory power than PFGE or ribotyping because of the clock speed or secular rate of change in the MLST encoding regions is relatively slow. Thus, this technique has been employed as a tool for analyzing the population dynamics of entire bacterial species. In this study, PFGE and ribotyping were able to identify 10 and 13 different patterns, respectively, among all the isolates studied. In contrast, MLST was able to identify only five different patterns among the same isolates. Similar results were observed by Johnson et al., 11 when 90 P. aeruginosa isolates were typed by PFGE and MLST techniques. The results were analyzed using Simpson's D value, and the PFGE technique had a higher discriminatory power than MLST (D value = 0.999 and 0.975, in that order). In contrast, a superior discriminatory power was observed for MLST when different species, Vibrio cholerae and Escherichia coli, were evaluated.13,16
In this study, the 50 SPM-1-producing P. aeruginosa isolates were classified in five distinct PFGE patterns and in 11 different ribogroups. Out of these 50 isolates, 49 were grouped under ST277, showing that they were probably originated from a common ancestor since the housekeeping genes are highly conserved during the evolutionary history. This suggests that the Brazilian SPM-1-producing P. aeruginosa samples are probably derived from a single clone that suffer genetics variations by accumulating mutations along the time. Only one SPM-1-producing P. aeruginosa strain (P3303) isolated from Brazilian southeastern region showed a distinct ST, ST235.
When our MLST results were compared to P. aeruginosa MLST database, it was observed that the ST277 was found in a P. aeruginosa strain isolated for the first time in Vienna. Thus, this strain could have a possible ancestral relationship with SPM-1-producing P. aeruginosa endemic clone. Unfortunately, not enough epidemiological information about this strain was available to establish a relationship between the Austrian and the Brazilian strains. The ST235 observed in the Brazilian strain P3303 producer of SPM-1 was also reported in other 11 P. aeruginosa strains isolated from European Countries, between 2002 and 2005. Empel et al. 6 reported three OXA-10-producing P. aeruginosa isolates grouped under ST235. These isolates showed high PFGE pattern similarity with strains from Turkey that were producers of PER-1, OXA-2, OXA-17, and OXA-74, suggesting, in this way, the possible genetic relationship among these strains.4,5 It would be interesting to evaluate if P. aeruginosa strain P3303 possesses other beta-lactamase encoding genes and has an epidemiologic link with the European strains.
Giske et al. 9 analyzed the genetic relationship among VIM-1-producing P. aeruginosa clones isolated from several European countries by PFGE and MLST. The authors concluded that strains collected from Mediterranean countries were originated from a common ancestral clone. In our study, the three IMP-1-producing P. aeruginosa strains had an identical and new allelic profile, ST593, indicating a common origin. The ST593 had relations with others several samples from different countries, as Russia, Hungary, and Serbia. These ST is related to ST622, recently isolated from Germany. Actually, it is known that the clonal spread of ESβLs and/or MβLs-producing isolates is contributing to spreading of resistance genes.1,15
Our study shows that the majority of SPM-1-producing P. aeruginosa isolates evaluated probably descend from a common ancestor. The same finding was observed among the IMP-producing P. aeruginosa isolates analyzed in this study. The establishment of an epidemiological link between the Brazilian and the European strains deposited on the MLST database could not be asserted since there was not enough epidemiological information available. The present study is pioneer on the insertions of the MLST profiles of P. aeruginosa isolated in Brazil in the MLST database. In addition, the results of this study could help future studies in the understanding of the temporal behavior of the SPM-1-producing P. aeruginosa Brazilian epidemic clone along the time.
Footnotes
Acknowledgments
This study was presented in part at the 48th Interscience Conference on Antimicrobial Agents and Chemotherapy, Washington, DC, October 25–28, 2008.
This study was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo Process Number 05/57496-1. A.C.G. is a researcher from the National Council for Science and Technological Development (CNPq), Ministry of Science and Technology, Brazil (process number: 307816/2009-5).
Disclosure Statement
No competing financial interests exist.