
Abstract
Multidrug-resistant (MDR) Salmonella enterica serovar Typhimurium is the most prevalent penta-resistant serovar isolated from animals by the U.S. National Antimicrobial Resistance Monitoring System. Penta-resistant isolates are often resistant to ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline. To investigate MDR in Salmonella Typhimurium (including variant 5-), one isolate each from cattle, poultry, and swine with at least the ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline phenotype were selected for each year from 1997 to 2007 (n = 33) for microarray analysis of antimicrobial resistance, incompatibility IncA/C, and HI1 plasmid genes. Cluster analysis based on these data separated 31 of the isolates into two groups A and B (15 and 16 isolates, respectively). Isolates in group A were phage type DT104 or U302 and were mostly swine isolates (7/15). Genes detected included intI1, blaPSE-1, floR, aadA, sulI, tet(G), and tetR, which are often found in Salmonella Genomic Island I. Isolates in group B had numerous IncA/C plasmid genes detected and were mostly cattle isolates (9/16). Genes detected included blaCMY-2, floR, aac(3), aadA, aphA1, strA, strB, sulI, sulII, dfrA, dhf, tet(A)(B)(C)(D), and tetR, which are often found on MDR-AmpC IncA/C plasmids. The IncA/C replicon was also detected in all group B isolates. The two remaining isolates did not cluster with any others and both had many HI1 plasmid genes detected. Linkage disequilibrium analysis detected significant associations between plasmid replicon type, phage type, and animal source. These data suggest that MDR in Salmonella Typhimurium is associated with DT104/Salmonella Genomic Island I or IncA/C MDR-AmpC encoding plasmids and these genetic elements have persisted throughout the study period.
Introduction
Genetic elements such as integrons and plasmids are frequently associated with the MDR phenotypes described in Salmonella.45,51 In Salmonella Typhimurium DT104, this includes Salmonella Genomic Island 1 (SGI1), which is a region containing the genes encoding the ACSSuT phenotype on a class I integron. Class I integrons require an integrase gene (intI1) encoding a site-specific recombinase that targets the attI site of the integron and the attC site of the resistance gene cassette.20,40 These MDR-encoding class I integrons are also widespread in other phage types of Salmonella Typhimurium, as well as other serovars of Salmonella and many other Gram-negative bacteria.10,39,49
MDR resistance in Salmonella can also be associated with plasmids, which are small, circular, self-replicating DNA elements, frequently capable of transfer via conjugation.5,41 Horizontal transfer of plasmids to other cells often provides a selective advantage to the organism if the plasmid carries genes encoding antimicrobial resistance, persistence, environmental adaptability, heavy metal resistance, and/or virulence.5,8,28,33 Plasmids are classified by incompatibility (Inc) groups, which are named as such because two members of the same Inc group cannot be stably maintained in a bacterium during cell division. 6 In Salmonella isolated in the United States, IncA/C plasmids have been associated with an expansion of MDR in several serovars of Salmonella. These plasmids are often large (150–200 kb) and some can carry genes encoding for resistance to 10 or more antimicrobials. 21
A recent study demonstrated that MDR in Salmonella Typhimurium veterinary diagnostic isolates were associated with either the DT104 phage type or with IncA/C plasmids. 33 However, because these Salmonella were isolated from clinically ill animals, it is unlikely that they would enter the food chain and be a potential source of foodborne MDR infections. Therefore, this present study was conducted to determine if the association of DT104 and IncA/C with MDR clinical isolates is also found in nonclinical Salmonella Typhimurium isolated from healthy food animals, on-farm or at slaughter. An MDR Salmonella Typhimurium isolate from each major commodity (cattle, chicken, and swine) was selected for each of the first 11 years (1997–2007) of the National Antimicrobial Resistance Monitoring System (NARMS) for genetic analyses (n = 33). Microarray detection of resistance genes and IncA/C and HI1 plasmid genes, as well as polymerase chain reaction (PCR) detection of class I integrons and plasmid replicon types, were used to determine the prevalence of integrons and plasmids and their association with antimicrobial resistance genes in these MDR Salmonella Typhimurium.
Materials and Methods
Isolate selection and antimicrobial susceptibility testing
The NARMS Salmonella isolates for this study were collected from 1997 to 2007. All isolates were subjected to susceptibility testing via the SensititreTM semi-automated antimicrobial susceptibility system (TREK Diagnostic Systems, Inc.) using a custom-made panel including amikacin, gentamicin, kanamycin, streptomycin, ampicillin, amoxicillin-clavulanic acid, ceftiofur, ceftriaxone, cefoxitin, sulfamethoxazole/sulfisoxazole, trimethoprim-sulfamethoxazole, chloramphenicol, ciprofloxacin, nalidixic acid, and tetracycline. Serovar and definitive phage types were determined by the National Veterinary Services Laboratory (Ames). For this study, only on-farm and slaughter Salmonella Typhimurium and Salmonella Typhimurium var. 5- (Copenhagen) isolates were chosen, one each from cattle, chicken, and swine for each year 1997 to 2007. Isolates were initially screened for resistance to ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline, and then selected based on resistance to the greatest number of additional antimicrobials. Thus, a total of 33 maximally resistant isolates were selected representing each food animal (cattle, chicken, and swine) for each of the 11 years.
DNA isolation, labeling, and microarray analysis
Salmonella isolates were grown overnight at 37°C in Luria-Bertani broth, and harvested by centrifugation. DNA was extracted using the GenElute Bacterial Genomic DNA kit (Sigma-Aldrich). Cye dye-labeled dCTP (Amersham), Klenow fragment (New England Biolabs), and random primers were used to label genomic DNA overnight at 37°C as previously described. 22 Labeled DNA was purified using Qiagen PCR clean-up kit (Qiagen).
Microarray design and hybridization
The microarray chip contained probes for 1,267 genes, of which 775 were designed to detect antimicrobial resistance genes found in the National Center for Biotechnology Information (NCBI) database. 23 The remaining 487 probes were designed to detect genes found in IncA/C plasmids from several different bacterial strains and the IncHI1 plasmid from Salmonella Typhi. 34 IncA/C plasmid gene probes on the array are based on sequence from six IncA/C plasmids: Yersinia ruckeri str. YR71 pYR1, Y. pestis biovar Orientalis str. IP275 pIP1202, Photobacterium damselae subsp. piscicida pP990180, S. enterica subsp. enterica serovar Newport str. SL254 pSN254, Photobacterium damselae subsp. piscicida pP91278 and Escherichia coli p1658/97. S. enterica subsp. enterica serovar Typhi strain CT18 plasmid, pHCM1, DNA sequence was used to design probes for the HI1 plasmid. Labeled DNA was hybridized to microarray slides as previously described. 22 Slides were scanned using ScanArray Lite microarray analysis system and ScanArray Express software version 1.1 (Packard BioChip Technologies). Positive hybridizations were determined and scored as previously described. 23 The assay is not always capable of differentiating between alleles due to single nucleotide polymorphisms; for genes of particular interest, PCR was used to define the allele detected by the microarray.
PCR detection of blaOXA-2, blaSHV-1, integrase, and integron genes
PCR analyses for the β-lactamase genes, blaOXA-2 and blaSHV-1, used primers, conditions, and controls previously described by Chen et al. 200410 and Rasheed et al. 199743 for blaOXA-2 and blaSHV-1, respectively. Integrase I (intI1) was amplified using previously described primers and methods. 22 DNA preparation for amplification of gene cassettes within the conserved regions of class I integron was performed using previously described methods. 32 Thermocycler parameters were also previously described 47 followed by examination of PCR products on 2% agarose gel with ethidium bromide.
DNA sequencing
DNA sequencing PCRs contained 10 μl water, 8 μl BigDye Ready Reaction Mix (Applied Biosystems Inc. [ABI]), 1 μl 3.2 pmol primer, and 1 μl template DNA. Product from the conserved sequence integron PCR was used as template and primers were the same as those used for PCR. Thermocycler parameters were rapid thermal ramp to 96°C, 96°C for 1 min; 25 cycles of rapid thermal ramp to 96°C, 96°C for 10 sec, rapid thermal ramp to 50°C, 50°C for 5 sec, rapid thermal ramp to 60°C, 60°C for 4 min; final rapid thermal ramp to 4°C where product was held until purification. All thermal ramps were 1°C per second. Ethanol/EDTA precipitation protocol was used to clean up sequencing reactions before analysis using an Applied Biosystems Inc. 3130xl genetic analyzer (ABI) as described by the manufacturer. PCR products were sequenced using both primers, and sequences were assembled with Geneious bioinformatics software (Geneious). 17 Sequences were compared to published sequences in the NCBI database (www.ncbi.nlm.nih.gov/) using the basic local alignment search tool.
Plasmid replicon typing
Three multiplex PCR primer panels were used to detect the presence of 18 plasmid replicons commonly found in Enterobacteriaceae. Primers were obtained from Eurofins MWG Operon. PCR primers, conditions, and controls were followed as previously described.7,33
Cluster analysis of isolates
Relationships between isolates and associated resistance and plasmid genes were determined by hierarchical cluster analysis of hybridization data using open source CLUSTER 3.0 with Euclidean distances for gene content.14,18 The dendrogram was constructed and viewed using Java TreeView version 1.1.4r3 (http://jtreeview.sourceforge.net). 44
Statistical analysis
Linkage disequilibrium (LD) was calculated as an extension of Fisher's exact probability test on contingency tables 46 as instituted by the program Arlequin. 19 Standard settings were used, 10,000 steps in the Markov chain and 1,000 dememorization steps; calculations of D, D′, and r 2 coefficients were made with a significance level of 0.05.
Results
MDR in NARMS S. enterica animal isolates and selection of Salmonella Typhimurium isolates for analysis
From 1997 to 2007, 14.4% (7,828/54,395) of NARMS Salmonella animal isolates were resistant to at least five antimicrobials often including ACSSuT. S. enterica serovar Typhimurium and Salmonella Typhimurium var. 5- (Copenhagen) combined were the serovars most often isolated from animals that were resistant to five or more classes of antimicrobials at 39.8% (3,116/7,828). For this study, only on farm and slaughter isolates from healthy cattle, chickens, and swine were chosen. One isolate from each animal resistant to ACSSuT and the maximum number of other antimicrobials tested were selected for each year from 1997 to 2007, for a total of 33 isolates (Table 1). In one case an isolate that fit the base antimicrobial resistance profile was not collected; therefore, the Salmonella Typhimurium isolate with the most resistances from that particular animal source (cattle) for 1997 was selected (SMG03). Nineteen of these isolates were Salmonella Typhimurium var. 5- and 15 isolates were phage type DT104. In addition to the base ACSSuT resistance profile, resistance among the isolates includes amoxicillin-clavulanic acid (21/33), amikacin (1/33), gentamicin (8/33), kanamycin (18/33), ceftiofur (18/33), ceftriaxone (1/33), nalidixic acid (3/33), and trimethoprim (6/33) (Table 1).
Serovar 5-, Copenhagen.
Phagetype: NT, nontypable.
Antimicrobials: AMC, amoxicillin-clavulanic acid; AMK, amikacin; AMP, ampicillin; CHL, chloramphenicol; CIP, ciprofloxacin; CRO, ceftriaxone; FOX, cefoxitin; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; SMX, sulfamethoxazole; STR, streptomycin; SXT, trimethoprim-sulfamethoxazole; TCY, tetracycline; TIO, ceftiofur.
Conserved sequences found in Class I integrons used to PCR amplify antimicrobial resistance gene cassettes if present.
Clusters as determined by Euclidean distance for gene content calculated by CLUSTER 3.0 and presented in Fig. 1.
bp, base pair; CS, conserved sequences; PCR, polymerase chain reaction; intI1, integrase I.
Detection of antimicrobial resistance and plasmid genes by microarray analysis
Multiple positive hybridizations to antimicrobial resistance gene probes on the microarrays were detected in all of the tested isolates (Table 2, and Supplementary Table S1; Supplementary Data are available online at www.liebertonline.com/mdr). The most prevalent aminoglycoside resistance genes among all isolates were aac(3) (20/33), aphA1 (10/33), aadA (27/33), strA (19/33), and strB (19/33). β-lactam resistance genes included blaCMY-2 (15/33), blaPSE-1 (18/33), and blaTEM (10/33). Isolates SMG13 and SMG14 were each positive for blaOXA-2, and SMG14 was also positive for blaSHV-1. PCR amplification for each of these genes failed to detect either of them in these isolates (data not shown), suggesting that the microarray results were likely due to cross-hybridization of the probes with other genes. Most prevalent chloramphenicol resistance genes included cat (1/33) and floR (32/33). Sulfamethoxazole resistance genes included sulI (30/33) and sulII (17/33). Tetracycline resistance genes included tet(A) (18/33), tet(A) class D (1/33), tet(B) (5/33), tet(C) (4/33), tet(D) (5/33), tet(G) (16/33), and the regulatory gene tetR (22/33). Trimethoprim resistance genes included dfrA (21/33) and dhf (7/33). In addition to resistance genes, multiple integrase genes, integrons, IS elements, and transposons were detected in all of the isolates (Supplementary Table S1).
Genes and gene families with multiple probes are summarized and presented only once; (-) indicates genes in this table were not detected in the isolate (full hybridization data are available in Supplementary Table S1).
Clusters as determined by Euclidean distance for gene content calculated by CLUSTER 3.0 and presented in Fig. 1.
Positive hybridization to plasmid gene probes indicated the possible presence of IncA/C and H1 plasmids in some isolates (Table 3). Sixteen isolates had a large number of IncA/C plasmid gene detected, including the core genes associated with the IncA/C plasmid backbone (pYR1 genes) and genes associated with the Salmonella Newport MDR AmpC plasmid (pSN254 genes) (Table 3 and Supplementary Table S1). Seven of these 16 isolates had genes detected in all of the 12 regions located around the genome of IncA/C plasmids and previously used to investigate the core backbone of IncA/C plasmids (Table 3). 50 Of the remaining isolates, eight did not have genes detected in regions 6–9 and one lacked genes in regions 7–9, all of the other regions were detected in these isolates (Table 3). Regions 6–9 and 7–9 are within a previously defined insertion/deletion for IncA/C plasmids, and is in the same location as an integrating and conjugative element hot spot 2.34,53 The hybridization patterns for these 16 isolates to genes in the IncA/C core backbone regions were very similar to regions previously reported in IncA/C plasmids from Salmonella Typhimurium, Salmonella Newport, and other Salmonella serovars.34,50 Only two isolates had a large number of HI1 plasmid genes detected, SMG13 with 100/206 genes and SMG14 with 202/206 genes (data not shown; available in Supplementary Table S1).
IncA/C plasmid core regions are based upon the assay by Welch et al. (2007).50 Core regions are numbered in order from 1 to 12 around the positive strand of the IncA/C plasmid sequence. Regions are scored as present if genes within them were detected by microarray analysis of the isolate. These regions are identified with the full hybridization data in Supplementary Table S1.
Clusters as determined by Euclidean distance for gene content calculated by CLUSTER 3.0 and presented in Fig. 1.
Integron intI1 detection and analysis by PCR and sequencing
Microarray analysis detected positive hybridizations to probes for integron genes in 29/33 isolates (Table 2). PCR analysis confirmed the presence of the intI1 gene in the same 29 isolates (Table 1). Integron gene cassettes were analyzed by PCR with primers designed to recognize the conserved regions (CS) of class I integrons, which yielded PCR products in 11 of 29 isolates positive for intI1 (Table 1). PCR amplicon sizes ranged from ∼1,000 to ∼1,400 bp and two isolates yielded two amplicons, one containing 1,100 and 1,200 bp amplicons and the other containing 1,200 and 1,400 bp amplicons. DNA sequencing of integron PCR products was used to identify genes located within the class I integron cassettes. CS PCR products were grouped together based on size and one isolate from each size group was selected for DNA sequencing. The 1,000 bp amplicon from SMG22 was analyzed by basic local alignment search tool analysis, and found to share 92% similarity to its closest match in the NCBI database, aadA12 (Table 1), an aminoglycoside resistance gene previously found in class I integrons of Salmonella animal isolates and human uropathogenic Escherichia coli in the United States. 1 The 1,100 bp product of SMG11 also shared similarity (100%) with aadA12. The 1,200 bp amplicon of SMG05 was 97.7% similar to its closest match in the database, another aminoglycoside resistance gene, aadA2, which is also associated with class I integrons and SGI1 in Salmonella and class I integrons in E. coli isolated from humans in the European Union and Asia. 1 The two PCR products of SMG30 correspond to two different resistance genes. The 1,100 bp band was 98.9% similar to the aminoglycoside resistance gene, aadA2, and the 1,200 bp product was 98.5% similar to the β-lactam resistance gene blaPSE-1, both of which are often found in Salmonella Typhimurium DT104 SGI1. 38
Plasmid replicon analysis
Multiplex PCR detected the presence of plasmid replicons in 30 (90.9%) of 33 isolates (Tables 1 and 3). Nine of the 18 plasmid replicons commonly found in Enterobacteriaceae were detected in these isolates. The most prevalent plasmid replicons detected were FIIA (16/33), A/C (16/33), FIB, (9/33), I1 (7/33), and P (5/33). HI1, N, HI2, and F were each present in one isolate. Only 9 of 18 plasmid groups commonly found in Enterobacteriaceae were detected in these isolates. Four isolates (SMG15, SMG21, SMG22, and SMG27) contained four plasmids, whereas three isolates (SMG09, SMG30, and SMG31) were not positive for any of the plasmid groups tested.
Cluster and LD analysis
The Salmonella Typhimurium isolates were divided into two major groups (Fig. 1A, B) and two single isolates (Fig. 1C, D) by cluster analysis based on resistance and plasmid gene probe hybridizations. Pairwise LD showed significant linkage (p value of ≤0.05) between several pairs of plasmid replicon types, phage types, and animal sources that often fell into separate cluster analysis groups (Figs. 1 and 2). The 15 isolates of hybridization cluster A had a large number of resistance and integron genes detected, but very few IncA/C or HI1 genes were detected (Table 2, Fig. 1 and Supplementary Fig. S1). All 15 members of cluster A were positive for the intI1 gene by PCR. Additionally, six isolates in cluster A had insertion sequences detected by PCR, two of which were sequenced and found to have resistance genes aadA2 or aadA2 plus blaPSE-1 inserted into the conserved sequence region of IntI1, which is indicative of SGI1. 38 Moreover, all isolates in group A were phage type DT104 (n = 11), DT104A (1), DT104B (1), or U302 (2), all of which have been often found to have a class I integron inserted into SGI1. 38 All but three of the isolates had FIIA plasmid replicons detected by PCR; two of these were also positive for I1, and no other replicons were detected in isolates of cluster A. LD analysis also showed significant linkage between phage type and replicon type FIIA, 12 out of 15 isolates in cluster A were positive for FIIA and all but two of these were DT104, DT104A, or DT104B (Figs. 1 and 2).
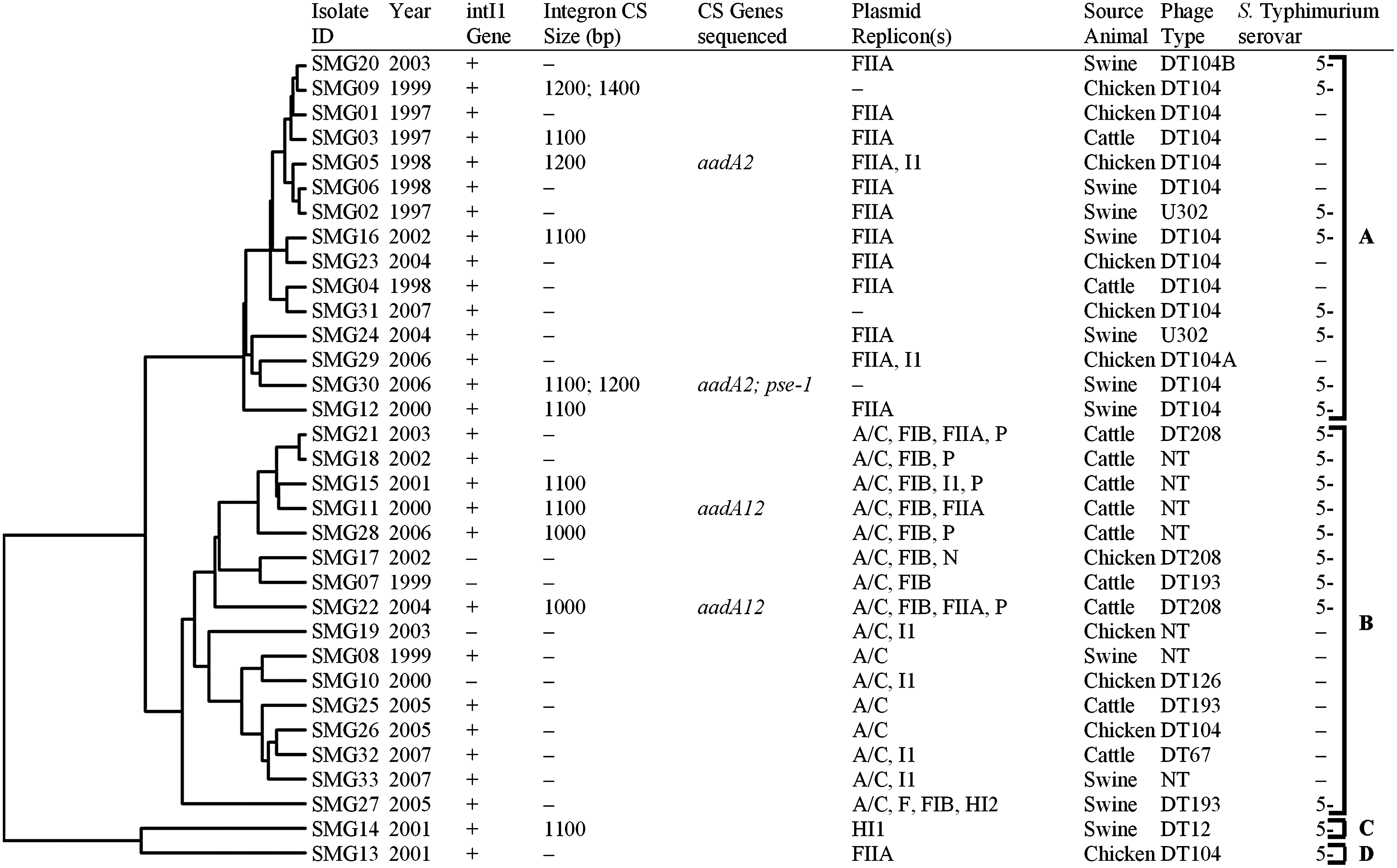
Cluster analysis of Salmonella Typhimurium isolates based on results of microarray detection of antimicrobial resistance and plasmid genes. Complete cluster analysis data are available in Supplementary Fig. S1. NT, not typable.

Pairwise LD based on nine plasmid replicon groups, serovar, phage type, and animal source of Salmonella Typhimurium strains. (+) indicates a p-value ≤0.05 or less and significant LD; and a (-) indicates a p-value >0.05 and no significant LD. Replicon groups B/O, FIC, T, K, W, Y, FIA, X, and L/M were not present in any isolate and were subsequently removed from the analysis. LD, linkage disequilibrium.
The IncA/C replicon was detected by PCR in all 16 isolates of cluster B and microarray analysis detected a large numbers of IncA/C plasmid genes in these isolates, including those identified as core IncA/C genes. 50 LD analysis detected significant disequilibrium between IncA/C, paired with phage type, and animal source. The isolates in cluster B were from a variety of phage types, with seven being not typable and only one being DT104 (Fig. 2). The dominant animal source in cluster B was cattle (9/15). Several isolates in cluster B also had other replicons detected, including FIB (n = 9), I1 (5), P (5), FIIA (3), and N (1) (Fig. 1). Significant LD was detected for all of these except I1, which is also found in cluster A (n = 2) (Figs. 1 and 2). Twelve isolates from cluster B were intI1 gene positive, four of which also yielded PCR products from the IntI1 conserved region, two of which were sequenced and identified as an aadA12 gene usually associated with class I integrons that can be located on plasmids (Fig. 1).1,52
The last two isolates, C and D, each formed a lone branch on the dendrogram (Fig. 1). Isolate C, SMG14, was DT12, had intI1 detected, a 1,100 bp cassette region, and one plasmid replicon type detected, HI1 (Fig. 1). This isolate grouped alone due to many more HI1 hybridizations than any other isolates (n = 202/206), but it also had many IncA/C gene detected (Fig. 1 and Table 3) despite not having an IncA/C replicon detected by PCR. Significant LD was detected between replicon type HI2 and F, which were found only in this isolate (Figs. 1 and 2). Isolate D, SMG13, also had a large number of HI1 hybridizations (100/206), although the HI1 replicon type was not detected and only the FIIA replicon type was detected (Fig. 1 and Table 3).
Discussion
This research was conducted to identify antimicrobial resistance associated genetic elements found in Salmonella Typhimurium, one of the most prevalent MDR serovars of S. enterica recovered from humans and animals. We used a newly developed DNA microarray to identify antimicrobial resistance genes, mobilizable elements, and genes found in IncA/C and HI1 plasmids known to carry MDR in Salmonella.23,34 Some of the genetic elements, plasmids, integrons, and resistance genes were also investigated by PCR and sequence analysis. As a result of the extensive nature of the analyses, only 33 Salmonella Typhimurium isolates were included in this study. To increase the likelihood of detecting as many resistance-associated elements as possible, those isolates with resistance to the greatest number of antimicrobials were selected. Interestingly, 31 of these isolates were split almost evenly between the two genetic elements previously found to be associated with Salmonella Typhimurium MDR: DT104/SGI1 (15 isolates) and IncA/C plasmids (16 isolates).
Due to the small sampling of isolates, care must be taken not to over interpret the data; however, some conclusions can be made. Although only 11 isolates per animal source were studied, it is interesting that isolates in the IncA/C-positive cluster were mostly from cattle (9/16). Significant LD was detected between animal source and IncA/C, indicating that these associations are not by chance. The association of IncA/C with MDR in cattle is interesting because numerous studies have shown that MDR in Salmonella isolated from U.S. cattle is often due to an IncA/C plasmid that encodes the MDR-AmpC phenotype, which includes resistance to third-generation cephalosporins.16,26,27,42,48,54 These plasmids are often found in MDR Salmonella Newport,16,26,27,42,48,54 which is isolated most frequently from cattle than from other sources (www.fda.gov/AnimalVeterinary/SafetyHealth/AntimicrobialResistance/NationalAntimicrobialResistanceMonitoringSystem/ucm209502.htm). Although it has been reported that the MDR-AmpC phenotype seems to be spread by clonal expansion of MDR isolates, horizontal exchange of the resistances are also possible.26,27 It was previously shown by PFGE analysis that in the distant past, Salmonella Typhimurium likely had an MDR IncA/C plasmid and that these MDR genes were transferred from the plasmid to the chromosome in DT104 followed by loss of the plasmid. 33 However, in Salmonella Newport it is plausible that the MDR IncA/C plasmid was acquired, perhaps recently, by horizontal exchange. 29 Therefore, it is difficult to say if the IncA/C plasmids in MDR Salmonella Typhimurium and MDR Salmonella Newport found in cattle could have originated from an MDR serovar and then transferred to a susceptible serovar, or if the cattle environment selected for clones of both serovars Newport and Typhimurium that happen to have similar MDR-AmpC encoding IncA/C plasmids.
Significant LD was also detected for IncA/C and phage type; only one DT104 isolate (SMG26) had an IncA/C replicon detected, and thus IncA/C is underrepresented in DT104 isolates. Therefore, cluster A, dominated by DT104, is significantly different from the other isolates. In cluster A, seven isolates were from swine, five of which were phage type DT104 and two were the closely related U302. Several previous studies have also found that MDR Salmonella Typhimurium DT104 is associated with swine.24,51 Plasmid replicon type FIIA also had significant LD to phage type, and 11 of 15 DT104 isolates were positive for FIIA. The resistance genes found in cluster A DT104 isolates could be located in SGI1 particularly since all of these isolates were intI1 positive, and six out of 15 were PCR positive for insertions at the class I integron conserved sequences. Alternately, the resistance genes, and even resistance genes in integrons, could be located elsewhere in the genome or on an FIIA, I1, replicon type plasmids, or other undetected plasmids.15,35,37
Emerging in 1996, ACSSuT Salmonella Typhimurium DT104 was the first recorded major outbreak of an MDR Salmonella clone in the United States. 25 Prevalence of DT104 peaked in the late 1990s and then decreased slightly from 2000 to 2007 (www.ars.usda.gov/Main/docs.htm?docid=17724). Interestingly, all six of the isolates from the first 2 years of the current study (1997 and 1998) are in the DT104-associated cluster A. Conversely, the IncA/C-associated cluster B has isolates collected mostly from the later years of the study. This may reflect the emergence and decline of DT104 MDR and the increase of IncA/C MDR in Salmonella Typhimurium throughout the study period.
The microarray hybridizations and antimicrobial resistance genes detected are similar every year and for every source, indicating that the genes causing MDR in Salmonella Typhimurium have been fairly stable in the two major MDR genotypes over the 11 years period of the study. The prototypical SGI1 integron found in DT104 has blaPSE-1, floR, aadA2, sulI, dfrA10, tet(G), and tetR; however, the gene content can vary greatly between isolates resulting in multiple subtypes of SGI1 (named SGI1-A, -B, -C, etc.). 38 The isolates of cluster A hybridized most often to probes for the genes found in SGI1. Therefore, data from this study indicate that DT104 ACSSuT, occasionally with additional resistance genes, is predominantly responsible for MDR in cluster A and that the genes detected have been largely stable in these animal isolates over the 11-year period. It is also important to make note that some of the MDR genes detected in DT104 SGI1 are very similar to the genes found on the IncA/C plasmids reflecting a possible common origin.21,33
Resistance genes detected in cluster B were genes often associated with the MDR-AmpC IncA/C plasmids. These genes included blaCMY-2, floR, aac(3), aadA, aphA1, strA, strB, sulI, sulII, dfrA, dhf, tet(A), tet(B), tet(C), tet(D), and tetR, all of which were detected in different combinations that could suggest variations in the IncA/C plasmid's complement of antimicrobial resistance genes. 21 Most of these genes have been previously reported as encoded by the IncA/C plasmids found in several Salmonella serovars and other Gram-negative bacteria.21,50 Sequencing of the conserved integron region of two isolates from this cluster identified the aadA12 gene that has been associated with class I integrons and could be located on an IncA/C or other plasmids. 1 All of the isolates in cluster B are resistant to the third-generation cephalosporin, ceftiofur, and have the blaCMY-2 gene (encoding an AmpC homolog) detected, which is characteristic of both the MDR-AmpC phenotype and the MDR IncA/C plasmid genotype. The spread of these plasmids in Typhimurium and other Salmonella serotypes such as Newport, Heidelberg, and Agona has made investigations of these plasmids a necessity for human health, animal health, and for understanding zoonotic transmission of MDR Salmonella. It has been proposed that use of ceftiofur in animals may have led to the development or selection of this MDR plasmid in Salmonella, especially in cattle, where it is used to treat respiratory diseases, mastitis, and lameness. 36 However, a recent study failed to find a significant correlation between ceftiofur use in cattle and transmission or selection for blaCMY-2-encoding plasmids in Salmonella. 12
The resistance genes detected in the remaining two isolates had similarities to both clusters A and B. In isolate D, isolate SMG13, a DT104 phage type, had blaPSE-1, floR, aac(3), aadA, dfrA, sulI, tet(G), and tet(R) detected similar to the DT104 cluster; however, blaOXA-2, strA, strB, dhf, and tet(A) genes were also detected. Isolate C, SMG14, also had hybridizations similar to SMG13; however, in addition to the blaOXA-2 gene, blaSHV-1, blaTEM-1, and catA were also detected. The blaTEM-1 gene is often found on HI1 plasmids as is catI, a homolog of catA that could be detected by the catA gene probe. In addition, isolate SMG13 was PCR and microarray (202/206) positive for the HI1 plasmid. Interestingly, a large number of IncA/C genes were also detected; however, the IncA/C replicon was not detected in this isolate; therefore, these genes could be located on a chimera of IncA/C and HI1, or could be located elsewhere in the genome. Because blaOXA-2 and blaSHV-1 have not been reported in U.S. Salmonella animal isolates, SMG14 and SMG13 were analyzed by PCR to determine if these were in fact present; however, the assays failed to detect either gene. Future investigations will have to be conducted to establish the prevalence of these types of resistances and determine whether these genetic elements play a major role in MDR of Salmonella Typhimurium or other serovars.
This study determined that DT104/U302-related class I integrons and IncA/C plasmids were two major forms of MDR present in Salmonella Typhimurium isolated from healthy food animals in the United States from 1997 to 2007. These results are similar to our previous study of 2005 Salmonella clinical veterinary isolates. 33 Cluster analysis based on microarray results divided all but two of the isolates into an IncA/C-positive group and an IncA/C-negative group, which was positive for DT104, DT104A, DT104B, or U302 phage types often associated with MDR encoded in SGI1. LD analysis of plasmid replicon types, phage type, and animal source confirmed that these associations and others had significant LD (p-value of <0.05). This study also demonstrated that many similar resistance and integron genes are detected in MDR Salmonella Typhimurium regardless of whether they are associated with DT104 or IncA/C plasmids, and that many of the resistance genes detected in MDR Salmonella Typhimurium did not change from 1997 to 2007 in these NARMS animal isolates.
Disclaimer
The mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Footnotes
Acknowledgments
The authors thank Dr. Jennifer Turpin, Jovita Haro, Dr. Sutawee Thitaram, Benny Barrett, and Takiyah Ball for technical assistance.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.