
Abstract
The mef(A) gene codes for an efflux protein that conveys resistance to 14- and 15-membered macrolides. Enterococci are emerging pathogens, as well as indicator and reservoir bacteria that are known to have a strong tendency to acquire resistance genes. A total of 485 Enterococcus faecalis strains of porcine (n = 239) and human origin (n = 246) were screened for the presence of the mef(A) gene by using polymerase chain reaction. In total, 29 E. faecalis of porcine (n = 10) and human (n = 19) origin were positive for the presence of the mef(A) gene. Most of the mef(A)-containing strains were isolated from fecal samples of healthy individuals; only one strain originated from a stool sample of a diseased pig. To our knowledge, this is the first report on the occurrence of the mef(A) gene in E. faecalis apart from mating experiments. The main clinical relevance of this study is that donor E. faecalis might transfer the mef(A) gene to recipients that are usually combated with macrolides. Hence, the role of E. faecalis as a resistance reservoir with respect to limited treatment options are a cause for concern.
Introduction
Materials and Methods
Between years 2002 and 2008, a total of 485 E. faecalis strains from Southern Germany were, after isolation and identification as previously described, 26 screened for the presence of the mef(A) gene. Of these, 239 were of porcine and 246 of human origin: 147 porcine and 98 human strains originated from manure/stool samples of individuals without clinical signs (“commensal” isolates), whereas 92 of the porcine and 148 of the human E. faecalis were mainly isolated from clinically indicated stool samples, wound swabs, or urine samples of hospitalized patients. The latter need not necessarily be the causative agents of infection. Nevertheless, we designate them as “clinical” isolates in this study, because systemic antibiotic therapy has an effect on all bacteria, regardless whether they are the cause of disease or only commensals.
Phenotypic resistance characteristics were tested by the broth microdilution method according to DIN 58940-81 6 and as previously described. 26 After DNA extraction with the QIAquick PCR Purification Kit (Qiagen, Hilden, Germany), a 348-bp product of the mef(A) gene was detected by polymerase chain reaction (PCR) using specific primers (upstream, 5′-AGT ATC ATTAAT CAC TAG TGC-3′; downstream, 5′-TTC TTC TGG TAC TAA AAG TGG-3′), followed by agarose gel electrophoresis as described elsewhere. 2 Additionally, all isolates were screened for the presence of the erythromycin resistance genes erm(A) and erm(B), as described elsewhere.12,18 Each PCR cycle contained one positive control [mef(A): Streptococcus pyogenes, RRR 334; erm(A): S. pyogenes, RRR 334; erm(B): E. faecalis, UW3553-1, and one negative control (master mix without template DNA)]. Of the mef(A)-positive isolates, four PCR products were sent for sequencing (Sequiserve, Vaterstetten, Germany). Sequence similarity was compared using NCBI BLAST database (www.ncbi.nlm.nih.gov/). Restriction fragment length polymorphism analysis of the mef(A)-positive isolates was conducted with the restriction endonuclease HindIII (Roche, Mannheim, Germany), according to manufacturer's instructions. E. faecalis DSM 2570 and Enterococcus faecium DSM 20477 were used as control strains. Restriction fragments were size-separated on EtBr-containing agarose gel (0.8%) and observed using UV light. The DNA Molecular Weight Marker II (Roche) was used for size estimation of the fragments. Cluster analysis was performed using the GelCompar Software V. 5.10 (Applied Maths, Sint-Martens-Latem, Belgium).
Results
In total, 29 E. faecalis were positive for the presence of the mef(A) gene in the PCR analysis. Hereof, 9 (6.1%) porcine and 19 (19.4%) human strains were commensal isolates. On the contrary, only one clinical strain, isolated from a fecal sample of porcine origin, was a carrier of the mef(A) gene. The diseased pig was from a separate farm without contact to the shedders of the commensal mef(A)-strains. Two of the nine commensal porcine strains were isolated from the same manure sample; the remaining seven strains originated from manure samples of different and geographically separated farms. All positive strains of human origin were obtained from different persons, mostly from different administrative districts. Phenotypic and genotypic resistance properties of all investigated bacteria are provided in Table 1. Most of the intermediate or resistant isolates without mef(A) were erm(B)-carriers. The nucleotide sequences of the sequenced amplicons were identical to each other and exactly matched a section of the “Streptococcus pyogenes macrolide efflux protein A (mefA),” GI/18478326. Pearson correlation coefficients obtained in a GelCompar-anlysis of the HindIII-restricted isolates were between 71% and 99%. The highest similarities were found between two isolates of healthy humans. Direct contact between these two persons is unlikely because the samples came from widely separated regions. The two strains isolated from the manure sample had only 78% similarity in the pairwise comparison. Similarities between samples and reference strain E. faecalis DSM 2570 were between 80% and 97%. The E. faecium DSM 20477 strain was clearly distinct from E. faecalis strains with similarity values between 52% and 83%. One exemplary cluster analysis is shown in Fig. 1.
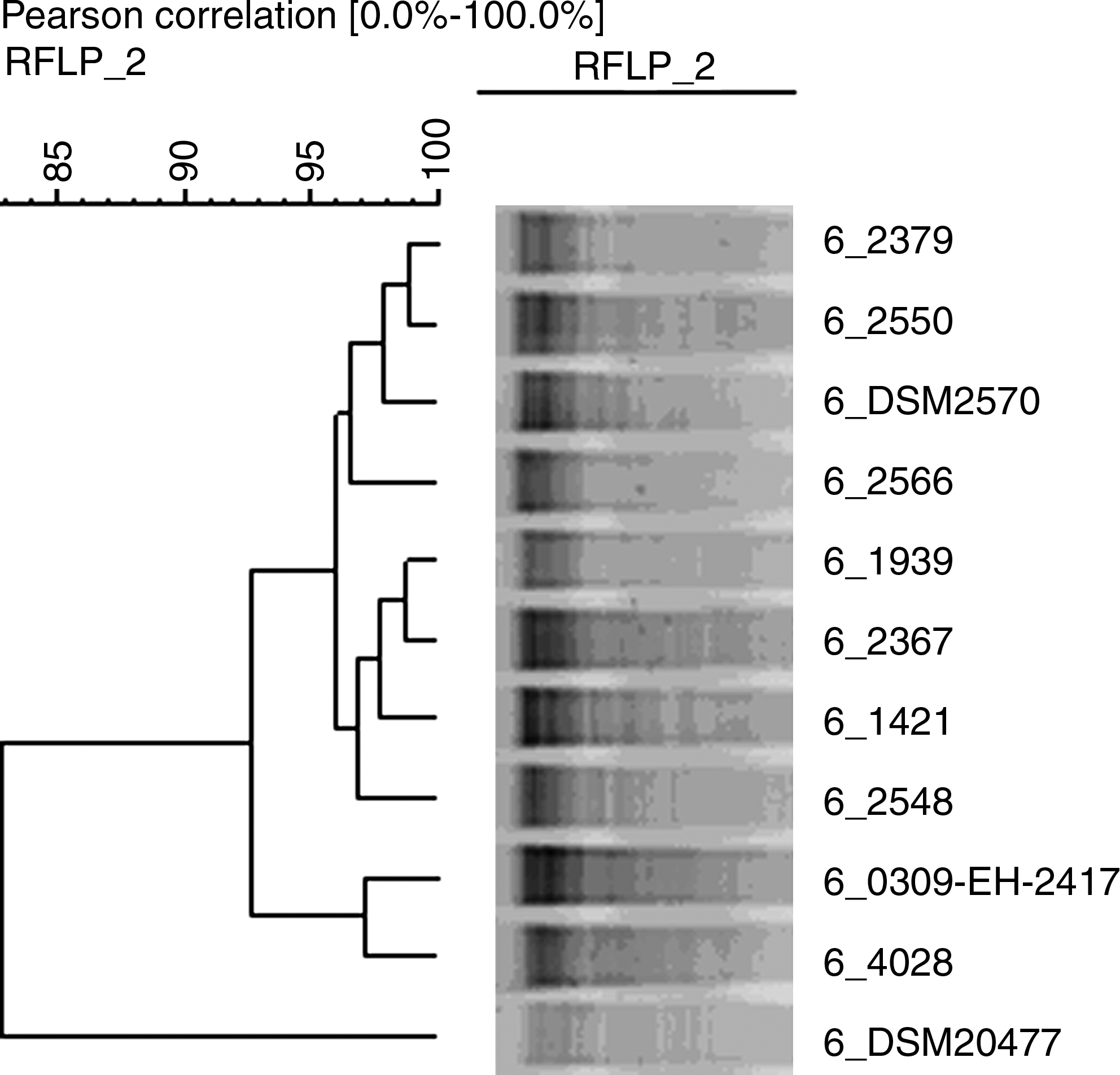
Exemplary RFLP patterns and cluster analysis from HindIII digested mef(A)-positive Enterococcus faecalis. Reference strains: E. faecalis DSM 2570 (6_DSM2570); Enterococcus faecium DSM 20477 (6_DSM20477); porcine isolates: 6_4028, 6_1421, 6_1939; human isolates: 6_2379, 6_2550, 6_2566, 6_2367, 6_2548, and 6_0309-EH-2417. RFLP, restriction fragment length polymorphism.
Hereof, n = 93/35/62/18 contained erm(B); n = 2/0/15/0 contained erm(A) + erm(B).
Hereof, n = 6/31/7/102 contained erm(B); n = 0/1/1/1 contained erm(A) + erm(B).
Hereof, n = 1/8/2/2 contained erm(B); n = 0/0/3/1 contained erm(A) + erm(B).
MIC, minimal inhibitory concentration.
Discussion
Petsaris et al. 21 detected a mef(A)-like gene in another Enterococcus species, particularly in a commensal E. faecium strain originating from a chicken cecum sample. Sapkota et al. 25 found the mef(A) gene in not differentiated Enterococcus spp., but not explicitly in E. faecalis. About 10 years ago, Luna et al.15,17 detected a gene in Enterococcus spp. that was initially described as mef(A), but due to sequence differences (the resulting protein products share only 88% identity) it has been subsequently renamed to mef(E). 13 It is well known that enterococci have a strong tendency to acquire new antimicrobial resistances.10,14,28 It is also known that the mef(A) gene can be transferred from a donor to an E. faecalis recipient.4,16,19,27 However, apart from these mating experiments, mef(A) has, to our knowledge, not been detected in wild-type E. faecalis to date, even if some scientists report on research on this topic.5,7,22,23 Compared to the above-mentioned studies, the mef(A) prevalence in the present study seems very high. Whether these findings are due to regional disparities has yet to be explored. Restriction fragment length polymorphism patterns reveal a high similarity of few strains. In these cases, a clonal spread cannot be excluded. However, certainly not all mef(A)-positive isolates result from clonal spread of one strain, since many isolates were less similar to each other than to the reference strain E. faecalis DSM 2570 (Fig. 1). There are four mef(A) carriers with susceptible phenotypes; three of them additionally harbor the erm(B) gene (Table 1). Phenotypically susceptible resistance gene carriers might result from mutations in the respective gene. Additionally, we cannot rule out that the susceptible phenotype is caused by an intact, but silent resistance gene. Further investigations would be necessary to determine antibiotic resistance gene silencing as described by Enne and Bennett. 9 It is remarkable that almost exclusively the commensal strains carry the mef(A) gene. The reason in this particular case has yet to be discovered too, but it is conceivable that clinical isolates have acquired virulence genes at the expense or instead of ballast antimicrobial resistance, since carrying mobile genetic elements is at the expense of viability. 8 This theory is supported by the fact that recent studies targeting (and not finding) the mef(A) gene in E. faecalis investigated only clinical strains. Interestingly, the mef(E) [formerly designated as mef(A)] containing enterococci described by Luna et al. 17 were also commensal strains, isolated from healthy children. However, the results are still of clinical relevance, since donor E. faecalis are able to transfer the mef gene. 15 This is of particular interest when the recipients are usually combated with macrolides, for example, pneumococci or streptococci. 11 Hence, the role of E. faecalis as a resistance reservoir with respect to limited treatment options are a cause for concern.
Footnotes
Acknowledgments
This work was supported by the Bavarian State Ministry of the Environment, Public Health and Consumer Protection and by the Bavarian State Ministry of Agriculture and Forestry. We are grateful to S. Hörmansdorfer and G. Mölle (Bavarian Health and Food Safety Authority, LGL) for providing the clinical isolates. Thanks also go to R.R. Reinert (Nationales Referenzzentrum für Streptokokken, Universitätsklinikum Aachen, Institut für Med. Mikrobiologie), as well as to I. Klare (Robert Koch Institut, RKI) and assistants for kindly providing the reference strains.
Disclosure Statement
No competing financial interests exist.