
Abstract
Introduction
PZA requires an acidic environment to function, making resistance testing difficult. 5 As such, it has often been excluded from efforts to develop tools to detect drug resistance. 2 Since testing is not routinely performed, the prevalence of PZA resistance is not well known; however, surveillance studies have reported prevalence of 52%–78% in treatment failure or MDR-TB cases, and 2%–10% in non-MDR-TB cases.1,7,9,10
PZA is a pro-drug that enters bacteria by passive diffusion and is converted to its active form, pyrazinoic acid (POA), by bacterial pyrazinamidase (PZAse).6,12,19 POA is expelled from the mycobacterium by an efflux pump, it is protonated in the acidic extracellular space, and the protonated form is then reabsorbed into the bacilli, where the proton is released. This cycle is repeated, resulting in a lethal combination of POA accumulation and intracellular acidification, causing disruption of membrane permeability and cellular damage.3,17,19
The major mechanism of PZA resistance in M. tuberculosis arises from mutations in the PZAse, resulting in a loss of enzymatic activity, preventing conversion of PZA to POA. 17 The same mechanism is responsible for the natural resistance to PZA of Mycobacterium bovis and Mycobacterium kansasii.6,12,15,18 However, these mutations cannot fully account for M. tuberculosis PZA resistance. Variations in the rate of bacterial POA expulsion (POA efflux) are implicated in PZA resistance15,18,20; these variations may depend on PZAse activity levels, intracellular PZAse concentrations, and POA efflux pump efficiency. Mycobacterium smegmatis, naturally PZA resistant, has a much more active POA efflux mechanism than M. tuberculosis, which prevents intracellular POA accumulation.15,18,19
M. tuberculosis strains exhibit a range of POA efflux rates, associated with different PZA-susceptibility levels. Recently, POA efflux rates from susceptible M. tuberculosis strains were found to fall within a critical range, with strains with rates below this range showing resistance. 20 The POA efflux rate accounted for 61% of the variability in BACTEC460 PZA susceptibility and 51% of the BACTEC460 growth index. POA efflux rates were able to account for differences in BACTEC460 susceptibility to a greater extent than the detection of PZAse mutations.
Based on these results, it was hypothesized that measurement of the POA efflux rate could lead to the development of a novel test for PZA resistance. The present study aims at evaluating the accuracy of POA efflux rate as a predictor of PZA resistance as determined by BACTEC460.
Materials and Methods
Thirty-four M. tuberculosis clinical isolates, including the PZA-susceptible reference strain H37Rv, were analyzed in a previous study 13 from which data concerning PZA susceptibility based on BACTEC460 were available.
BACTEC460 was performed at pH 6.0 using 100 μg/ml PZA. The resistance level was estimated using the ratio of the radioactive growth index of the culture media containing bacteria and PZA to the radioactive growth index of the control (media without PZA), expressed as a percentage (Table 1). PZA-resistant strains were classified according to the BACTEC growth index. Two classifications were used for considering PZA resistance: a growth index greater than 11% (Bactec11) and greater than 20% (Bactec20).
ND, nondetermine; POA, pyrazinoic acid.
In a previous study, 20 the POA efflux rate was measured for each strain. Briefly, bacteria were incubated at 37°C for 12, 24, 36, and 48 hr with 1 mM of PZA. Samples were centrifuged, and the supernatants were recovered. POA in the extracellular environment was quantified by Wayne reaction and fitted to a linear regression in time. The POA efflux rate was estimated using the readings until 48 hr [VPOA(48h)]. We extended the previous study by estimating the POA efflux rate using only the reading at 12 hr (Table 1).
BACTEC460 susceptibility (BACTEC11 and BACTEC20) was modeled in a logistic regression with the POA efflux rate [VPOA(48h) and VPOA(12h)] as the predictor covariate. Sensitivity and specificity were estimated for different probability cutoffs.
Results
The POA-efflux rate estimated with a single reading at 12 hr [VPOA(12h)] was highly correlated with the corresponding POA-efflux rate estimated with readings at 12, 24, 36, and 48 hr [VPOA(48h)], with a Pearson correlation coefficient of 0.94 (p<0.0001) (Fig. 1).
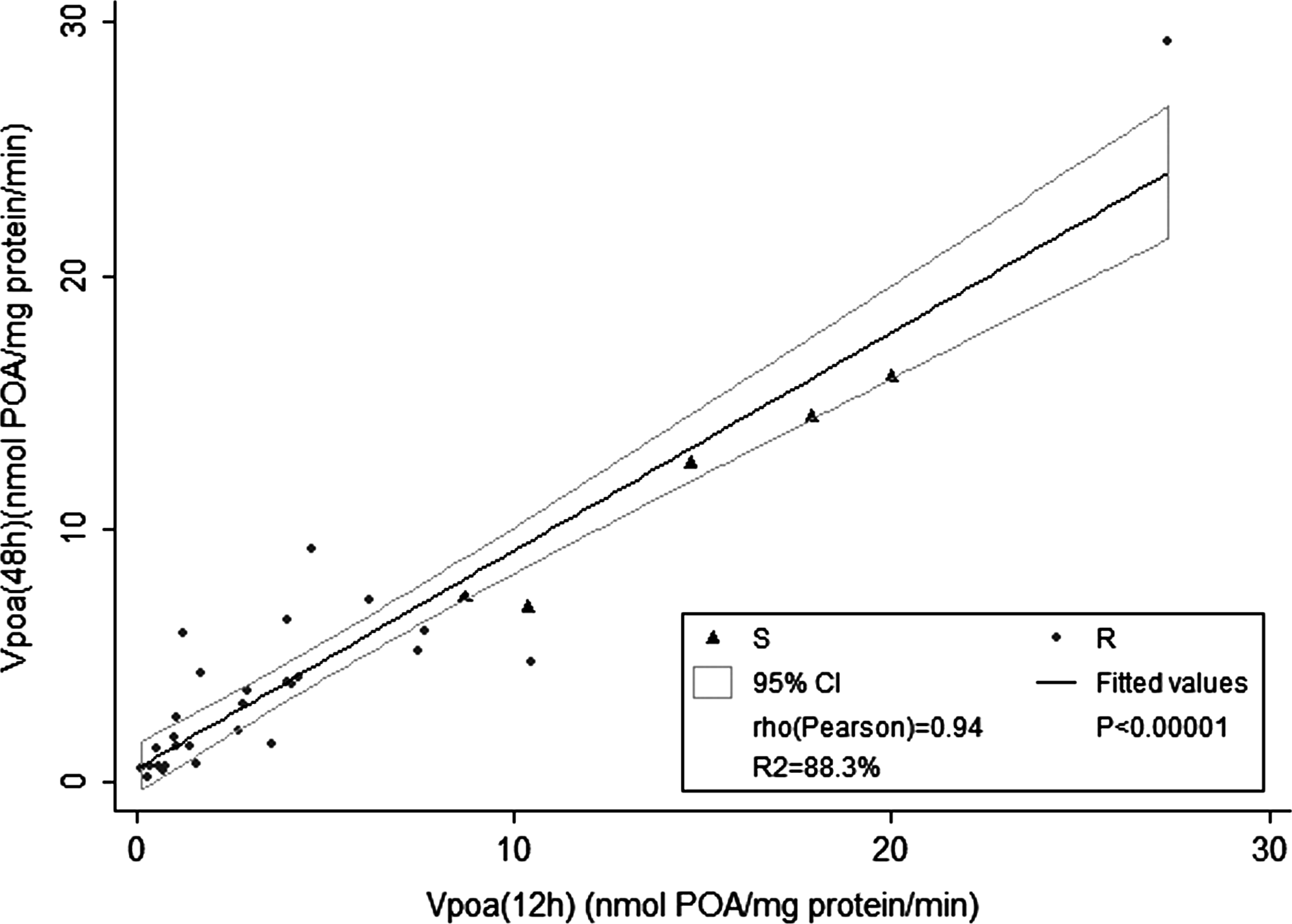
Correlation between the pyrazinoic acid (POA) efflux rate estimated from a 48-hr culture, VPOA(48h) (0, 12, 24, 36, and 48 hr), and estimated from a 12-hr culture, VPOA(12h) (0, 12 hr).
The POA efflux rate at 12 hr was significantly associated with BACTEC460 susceptibility (OR=0.80, p=0.024). For the growth index >11%, the highest specificity, 100%, was associated with 89.7% sensitivity for VPOA(48h) and 92.9% for VPOA(12h). For the growth index >20%, the highest specificity, 100%, was associated with 70.8% sensitivity for VPOA(48h) and 78.3% for VPOA(12h) (Table 2). The area under the receiver-operating curve (ROC) ranged in all cases from 0.92 to 0.95. Sensitivity and specificity to determine PZA resistance based on the POA efflux rate for different cutoffs of the predicted probability are shown in the nonparametric sensitivity-specificity curves in Fig. 2.

Sensitivity and specificity that determine pyrazinamide resistance based on the POA efflux rate for different cutoffs of the predicted probability.
ROC, receiver-operating curve.
Discussion
The analysis performed in this study showed that the POA efflux rate was able to predict PZA resistance with high specificity and sensitivity. A 7% of true resistant strains are not detected when using the growth index cutoff of 11%, while a 22% is missing if using a 20% cutoff.
PZA plays a key role in TB control. However, although PZA resistance is common in some countries, 16 the lack of reliable culture-based resistance tests and the high cost and modest accuracy of molecular methods means that testing is not generally performed. Consequently, patients with PZA-resistant TB are usually treated inappropriately with this hepatotoxic drug. The frequency and significance of undetected PZA resistance to the success of treatment regimens is unknown.
This study confirms the “proof of concept” that the quantitative measurement of POA accumulation is an important predictor of PZA resistance, although the direct measurement of extracellular POA accumulation with a protocol similar to the one described in this article may not be practical for routine testing given the demanding amount of the cultured strain required.
However, the evidence found in this study suggests that it could be feasible to develop a novel approach and a simple assay for testing PZA resistance. We are currently in the process of developing rapid immunoassays for measuring POA levels, and this will be the subject of a future article.
The development of a reliable tool for detecting PZA resistance is likely to have an impact on the clinical outcomes of individuals and populations, and the cost of providing TB control services. If widely implemented, this could have significant public health implications.
Footnotes
Acknowledgments
This research was funded by the National Institute of Allergy and Infectious Diseases, National Institutes of Health US, under the terms of Award 1R01TW008669-01, TDR-WHO Reference 2009/53662-0, TWAS 08-070RG/BIO/LA-UNESCO FR:3240204464. PS and MZ were supported by TMRC New Tools to Understand and Control Endemic Parasites # 1 P01 AI51976 and Global Research Training Grant # 3 D43 TW006581.
Disclosure Statement
No competing financial interests exist.