
Abstract
The emergence of Escherichia coli resistant to extended-spectrum cephalosporins (ESCs) is of concern as ESC is often used to treat infections by Gram-negative bacteria. One-hundred and ten E. coli strains isolated in 2009–2010 from children warded in a Malaysian tertiary hospital were analyzed for their antibiograms, carriage of extended-spectrum beta-lactamase (ESBL) and AmpC genes, possible inclusion of the beta-lactamase genes on an integron platform, and their genetic relatedness. All E. coli strains were sensitive to carbapenems. About 46% of strains were multidrug resistant (MDR; i.e., resistant to ≥3 antibiotic classes) and almost half (45%) were nonsusceptible to ESCs. Among the MDR strains, high resistance rates were observed for ampicillin (98%), tetracycline (75%), and trimethoprim/sulfamethoxazole (73%). Out of 110 strains, blaTEM-1 (49.1%), blaCTX-M (11.8%), and blaCMY-2 (6.4%) were detected. Twenty-one strains were ESBL producers. CTX-M-15 was the predominant CTX-M variant found and this is the first report of a CTX-M-27-producing E. coli strain from Malaysia. Majority (3.1%) of the strains harbored class 1 integron-encoded integrases with a predominance of aadA and dfr genes within the integron variable region. No gene cassette encoding ESBL genes was found and integrons were not significantly associated with ESBL or non-ESBL producers. Possible clonal expansion was observed for few CTX-M-15-positive strains but the O25-ST131 E. coli clone known to harbor CTX-M-15 was not detected while CMY-2-positive strains were genetically diverse.
Introduction
There have been very few reports on the prevalence of beta-lactamase genes in E. coli isolated from hospitals in Malaysia19,36 and the Southeast Asian region. Any association between ESBL genes and integrons has not been investigated among Malaysian E. coli strains. Thus, this study was undertaken to (1) determine the prevalence of different types of beta-lactamase genes and their respective variants, (2) determine the level of resistance to ESCs conferred by different variants of ESBL and AmpC genes, (3) investigate the association between integrons and ESBL genes, and (4) investigate the clonality of ESBL- and AmpC-producing E. coli strains isolated from children in a Malaysian tertiary hospital.
Materials and Methods
Bacterial strains
One-hundred and ten nonrepeat E. coli strains from stool samples of children under the age of 5 years were obtained in 2009–2010, from a tertiary hospital in Johor Bahru, a city at the southern tip of Peninsular Malaysia. These strains were identified by the clinical microbiology laboratory using standard cultural methods and biochemical tests. The purity of the strains was confirmed using eosin methylene blue selective medium and the strains were stored in cryovials of veal infusion broth with 50% glycerol (Invitrogen) at −20°C and −85°C.
Antimicrobial susceptibility testing
Antimicrobial susceptibility of E. coli was determined using the disk diffusion method according to Clinical and Laboratory Standards Institute (CLSI) guidelines 8 on Mueller-Hinton agar (BD) with commercial disks (Oxoid Ltd.). The antimicrobial agents used were: ampicillin (AMP, 10 μg), tetracycline (TET, 30 μg), trimethoprim/sulfamethoxazole (SXT, 30 μg), nalidixic acid (NAL, 30 μg), streptomycin (STR, 10 μg), cefoperazone (CFP, 30 μg), kanamycin (KAN, 30 μg), chloramphenicol (CHL, 30 μg), ceftriaxone (CRO, 30 μg), amoxicillin/clavulanic acid (AMC, 20/10 μg), cefotaxime (CTX, 30 μg), ciprofloxacin (CIP, 5 μg), aztreonam (ATM, 30 μg), cefepime (FEP, 30 μg), gentamicin (GEN, 10 μg), ceftazidime (CAZ, 30 μg), amikacin (AMK, 30 μg), meropenem (MEM, 10 μg), and imipenem (IPM, 10 μg). The minimum inhibitory concentration (MICs) for CAZ, CTX, CRO, and ATM were determined using E-test strips (AB Biodisk). E. coli ATCC 25922 and ATCC 35218 were used as control strains.
Phenotypic detection of ESBL
E. coli strains were screened for presumptive ESBL production using four antimicrobial disks containing CAZ (30 μg), CRO (30 μg), CTX (30 μg), and ATM (30 μg). Presumptive-positive ESBL-producing E. coli strains were further tested for the ESBL production using the modified double-disk synergy test (DDST) 17 and E-test ESBL strips (CTX/CTX + clavulanic acid [CT/CTL] and CAZ/CAZ + clavulanic acid [TZ/TZL]) (AB Biodisk). These two methods were used to increase the efficiency of ESBL detection. Strains with positive results for DDST or ESBL E-test were termed ESBL producers. E. coli ATCC 25922 and ATCC 35218 were used as control strains. 8
Detection of genes encoding for beta-lactamase
Polymerase chain reaction (PCR) was used to detect the presence of seven types of beta-lactamase genes with primers previously described for the amplification of blaCTX-M 27 ; blaTEM, blaOXA-1, and blaSHV 26 ; blaVEB 19 ; blaDHA 28 ; and blaCMY-2. 1 Boiled suspensions of bacterial cells for the 110 strains were used as DNA templates. For ESC nonsusceptible strains that did not yield any amplicon for ESBL or AmpC genes, PCR detection of these beta-lactamase genes was repeated using genomic DNA prepared using the Wizard Genomic DNA Purification Kit (Promega). Strains that yielded positive amplification for blaCTX-M were further subgrouped into CTX-M groups 1, 2, 8/25, and 9 as described by Ensor et al. 11 PCR detection for the CTX-M-15 gene was carried out using primers described by Conceicao et al. 9 Representative amplified products were purified and submitted to a commercial facility (First BASE Laboratories) for sequencing to validate their identities. Strains with the expected amplicon size for blaCTX-M, blaTEM-1, blaOXA-1, and blaSHV genes that had been sequenced and validated by sequence alignment were used as positive controls.
Detection of O25-ST131 clone
Rapid detection of the O25-ST131 E. coli clone was carried out by PCR as previously described 7 for CTX-M-15-producing strains with inclusion of a positive control validated by DNA sequencing and alignment analysis.
Detection of class 1, 2, and 3 integrons
All 110 strains were tested for the presence of class 1, class 2, and class 3 integron-encoded integrases using PCR as described previously. 20 Amplification of the gene cassette regions was carried out for integrase-positive strains (using the 5′_CS/3′_CS primers for gene cassette-bearing class 1 integrons and the attI2-F/orfX-R primers for gene cassettes in class 2 integrons). 20 All PCR amplicons were purified and submitted to a commercial company for sequencing (First BASE Laboratories). For strains that shared similar pulsed-field profiles (PFPs; ≤1 band difference) but with different integron profiles, PCR detection of integrases and integrons was repeated using genomic DNA prepared using the Wizard Genomic DNA Purification Kit (Promega).
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) for the 110 strains was performed according to Thong et al. 38 with minor modifications. DNA plugs were digested with 12 U of XbaI (Promega) and DNA fragments were separated by electrophoresis for 25 hr in 0.5×TBE buffer at 14°C in a CHEF Mapper system (Bio-Rad) using pulse times of 2.16 to 54.17 sec with XbaI-digested Salmonella enterica serovar Braenderup H9812 as DNA size marker. 38 BioNumerics 6.0 (Applied Maths) was used for data analysis and a dendrogram was generated using the Dice coefficient and unweighted pair group method using arithmetic averages (UPGMA) at 1.5% tolerance level. 38
Statistical analysis
Statistical analysis was performed with R, version 2.12.2, 33 and Fisher's exact test was used to determine the association of integrons and ESBL-producing E. coli strains. A significance level of p<0.05 was considered as statistically significant.
Results
Antimicrobial resistance rates among the E. coli strains
Eighteen of the 110 (16.4%) E. coli strains were sensitive to all the 19 antimicrobials tested. The antimicrobial resistance rates were: AMP (60.0%), TET (42.7%), SXT (34.6%), NAL (30%), STR (25.5%), CFP (19.1%), KAN (18.2%), CHL (16.4%), CRO (13.6%), AMC (11.8%), CTX (10.9%), CIP (10.0%), ATM (6.4%), FEP (4.6%), GEN (2.7%), CAZ (2.7%), and AMK (0.9%). All the strains were sensitive to carbapenems. About 46% (51/110) of the strains were multidrug resistant (MDR). Table 1 summarizes the resistance profiles of the 51 MDR. Among the MDR strains, high resistance rates were observed for AMP (98%, n=50/51), TET (75%, n=38/51), and SXT (73%, n=37/51) (Table 1). Three MDR strains had the highest number of antimicrobial resistance (n=14) and were resistant to all broad-spectrum cephalopsorins and monobactam tested. Fourteen ESC-resistant and nine ESC-intermediate-resistant E. coli strains were also MDR.
AMP, ampicillin; TET, tetracycline; SXT, trimethoprim/sulfamethoxazole; NAL, nalidixic acid; STR, streptomycin; CFP, cefoperazone; KAN, kanamycin; CHL, chloramphenicol; CRO, ceftriaxone; CTX, cefotaxime; CIP, ciprofloxacin; ATM, aztreonam; FEP, cefepime; CAZ, ceftazidime; AMC, amoxicillin/clavulanic acid; AMK, amikacin; GEN, gentamicin.
Thirty percent (33/110) of the E. coli strains were presumptive ESBL producers based on initial four-antimicrobial disk-diffusion screening. Eighteen were ESBL producers based on DDST. Based on the phenotypic confirmatory test (ESBL E-test), 15 strains were ESBL producers while 2 were nondeterminate (Nd) as their MIC values for CTX/CTX + clavulanic acid and CAZ/CAZ + clavulanic acid were above the test range. According to the manufacturer's interpretation, Nd results might suggest the presence of an inhibitor-resistant enzyme (such as AmpC enzyme). Thirteen strains were confirmed as ESBL producers by both phenotypic detection tests (DDST and ESBL E-test methods).
Characterization of beta-lactamase genes
Out of the 7 beta-lactamase genes tested (blaTEM, blaCTX-M, blaOXA-1, blaSHV, blaVEB, blaDHA, and blaCMY-2) using PCR, only 3 were detected among the 110 E. coli strains: blaTEM (49.1%, n=54), blaCTX-M (11.8%, n=13), and blaCMY-2 (6.4%, n=7). Analysis of the DNA sequence of the blaTEM amplicons revealed that they belonged to subgroup blaTEM-1 and this was the most prevalent beta-lactamase gene detected.
All blaCTX-M-positive strains were ESBL producers with positive results for both DDST and ESBL E-test methods and the 13 blaCTX-M genes were further subgrouped into CTX-M-15 (n=10) and CTX-M-9 (n=3). Further DNA sequence analyses of the CTX-M-9 genes revealed two subtypes, CTX-M-14 (n=2) and CTX-M-27 (n=1). No CTX-M subgroup 2 or 8/25 was detected. All strains with blaCMY-2 or blaCTX-M genes were nonsusceptible to third-generation cephalosporins.
CTX-M enzymes had higher hydrolytic activity against CTX (MIC, 24–256 μg/ml) than CAZ (MIC, 1–16 μg/ml). Majority of the CTX-M-positive E. coli strains (11/13) showed high resistance levels to CTX with MIC values of ≥32 μg/ml. The MIC values for CTX-M-15- and CTX-M-14-producing E. coli for the four antimicrobial agents were as follows: CTX (16–256 vs. 24–48 μg/ml), CAZ (2–16 vs. 1–2 μg/ml), CRO (16–256 vs. 48–64 μg/ml), and ATM (3–96 vs. 3–8 μg/ml).
MIC values of CTX, CAZ, CRO, and ATM for CMY-2-producing strains were 1.5–3, 3–12, 2–8, and 3–12 μg/ml, respectively.
Prevalence of O25-ST131 clone
No O25-ST131 clone was present among the 10 CTX-M-15-positive E. coli strains based on primers described by Clermont et al. 7 although the positive control strain gave the expected band size.
Prevalence of class 1, 2, and 3 integrons
All 110 E. coli strains were screened for class 1, 2, and 3 integron-encoded integrase genes. No class 3 integron-encoded integrase gene was detected. Among the 110 strains, 48 (43.6%) and 4 strains (3.6%) harbored the intI1 and intI2 integrase genes, respectively. Of the 48 intI1-positive strains, only 21 yielded amplified products with the 5′_CS/3′_CS primers, implying the presence of gene cassettes, while all 4 intI2 integrase-positive strains harbored gene cassettes as part of class 2 integrons. Six types of gene cassettes were identified (Table 2). Three types of aadA genes (aadA1, aadA2, and aadA5) were identified, which confer resistance to STR and spectinomycin. The aadB gene (n=2), which confers resistance to a range of aminoglycosides, namely, GEN, KAN, and tobramycin, was found to be associated with aadA2 in the class 1 integrons. Four types of dfr genes (dfrA12, dfrA17, dfrA5, and dfrA7) were detected in class 1 integrons and one type (dfrA1) in a class 2 integron. The dfr genes encode for the enzyme dihydrofolate reductase that mediates resistance to trimethoprim. The dfrA and aadA genes can exist alone or in combination with other resistance genes. All E. coli strains that harbored dfr and aad genes were nonsusceptible to SXT and aminoglycosides, respectively. The genotypes of the gene cassettes found in the integrons corresponded to the resistotypes. Only sat1 gene was identified in class 2 integron strains, which confers resistance to streptothricin. No beta-lactamase genes were found within the gene cassettes that were amplified from the integrase-positive E. coli strains.
Genetic relatedness of the E. coli strains based on PFGE
PFGE subtyped XbaI-digested genomic DNA of 107 strains into 72 PFPs with a Simpson's index diversity value of 0.98, and F-value (coefficient of similarity) ranging from 0.45 to 1.0 (Fig. 1). XbaI digestion of the genomic DNA resulted in a total of 14–26 DNA fragments with sizes ranging from 20 to 800 kb. Three strains (EC062/10, EC063/10, and EC075/10) were untypeable although their DNA preparation was repeated three times. At 85% similarity, a total of 78 strains were clustered into 29 clonal groups, each comprising of 2–4 strains. Most of the strains within the same clonal group shared high degree of similarities with closely related PFPs and similar genotypes. Another 29 strains were less defined and were not clustered into any clonal group.

Dendrogram generated from PFGE profiles of the Escherichia coli strains using UPGMA based on Dice coefficients of similarity. aStrain code; bbeta-lactamase genes determined by polymerase chain reaction and validated by DNA sequencing; cintegron-type gene cassettes found in class 1 integrons: (1A) dfrA12-orfF-aadA2; (1B) aadB-aadA2; (1C) dfrA5; (1D) aadA1; (1E) dfrA7; (1F) dfrA17-aadA5; gene cassette found in class 2 integron: (2A) dfrA1-sat-aadA1; dpulsed-field profiles. The scale on top of the dendrogram indicates the percentage of similarity. ND, not detected; PFGE, pulsed-field gel electrophoresis; UPGMA, unweighted pair group method using arithmetic averages.
The 13 CTX-M-positive strains were subtyped into 11 PFPs. Three out of four strains (EC009/09, EC013/09, and EC019/09) that had the highest detectable MICs for CTX and CRO (256 μg/ml) were grouped together into a clonal group. These three strains also conferred relatively high resistance to ATM (MIC, 32–96 μg/ml) and CAZ (8–16 μg/ml). All three strains were highly related with one band difference and carried both blaCTX-M-15 and class 1 integron (dfrA12-orfF-aadA2). Seven blaCMY-2-positive strains from different pediatric wards had no apparent linkage as they were found scattered in the dendrogram.
The PFGE profiles of E. coli in the present study (2009/2010) were also compared with profiles of previously isolated E. coli (2004) in the same hospital. 19 The genetic relatedness of the E. coli strains obtained from the same hospital over a gap of more than 5 years was established from a dendrogram based on the PFGE profiles (Fig. 2). Overall, E. coli strains were genetically diverse except for two strains, EC096/10 (stool sample) and EC33/04 from our earlier study 19 (isolated from tracheal aspirate) that differed by only a single band (PFGE analysis for these two strains was repeated three times).
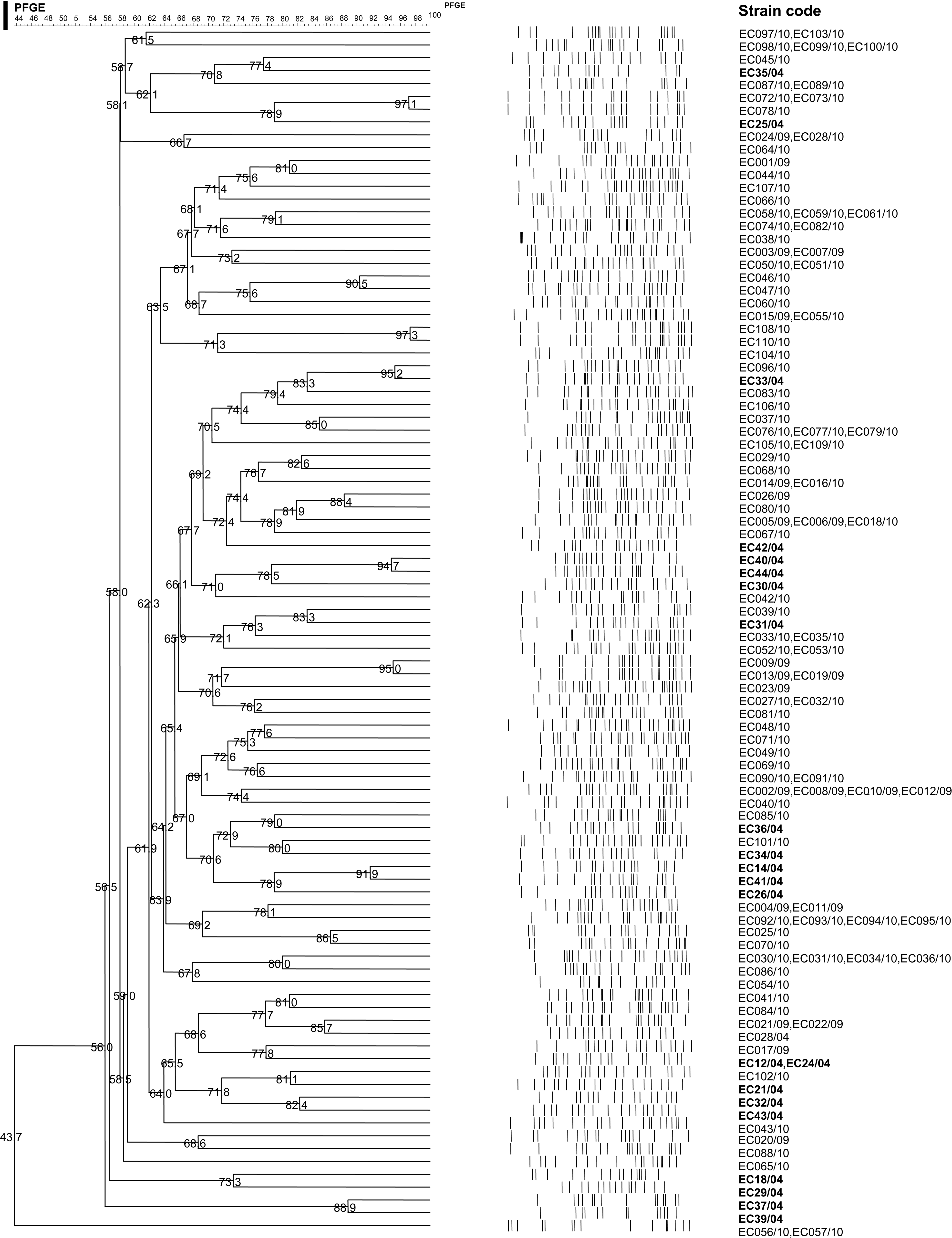
Dendrogram generated from PFGE profiles of the E. coli strains isolated in years 2004 and 2009/2010 obtained from the same hospital. Bold letters of strain codes (/04) indicate that the strains were obtained from the previous study (year 2004). 19 Strains EC33/04 and EC096/10 isolated 5 years apart showed one band difference.
Prevalence of integrons in ESBL- and non-ESBL-producing strains
A total of 15 out of the 24 integrons (class 1 + class 2) detected were found in non-ESBL-producing E. coli and no significant correlation was found between the ESBL phenotype and the carriage of integrons (p=0.23).
Discussion
Antimicrobial resistance in E. coli from children's stool samples in this study was compared with previous reports from Malaysia15,19 to observe the trend in antimicrobial resistance keeping in mind that the respective study populations were different. All three reports shared similar findings whereby E. coli strains had low resistance rates to carbapenems (0%–0.5%) and high resistance rates to AMP (60%–77%). The resistance rates for ESCs in this study were generally similar (∼±3%) to those reported in the earlier-mentioned studies with the exception of FEP and CAZ where resistance rates were approximately fourfold lower in our study. The MDR rate was also lower (46%) than those reported by Lim et al. 19 (64%) and it should be noted that 23 strains from our previous study 19 were from the same hospital, although from an earlier period (i.e., 2004).
Approximately 54% (n=13/24) of the nonsusceptible ESC (intermediate and resistant) strains were CTX-M positive. CTX-M was the most common type of ESBL observed in both studies (this study: 11.8%; Lim et al. 19 : 17.0%) with CTX-M-15 as the most common subtype in Malaysia19,36 and India. 35 This is in contrast to the report by Hawkey 13 that prevalence of CTX-M-15 is low in Asia. Although CTX-M-14 is the most prevalent variant of CTX-M in other Asian countries, such as Korea, China, and Taiwan,37,43,44 it is relatively low in Malaysia with rates of 15.4% (n=2/13) in this study and 31% (n=4/13) in a previous study. 36 Perhaps, the prevalence of a particular genotype is associated with different geographical regions. To the best of our knowledge, this is the first report of the presence of a blaCTX-M-27-producing E. coli strain in Malaysia. The low prevalence of blaCTX-M-27 concurred with the reports from Korea 37 and Canada. 32
E. coli strains that carried blaCTX-M genes exhibited higher resistance to CTX than CAZ (MICs of 8–32-fold dilutions), consistent with the classical phenotype of CTX-M enzymes. Nevertheless, a broad range of resistance levels to CTX (MICs of 24–256 μg/ml) for the E. coli strains that carried the CTX-M-encoding genes was observed, similar to a previous report by Edelstein et al. 10 This study shows that CTX-M-15 (MIC 2–16 μg/ml) has stronger hydrolytic activities against CAZ compared with its parental enzyme, CTX-M-14 (MIC 1–2 μg/ml). E. coli strains containing CTX-M-27 also exhibited higher resistance levels to CAZ compared with CTX-M-14 (8 vs. 1–2 μg/ml), which is in agreement with report from Bonnet et al. 3 Although the level of resistance (MIC value) appeared to be associated with the CTX-M subtype, however, it should be noted that the MIC value only provides an estimation of the antimicrobial resistance level.
Besides CTX-M-encoding genes, AmpC genes also play an essential role in conferring nonsusceptibility to broad-spectrum cephalosporins. In this study, blaCMY-2 was the only AmpC gene detected in 7 of the 110 E. coli strains. Six out of seven CMY-2-positive strains were nonsusceptible to ESCs while another CMY-2-positive strain had reduced susceptibility. All CMY-2-positive strains were resistant to AMC, a known trait of AmpC-type beta-lactamase (weakly inhibited by beta-lactamase inhibitors), 2 and consistent with an AmpC-type beta-lactamase phenotype. The CMY-2-producing strains also had a higher resistance level (MICs) to CAZ than for CTX, which is in agreement with the findings of Philippon et al. 31 Although the MIC values for CRO were elevated (3–12 μg/ml), it was interpreted as reduced susceptibility rather than resistance according to CLSI guidelines. 42 Plasmid-encoded AmpC-type beta-lactamase-producing strains are often not resistant to third-generation cephalosporins because they are found in low copy numbers. 12
The remaining five strains that were nonsusceptible to ESCs did not contain any ESBL or AmpC genes, indicating that resistance to cephalosporins may be conferred by other resistance mechanisms. 30
The association between ESBL genes and integrons has been reported worldwide,20,21,40 but no studies have yet been reported for Malaysian E. coli strains. In this study, class 1 integrons were the most common integrons in the Malaysian E. coli strains, but no gene cassette encoding ESBL genes was detected. Sequence analyses of the amplicons revealed a predominance of gene cassettes that conferred resistance to STR and spectinomycin (aadA genes) and trimethoprim (dfr genes), similar to the report of White et al. 41 A total of 15 out of 25 integrons (class 1 + class 2) were found in non-ESBL-producing E. coli. Integrons were also not significantly found in either ESBL or non-ESBL producers (p>0.05). These findings infer that integrons are unlikely to have contributed to the dissemination of the ESBL genes in the Malaysian E. coli strains. Previous studies by Machado et al.20,21 and Vinué et al. 40 also indicated that integrons do not contribute much to the dissemination of ESBL genes unless these genes are located within integrons (e.g., CTX-M-9 that linked to In60 integron). The transmissibility of ESBL genes could be linked to other mechanisms such as insertion elements and plasmids. 34
E. coli strains that contain blaCTX-M genes were genetically diverse. This observation concurred with other studies25,40 that reported a low clonal relationship among the strains containing blaCTX-M genes. This indicates that the dispersion of the E. coli strains with blaCTX-M genes was not likely to be due to the dissemination of a particular resistant clone harboring the gene. However, the three strains that contained blaCTX-M-15 (EC013/09, EC019/09, and EC009/09) were highly related. These strains shared similar PFPs and resistotypes and might have been derived from the same clone. It was also found that EC096/10 from the present study and EC33/04 from the previous study 19 were closely related (one band difference) (Fig. 2). Similar observations where epidemiological unrelated E. coli strains had closely related PFGE patterns have been reported by Mamlouk et al. 22 Further analysis of these strains showed that they were of different resistotypes. No integron was detected in both strains while blaTEM-1 was detected only in EC33/04. According to van Belkum et al., 39 strains with indistinguishable typing patterns do not necessarily belong to the same clone because different strains may yield indistinguishable patterns using the same typing method. Nevertheless, the high discriminatory power of PFGE strongly implies that EC096/10 and EC33/04 are derived from the same clone. The clonality of these two strains was further confirmed by rep-PCR 24 and plasmid profiling. Both strains yielded the same rep-PCR profile and both did not harbor any plasmids (unpublished data). These results strongly imply that EC096/10 and EC33/04 were likely from the same clone that had persisted in the hospital environment with gain or loss of resistance due to the acquisition or deletion of respective genes that are likely to be located on MGEs.
In summary, the 110 E. coli strains isolated from pediatric wards of a tertiary hospital in Malaysia showed high resistance rates to AMP and total sensitivity to carbapenems. The MDR rates were lower compared with other previously reported Malaysian strains. Class 1 integrons remained the major class of integrons. No gene cassette encoding ESBL genes was found and no significant association could be made between ESBL production and the carriage of integrons. CTX-M-15 was the most common ESBL found in this study although low prevalence was reported in other parts of Asia, supporting that the prevalence of genotypes is associated with different geographical regions. Possible clonal expansion was observed for a few CTX-M-15-positive strains but no O25-ST131 clone was detected. Further studies to determine the transferability of MGEs in clinical strains of E. coli are necessary to further understand the dissemination of resistance determinants.
Footnotes
Acknowledgments
This work was supported by Postgraduate Research Fund (PS318/2010B) from University of Malaya and National Japanese Institute of Infectious Diseases (57-02-03-1015). Wing Sze Ho is supported by a fellowship from University of Malaya.
Disclosure Statement
No competing financial interests exist.