
Abstract
Ampicillin and vancomycin are important antibiotics for the therapy of Enterococcus faecalis infections. The ampicillin resistance gene pbp5 is intrinsic in Enterococcus faecium. The vanC1 gene confers resistance to vancomycin and serves as a species marker for Enterococcus gallinarum. Both genes are chromosomally located. Resistance to ampicillin and vancomycin was determined in 484 E. faecalis of human and porcine origin by microdilution. Since E. faecalis are highly skilled to acquire resistance genes, all strains were investigated for the presence of pbp5 (and, in positive strains, for the penicillin-binding protein synthesis repressor gene psr) and vanC1 (and, in positive strains, for vanXYc and vanT) by using polymerase chain reaction (PCR). One porcine and one human isolate were phenotypically resistant to ampicillin; no strain was vancomycin resistant. Four E. faecalis (3/1 of porcine/human origin) carried pbp5 (MIC=1 mg/L), and four porcine strains were vanC1 positive (minimum inhibitory concentration [MIC]=1 mg/L). Real-time reverse transcriptase (RT)-PCR revealed that the genes were not expressed. The psr gene was absent in the four pbp5-positive strains; the vanXYc gene was absent in the four vanC1-positive strains. However, vanT of the vanC gene cluster was detected in two vanC1-positive strains. To our knowledge, this is the first report on the presence of pbp5, identical with the “E. faecium pbp5 gene,” and of vanC1/vanT in E. faecalis. Even if resistance is not expressed in these strains, this study shows that E. faecalis have a strong ability to acquire resistance genes—and potentially to spread them to other bacteria. Therefore, close monitoring of this species should be continued.
Introduction
Materials and Methods
Bacterial strains
In the period 2002–2009, a total of 484 E. faecalis strains from Bavaria were isolated on Citrate Azide Tween Carbonate (CATC) Agar (VWR) and biochemically identified as previously described. 44 Of these, 238 were of porcine and 246 of human origin (Table 1). Species verification of the vanC1- and pbp5-positive isolates was performed by multiplex polymerase chain reaction (PCR) as described by Jackson et al. 17 For the characterization of the genetic variability of these strains, the randomly amplified polymorphic DNA (RAPD)-PCR technique was applied by using the M13R2 primer as described by Martín et al. 29 Cluster analysis was performed by using the GelCompar Software V 5.10 (Applied Maths).
Breakpoints according to DIN. Breakpoints according to EUCAST (available at: www.eucast.org/eucast_disk_diffusion_test/breakpoints) deviate slightly (S≤4; R>8).
Three isolates were pbp5 positive.
One isolate was pbp5 positive.
According to DIN (S≤4; R>8 mg/L) and to EUCAST (S≤4; R>4 mg/L), all isolates were susceptible to vancomycin.
Four isolates were vanC1 positive.
MIC, minimum inhibitory concentration; DIN, Deutsches Institut für Normung (German Institute for Standardization).
Antimicrobial susceptibility testing
Phenotypic resistance characteristics were tested by the broth microdilution method according to DIN 58940-81 (2002) as previously described.9,44 Resistance testing was repeated for the pbp5- and vanC1-positive isolates. Incubation time for the vanC1-positive isolates was additionally prolonged to 48 hr, since Hirakata et al. 15 recognized a shift to resistance dependent on inoculation time.
Detection of pbp5 and vanC1 by PCR
DNA was extracted from three colonies of pure cultures growing on Standard I Nutrient Agar (Merck), containing 7% defibrinated sheep blood (Fiebig) with the DNeasy Blood & Tissue Kit (Qiagen) following the manufacturer's instructions. Primers were synthesized by TIB MOLBIOL Syntheselabor GmbH. PCR protocols for the detection of pbp5 and vanC1 are given in Table 2. Each PCR cycle contained one positive control (pbp5: E. faecium No. 51 21 ; vanC1: Enterococcus gallinarum BM 4174; Institut Pasteur) and one negative control (master mix without template DNA). Reaction products were visualized under UV light after electrophoresis through a 1.5% agarose gel (BioRad) containing 0.285 μg/ml ethidium bromide (Sigma-Aldrich). Extraction and PCR runs were repeated twice for each pbp5- or vanC1-positive result in order to verify the results and to exclude contamination.
Primers were designed with the help of Primer3 (v. 0.4.0), available at http://frodo.wi.mit.edu/; vanC1 primers were designed as described elsewhere. 11
Each mixture for endpoint PCR (total volume=25 μl) consisted of 3.5 μl of 10× PCR buffer, 1.5 μl MgCl2, 1.0 μl dNTPs, 0.25 μl Taq-Polymerase (Qiagen), forward and reverse primer (variable), and 1.0 μl template DNA.
Thermocycler settings: initial denaturation at 94°C for 5 min; 35 cycles of denaturation at 94°C for 1 min, annealing (variable), elongation at 72°C for 1:30 min; final extension at 72°C for 10:00 min.
Each mixture for real-time PCR (total volume=20 μl) consisted of 1.6 μl MgCl2, 2 μl SYBR Mix (Roche), and 2 μl template DNA.
LightCycler® settings: initial denaturation at 95°C for 10 min; 35 cycles of denaturation at 95°C for 10 sec, annealing (variable), elongation at 72°C for 8/10/8/10/33 sec (psrfm/vanXYc/vanT/pbp5rt/vanC1rt); cooling at 40°C for 30 sec.
PCR, polymerase chain reaction.
Detection of psr, vanXYc, and vanT by real-time PCR
The pbp5-positive strains were tested for the presence of the penicillin-binding protein synthesis repressor gene psr 56 (gi|790434). The vanC1-positive strains were tested for the presence of the D,D-dipeptidase-carboxypeptidase encoding vanXYc gene 1 (gi|9294728). The vanC1-positive strains were also tested for vanT, which encodes a membrane-bound serine racemase 1 (gi|9294728). For primer sequences and PCR conditions, see Table 2. For positive and negative controls, see “Detection of pbp5 and vanC1 by PCR”.
Analysis of gene expression by real-time reverse transcriptase-PCR
For quantitative reverse transcriptase (RT)-PCR, bacterial suspensions of the pbp5- and vanC1-positive strains were adjusted to optical density (OD)600=0.8. Total RNA was extracted with the NucleoSpin® RNAII kit (Macherey-Nagel) using 5 mg/ml lysozyme (Sigma-Aldrich). For primer sequences (pbp5rt and vanC1rt) and PCR conditions, see Table 2. For positive and negative controls, see “Detection of pbp5 and vanC1 by PCR”. DNA contamination of the RNA extracts was excluded by performing a pooled “no-RT” control.
Sequence analysis
The bands of two pbp5-positive strains (one of porcine and one of human origin) were excised from the gel and sent for sequencing (Sequiserve). One vanC1 and one vanT bands were also sent for sequencing. Sequence similarity was compared by using the NCBI BLAST database (Basic Local Alignment Search Tool; www.ncbi.nlm.nih.gov).
Results
Table 1 shows that two of the 484 isolates were phenotypically resistant to ampicillin (minimum inhibitory concentration [MIC]>8 mg/L), whereas no vancomycin-resistant strain was observed. The ampicillin-resistant strains originated from a stool sample of a diseased pig and from a stool sample of a person without clinical disease; none of them contained pbp5. All isolates that were intermediately resistant to ampicillin, as well as all isolates with MIC=4 mg/L for vancomycin originated from pig manure samples.
In total, four E. faecalis with an MIC of 1 mg/L for ampicillin were positive for the presence of the pbp5 gene. The strains of porcine origin (n=3) were isolated from stool samples of diseased pigs from different farms. The human strain originated from a stool sample of a healthy person. The sequence analysis confirmed the PCR results. According to the NCBI BLAST database, the nucleotide sequences of the two strains had 99% similarity to a section of the “E. faecium pbp5 gene (D63r)” (GenBank: X84860.1). In the porcine strain, one point mutation was present on position 882 of the PCR product. The A-to-C polymorphism is a transversion that leads to a neutral amino acid substitution at position 1,482 in the PBP5 polypeptide. This change is consistent with the substitution of an isoleucine residue with leucine. In the human strain, two point mutations were present: the T-to-C polymorphism at position 301 leads to a substitution of tyrosine with histidine; the T-to-C polymorphism at position 481 leads to a substitution of cysteine with arginine. The psr gene was not detectable in the pbp5-positive strains.
Four vancomycin-susceptible isolates (MIC=1 mg/L), all originating from stool samples of diseased pigs from two different farms, were vanC1 positive. The sequence analysis confirmed the PCR results. In detail, the nucleotide sequence of the vanC1 PCR amplicon exactly matched a section of the “E. gallinarum strain eS464 VanC1 (vanC1) gene” (GenBank: EU151772.1; 100% identities; 0 gaps). The vanXYc gene was not detectable, but the vanT gene was present in two strains. The PCR result was also confirmed by sequence analysis. In detail, the respective nucleotide sequence exactly matched a section of the “E. gallinarum vanC vancomycin-resistance gene cluster,” which harbors the vanT gene 1 (GenBank: AF162694.1; 100% identities; 0 gaps).
The species-specific multiplex PCR confirmed that the positive samples were really E. faecalis. In detail, all investigated isolates were positive for E. faecalis, but negative for E. faecium and for E. gallinarum, respectively. The Pearson correlation coefficients obtained by RAPD-PCR were between 86% and 71% in pbp5-positive strains, with the strain of human origin being the most dissimilar. Pearson correlation coefficients of vanC1-positive strains were between 93% and 73% (Fig. 1). No corresponding transcripts of the pbp5 and vanC1 genes were detectable by real-time RT-PCR in the positive strains.
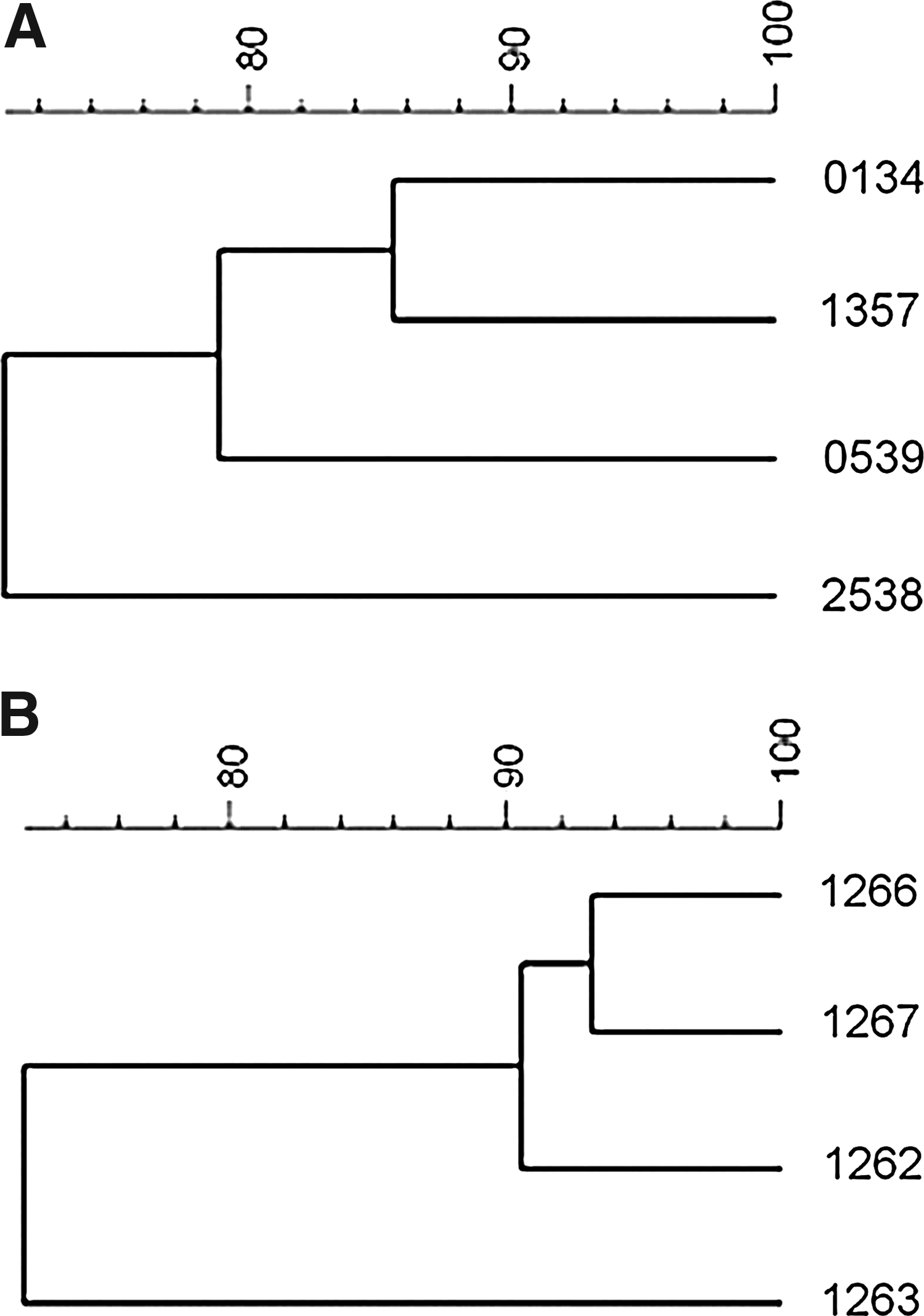
Cluster analysis of
Discussion
The low-affinity PBP5 in E. faecium, encoded by the pbp5 gene, confers low-level resistance to ampicillin. More than 90% of the strains isolated from German hospitals are classified as ampicillin resistant. 14 On the contrary, ampicillin is still a valuable therapeutic in E. faecalis infections. 20 This is in accordance with the MIC results of our study, where only two isolates were resistant to ampicillin.
There is one report on the occurrence of a gene designated “pbp5” in one E. faecalis termed “56 R” 47 (gi|467805), but the sequence homology with the amplified product of the present study is <70%. Actually, the 56 R pbp5 of Signoretto et al. 47 is almost identical (99%; 0 gaps) to the pbp4 gene of E. faecalis JH2-2 as described by Duez et al. 10 Subsequent investigations, working with “pbp5” containing E. faecalis, always refer to the 56 R strain.10,25–27,43,48–50,56 The pbp5 gene of E. faecium is chromosomally localized, but its transferability to E. faecalis has been experimentally proven. 41 Nevertheless, apart from Poeta et al. 36 and Radimersky et al., 37 who tested 2 and 36 E. faecalis strains negative for the presence of pbp5, no further screenings have been reported. This means that the present study is the first report on the detection of pbp5 in E. faecalis apart from mating experiments.
Likewise, no scientist who searched for vanC1 in E. faecalis detected the gene in this species.3,4,7,15,16,19,23,28,32,35,38,46,51,55 Actually, it is claimed to be even superfluous to test E. faecalis or other Enterococcus species for the presence of vanC1, since this gene is supposed to be species specific for E. gallinarum. 22 There is one report on the detection of vanC1 in one E. faecalis and in one E. faecium strain, 34 but a subsequent analysis of the 16S rRNA sequence revealed that both vanC1-containing strains were in reality E. gallinarum. 33 In the present study, a species identification error is unlikely because the distinctly red well-developed colonies on CATC-agar are very characteristic for E. faecalis; biochemical reactions were typical, and what is more, the species-specific PCR 17 confirmed the results of the conventional bacteriological investigation. Noticeably, all vanC1-positive isolates were isolated from pigs notwithstanding that vancomycin is a reserve antibiotic in human medicine and not approved for livestock use in Germany. The glycopeptide growth promoter avoparcin was banned from livestock feed in Germany in 1996 in order to reduce the risk of cross-resistance to vancomycin. 5 This indicates that E. faecalis might have acquired the vanC1 gene from the natural vanC1 carrier E. gallinarum, which is a normal inhabitant of the porcine gut.8,42 Normally, the three genes vanC1, vanT, and vanXYc are clustered. 1 The finding that, in our study, only two of the four vanC1-positive strains were vanT positive and none contained vanXYc might be attributable to a high instability of these genes in E. faecalis after cell division, as it has been reported, for example, for unstable elements in Staphylococcus aureus. 52
All pbp5-positive strains were susceptible to ampicillin. The same is true for vanC1; the vancomycin MIC remained as low as 1 mg/L for the vanC1-positive isolates even after prolonged incubation (48 hr), as recommended by Hirakata et al. 15 The phenotypic susceptibility can be explained by the fact that the resistance genes pbp5 and vanC1 were not expressed in the investigated strains, as shown via real-time RT-PCR. The regulating mechanisms of expression of pbp5 are unknown, but it has been suggested that the upstream open reading frame (ORF) psr (penicillin-binding protein synthesis repressor) might have a negative impact on PBP5 production. 24 On the contrary, Rice et al. 40 suggest that PSR does not serve as a repressor of pbp5 transcription. Further investigations are necessary to clarify these questions.
Nevertheless, even if the strains of the present study were phenotypically susceptible to ampicillin and vancomycin, it is a cause for concern that E. faecalis are able to naturally acquire pbp5 and vanC1 from the bacterial community, since the possibility that complete gene clusters and functional genes will be transferred and expressed cannot be ruled out. This study underlines the need to continue being aware that E. faecalis are emerging nosocomial pathogens on the one hand 31 and very potent resistance gene collectors—and possibly donors—on the other hand.13,44,45 These risk factors affirm that enterococci should always be closely monitored, and this applies not only to the human clinical isolates, but also to the commensals from their diverse habitats.
Footnotes
Acknowledgments
This work was supported by the Bavarian State Ministry of the Environment, Public Health and Consumer Protection and by the Bavarian State Ministry of Agriculture and Forestry. The authors thank Heike Kliem and technicians for providing the laboratory and equipment for real-time RT-PCR. We also thank Dr. Ingo Klare (Robert Koch Insititut, Wernigerode, Germany) for kindly providing the reference strains.
Disclosure Statement
No competing financial interests exist.