
Abstract
Klebsiella pneumoniae carbapenemase (KPC)-producing Klebsiella pneumoniae infections are associated with high mortality; however, little is known about the virulence determinants of KPC-producing K. pneumoniae. At the Veterans Affairs New Jersey Healthcare System (VA NJHCS), we investigated the virulence and plasmid transferability of 60 clinically unique KPC-containing K. pneumoniae isolates. All 60 isolates were negative for known virulence factors K1, K2, and K5 capsular antigens; rmpA; and the aerobactin gene by polymerase chain reaction. Isolates varied in their susceptibility to neutrophil phagocytosis, but were less resistant than the virulent serotype K1 isolate. Additionally, no deaths were seen on murine lethality studies. Conjugation results of this study showed that the blaKPC gene can be transferred into an Escherichia coli J-53 strain but not to E. coli JP-995. However, the stability is very limited as E. coli J-53 does not retain blaKPC-containing plasmids for any period of time. The lack of virulence factors in the set of KPC-producing K. pneumoniae studied suggests that morbidity and mortality may be due to detection issues or lack of effective antibiotics.
Introduction
Materials and Methods
From January 2007 to June 2010, 60 clinically unique, ertapenem-resistant KP isolates were acquired from the clinical microbiology laboratory at the Veteran Affairs New Jersey Health Care System (VA NJHCS) for this Institutional Review Board (IRB)-approved study (MIRB 00937). Virulence and conjugation testing was performed at the National Health Research Institutes of Taiwan.
Susceptibility testing
Antimicrobial susceptibility was determined by the broth microdilution method, 1 according to the Clinical and Laboratory Standards Institute's (CLSI) guidelines published in 2011. The following antimicrobial agents were used: cefazolin, amoxicillin/clavulanic acid, cefoxitin, cefotaxime, ceftazidime, imipenem, amikacin, gentamicin, ciprofloxacin, and trimethoprim-sulfamethoxazole.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) was performed as described previously. 9 The resulting genomic DNA profiles, or fingerprints, were interpreted according to established guidelines.20,24
Confirmation of KPC production and KPC types by sequencing
All isolates selected were confirmed for KPC production by polymerase chain reaction (PCR). A gene-specific primer set, KPC-F (5′-ATG TCA CTG TAT CGC CGT CT-3′) and KPC-R (5′-TTT TCA GAG CCT TAC TGC CC-3′), was used to determine the presence of the KPC β-lactamase sequence. 2 The amplicons (893 bp) were sequenced using an automated sequencer by the Sanger method (ABI Prism 377 sequencer; Perkin-Elmer). 21 The generated sequences were compared to the National Center for Biotechnology Information (NCBI) database at www.ncbi.nlm.nih.gov/blast/
Conjugation experiments
In this part of the study, two consecutive conjugation experiments were performed. In the first set of experiments, the sodium azide-resistant strain Escherichia coli J-53 (kindly provided by Prof. Jacoby, Division of Infectious Diseases, Lahey Clinic, Burlington, MA) or a rifampin-resistant strain of E. coli JP-995 (kindly supplied by J. Pittard, Department of Microbiology, University of Melbourne, Melbourne, Australia) was used as the recipient.23,26 None of the donor strains or wild-type KP with KPC could grow on MacConkey agar with sodium azide or rifampin (100 μg/ml). The recipient and donor strains were inoculated separately into a brain–heart infusion broth (Oxoid) and were incubated at 37°C for 4 hours. They were then mixed together at a ratio of 1:10 (by volume) and were incubated overnight at 37°C. A 0.1-ml volume of the overnight broth mixture was then spread onto a MacConkey agar plate containing azide or rifampin (100 μg/ml) and imipenem (1 μg/ml). Transconjugants were then selected from the agar plate.
In the second phase of conjugation experiments, the transconjugant of E. coli J-53 containing the KPC plasmid acted as a donor. A serotype K2 KP with intrinsic resistance only to ampicillin was used as a recipient. Mating procedure was described as above except by using specific selection brilliant green agar-containing inositol-nitrate-deoxycholate (BIND) for KP. 30 Serotype K2 KP transconjugants were selected using the Klebsiella-selective medium BIND supplemented with 2 μg/ml of imipenem. None of the donors, including E. coli J-53 with KPC, recipient, and serotype K2 K. pneumoniae, could grow on the selective BIND medium with imipenem. The only colonies that could grow on BIND with imipenem were the serotype K2 KP strain harboring the imipenem-resistant KPC plasmid. Transconjugants from the first- and second-round conjugation experiments were subcultured onto MacConkey or Mueller Hinton agar (BBL) for 14 days to check for KPC plasmid stability in E. coli and KP after conjugation.
PCR detection of virulence-associated genes rmpA and aerobactin, and serotypes K1, K2, and K2
PCR experiments to determine the presence of the specific genes for serotype K1, K2, and K5; rmpA; and the aerobactin gene were performed as previously described. 25
Virulence studies
In vitro and in vivo assays to assess virulence were performed by neutrophil phagocytosis and mice lethality experiments. Neutrophil phagocytosis was performed as previously described. 18 A serotype K1 KP (STL-43) isolated from a liver abscess was used as a control strain. This strain is hypervirulent with a median lethal dose (LD50) <102 colony forming unit (CFU) and is highly phagocytic resistance (<10% of phagocytic rate 15 minutes). Two isogenic mutants from wild-type STL-43 with capsule deficiency and became highly susceptible to phagocytosis (∼90% of phagocytic rate at 15 minutes) were also used as controls. 18 For the determination of LD50 in mice, pathogen-free, adult, 6–8-week-old, male BALB/c mice weighing 20–25 g were obtained from the National Laboratory Animal Center. This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. All animal care procedures and protocols were approved by the Institutional Animal Care and Use Committee of the National Health Research Institute (NHRI-IACUC-096004-A). Tenfold serial dilutions of KP were made from a starting concentration of 106 CFU, and the adult BALB/c mice had intraperitoneal injections with 0.1 ml of each concentration. Six mice were used as a sample population for each bacterial concentration. 4 Symptoms and signs of infection were observed for 14 days. Survival of the inoculated mice was recorded, and the LD50 was calculated using SigmaPlot version 7.0 from SPSS Inc.
Results
Antimicrobial susceptibility and KPC types
Among 60 clinically unique KPC-positive KP isolates, 100% resistance was found to piperacillin–tazobactam, cefazolin, cefotaxime, ceftazidime, ceftriaxone, and ciprofloxacin followed by 92% resistance to cefoxitin and 33% to gentamicin. Reduced susceptibility to imipenem and/or meropenem was seen in 95% of the isolates. Sequencing results showed two types of KPC (17 KPC-2 and 43 KPC-3 isolates).
Conjugation and blaKPC plasmid stability test
On day 1 of conjugation experiments, none of the 13 rifampin-susceptible KP strains, including 7 KPC-2 and 6 KPC-3 isolates, could transfer the blaKPC plasmid into recipient E. coli JP-995. On the contrary, plasmids carrying blaKPC from a parental nonserotype K2 KP, in all 60 isolates, were successfully transferred into sodium azide-resistant E. coli J-53 on the day-1 conjugation experiment. E. coli J-53::blaKPC transconjugants were obtained from the MacConkey agar plate containing azide (100 μg/ml) and imipenem (2 μg/ml) (Fig. 1A). Subculture E. coli J-53::blaKPC into the same selective plates showed no growth of E. coli J-53::blaKPC transconjugants, suggesting the loss of blaKPC or instability of the blaKPC plasmid in E. coli (Fig. 1B). By using E. coli J-53
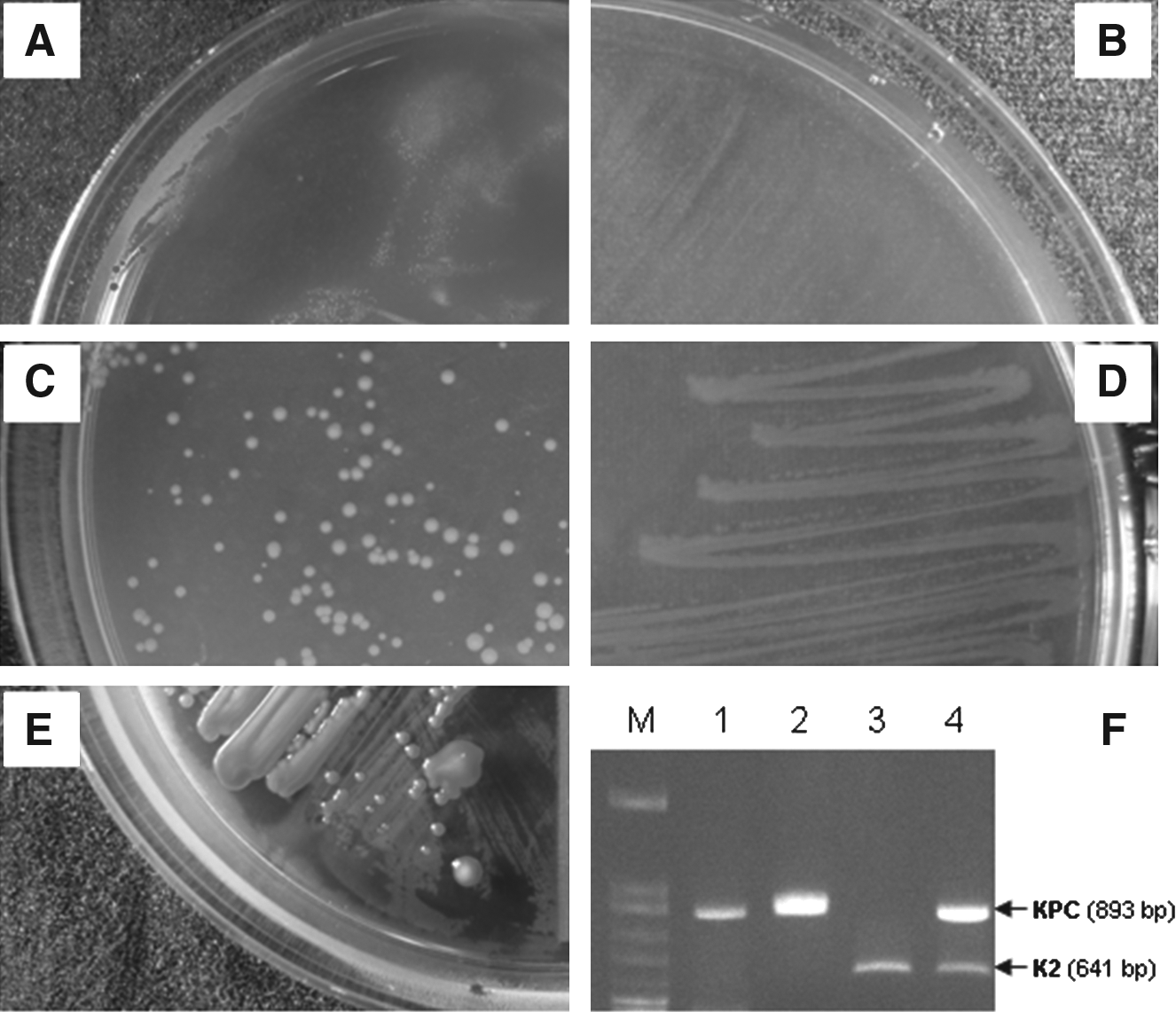
Conjugation experiments of plasmids carrying blaKPC.
Detection of virulence-associated determinants, neutrophil phagocytosis, and mice lethality experiments
Detection of K1, K2, and K5 virulence-associated serotypes, and rmpA and aerobactin genes, revealed that all 60 isolates were non-K1, -K2, or -K5 serotypes and did not carry rmpA and aerobactin genes. Twenty-five isolates were selected for neutrophil phagocytosis assays based on their diversity of PFGE profiles. These isolates varied in neutrophil phagocytic resistance (Fig. 2). Seven isolates were phagocytic resistant as determined by the phagocytosis rate calculated using the percentage of neutrophil-ingested fluorescein isothiocyanate-labeled bacteria over time. Phagocytic rates around 25%–35% at 15 minutes and the susceptible phagocytic rate >40% at 15 minutes, respectively, were selected for further mice lethality studies. Murine lethality studies by IP injection showed that LD50 were >106 CFU for all seven isolates, indicating low virulence. In contrast, LD50 of the known virulent strain STL-43 was <102 CFU.

Phagocytosis assay for randomly selected blaKPC isolates. The phagocytosis rate was calculated using the percentage of neutrophil-ingested fluorescein isothiocyanate-labeled bacteria over the time. The blaKPC isolates are labeled as filled diamonds. Serotype K1 isolates obtained from previous studies30 were used as a control for virulence assessment. STL-43 (open triangle) was obtained from patients with liver abscess. STL43ΔrfbP (open rectangle) and STL43ΔWzyKPK1 (open circle) were capsule-deficient nonpolar-rfbP and -WzyKPK1 mutant strains derived from STL-43, respectively. 4
Discussion
KPC KP has become an increasing prevalent pathogen and is now endemic in the Northeastern United States.3,5,29 Clinical mortality due to KPC KP has been well documented. 11 However, virulence and plasmid transmissibility of KPC KP have not been well investigated. The findings in this study hope to increase the understanding of virulence and plasmid transmissibility of KPC-producing KP. KPC-producing KP infections are usually associated with resistance to multiple classes of antibiotics, and treatment options are very limited. Furthermore, testing for KPC has been problematic. 14 Improper therapy may contribute to the increased morbidity and mortality seen in KPC KP infections.
After probing for known virulence factors, we found that these 60 KPC KP isolates did not contain genes for K1, K2, and K5 capsular antigens; rmpA; or the aerobactin gene. We selected these virulence markers as these have been found in hypervirulent strains of KP associated with liver abscess and metastatic pyogenic infections. 22 In addition, intraperitoneal injections of KPC KP in murine lethality models did not cause mortality even at a very high dose of >106 CFU of KP. The combination of the lack of KP virulence factors and no deaths seen in murine models suggests that mortality in KPC-producing KP infections may not be attributed to increased virulence.
Transmissibility of plasmids containing the blaKPC gene to other Enterobacteriaceae has also been of great concern. Although the conjugation results of this study showed that the blaKPC gene can be transferred into the E. coli J-53 strain, its stability is very limited as E. coli J-53 does not seem to be able to retain blaKPC for any period of time. In addition, we were not able to transfer the blaKPC gene into E. coli JP-995. The blaKPC-containing plasmid is conjugable, but was most stable on a species-specific level. These results seem to agree with what we see at the VA NJHCS. Since the first reporting of KPC KP at VA NJHCS in 2007, we have not seen carbapenem-resistant E. coli. However, although rare, E. coli containing blaKPC has been documented. 16 This may be due to the difference in the strains of E. coli. A previous study has shown that stability and the frequency of plasmid conjugal transfer are varied due to genetic elements such as replicons and transposons. 1 The KPC gene is located in a Tn3-based transposon (Tn4401), which explains the rapid spread among Enterobacteriaceae. 8 Cuzon et al. were able to demonstrate the great efficiency of integration and transpostion of Tn4401 among different plasmids without insertion-site specificity. 8 In this study, transmission of the KPC plasmid could be obtained in the first day of co-culturing the donor KP and recipient E. coli. However, losing carbapenem resistance by single colony subculture of the transconjugant into the same medium after 24 hours suggests the lack of genetic elements required for maintaining the stability of the KPC plasmid in recipient E. coli. Further investigation on the transmissibility of these KPC-containing E. coli needs to be done.
An important finding in this study is that the KPC-producing KP does not have known virulence factors found in KP strains causing liver abscesses in Asia; however, other virulence factors have been recently described. Adhesins, biofilm production, and heat resistance (the clpK gene) have been associated with increased pathogenicity in K. pneumoniae isolates. 13 Fuursted et al. evaluated the presence of virulence factors in a single strain of carbapenemase-producing KP carrying the New-Delhi metallo-beta-lactamase-1 (NDM-1) and compared it to noncarbapenemase-producing strains of KP. 13 The NDM-1-producing KP was found to be a K2 serotype and a biofilm producer. In addition, the isolate was more virulent than the comparative strains in the murine sepsis model. We did not investigate these other potential virulence factors; however, no murine deaths on lethality studies suggest that mortality due to KPC KP is not due to increased virulence. Other contributing factors may include advanced age, procedures, and line placement, among other reported risk factors. However, further dissemination of the KPC plasmid into virulent types of KP cannot be excluded in the future. Efforts to assess the factors that contribute to attributable mortality in KPC KP infections should be continued, including the study of other virulence determinants, antibiotic resistance, host responses, and their relationship with clinical characteristics and outcomes.
Footnotes
Acknowledgment
This work was supported by grants from the National Science Council (NSC-98-2314-B-016-024-MY3), Taiwan Center for Diseases Control (DOH-100-DC-1025), and from the National Health Research Institutes, Taiwan.
Disclosure Statement
No competing financial interests exist.