
Abstract
Abstract
The aim of this study was to develop and analyze in vitro azithromycin (AZM)-resistant mutants of Escherichia coli and Shigella boydii. Three clinical isolates of E. coli and one S. boydii isolated from feces samples collected from children under 5 years of age with diarrhea in Lima, Peru were inoculated onto Mueller-Hinton plates containing increasing serial dilutions of AZM ranging from their specific minimal inhibitory concentration (2 or 4 mg/l) to 64 mg/l. From these plates, 16 AZM-resistant mutants were selected to determine the stability of the resistance and the presence of cross resistance with other antibiotics. The role of Phe-Arg-β-Naphthylamide (PAβN)-inhibitible efflux pumps as well as the presence of mutations in the rplV, rplD, and rrlH (23S rRNA) genes and alterations in the outer membrane profiles were determined in these 16 mutants. The rate of mutation ranged from < 2.70×10−10 to 2.17×10−7 for E. coli and from < 9.58×10−10 to 1.05×10−8 for S. boydii. E. coli mutants showed an increase in the AZM-MIC up to sixfold with one strain achieving a MIC >256 mg/l. In contrast, S. boydii only presented increases of up to twofold in MIC levels. All the strains obtained, but one showed stable AZM resistance. In the presence of PAβN, the AZM MICs decreased to parental levels in Shigella mutants, while no MIC returned to parental levels among the E. coli mutants. No cross resistance to other classes of antibiotics was found. These results show the relevance of PAβN-inhibitible efflux pumps in the basal levels and development of AZM resistance. Further studies to characterize the remaining unidentified mechanisms of AZM resistance are needed.
Introduction
In Peru, pediatric diarrhea has special relevance in periurban and rural areas. 23 Previous studies have reported high levels of antibiotic resistance by diarrheogenic pathogens in this country,23,24 as well as their extension to commensal microorganisms.1,27
Azithromycin (AZM) is an antimicrobial agent belonging to the macrolide family that has mainly been used to treat infections caused by Gram-positive microorganisms. 6 However, this antimicrobial agent has also shown good activity against different Gram-negative microorganisms.3,26,28 In fact, AZM currently plays a relevant role in the treatment of Campylobacter spp.,28,9 and its use has been recommended in the treatment of other diarrheogenic bacteria, such as E. coli or Shigella spp. 9 Nonetheless, despite the absence of an established breakpoint value, recent reports have shown the presence of microorganisms showing MIC-levels higher than 64 mg/l or in which no halo was detected using AZM discs, and then exhibiting high levels of AZM resistance.16,24
To date, different molecular mechanisms have been described in the development of resistance to AZM. The most important are probably those encoded in mobile elements, such as ere(A), ere(B), mph(A), among others. 16 However, other mechanisms, such as punctual mutations in the rplD (L4), rplV (L22), and rrlH (23 rRNA) genes as well as the potential of AZM to act as a substrate of some chromosomal efflux pumps have also been described.15,19,32
Considering that antibiotic resistance is an emerging problem that can lead to the inefficiency of antimicrobial agents and therapeutic failure, 11 development of antimicrobial resistance should be monitored and the mechanisms underlying this resistance should be investigated.
The aim of this study was to obtain in vitro AZM-resistant E. coli and Shigella mutants and analyze the ability to acquire chromosomal mechanisms of AZM resistance, calculate the rate of mutation, analyze the stability of the mutants obtained and the cross resistance with other agents.
Materials and Methods
Bacterial strains
Three clinical isolates of enterotoxigenic E. coli (E. coli 1- LT; E. coli 2- LT/ST; E. coli 3- LT) and one Shigella boydii with an AZM-MIC ranging between 1 and 4 mg/l were isolated from feces collected from children under 5 years of age with diarrhea in Lima, Peru.
Antimicrobial susceptibility testing
The MICs of AZM, ampicillin, chloramphenicol, nalidixic acid and tetracycline were determined by the agar dilution method in accordance with the CLSI guidelines. 5
Antimicrobial susceptibility testing in the presence of Phe-Arg-β-Naphthylamide
The MICs of the aforementioned antibiotics were determined as described, by adding 20 mg/l of Phe-Arg-β-Naphthylamide (PAβN). The effect of this concentration of PAβN on the viability of microorganisms was also assessed.
Development of AZM-resistant mutants
The isolates were grown in the Luria-Bertani Broth overnight and 100 μl were inoculated onto several Mueller-Hinton plates containing serial dilutions of AZM from their MIC to 64 mg/l. The plates were incubated at 37°C and read at 48 hr. The experiments were done in duplicate. In parallel, serial dilutions were performed to determine the original inoculum. In all cases, the rate of mutation was established as the mean of both experiments as previously described.
10
Briefly, the rate of mutation is the number of colonies growing in the selective plates, counted and divided by the number of plates and the inoculum, calculated following the formula:
In plates where no mutants have grown, the rate of mutation was the reciprocal of the inoculum.
The presence of contaminations was discarded by the study of clonal relations between parental and derived mutant strains by repetitive extragenic palindromic-polymerase chain reaction (PCR) as described elsewhere. 22 From these plates, 16 different colonies (13 E. coli and 3 S. boydii) were randomly selected for further characterization.
Stability of the selected AZM-resistant mutants
To establish the AZM resistance stability of the mutants, the selected mutant strains were cultured in the Mueller-Hinton agar in the absence of antibiotic pressure for 20 cultures (20 consecutive days). 29 After these 20 cultures, the MIC to AZM was performed as aforementioned.
Ribosomal target gene amplification and DNA sequencing
The rplD and rplV genes were amplified both in parental and mutant strains by PCR using the primers listed in Table 1, as previously described by Chittum et al. 4 with the following modifications in the amplification cycle: 94°C, 45 sec; 56°C, 45 sec; 72°C, 2 min; 30 cycles. On the other hand, the 23S rRNA gene (rrlH) was studied in all parental and mutant strains using the specific primers listed in Table 1 and the conditions described by Haanperä et al. 15 PCR products were recovered with the Wizard SV Gel and PCR Clean Up System (Promega, Madison, WI) following the manufacturer's instructions and sequenced (Macrogen, Seoul, Korea).
Outer Membrane Protein analysis
The proteins were extracted according to the methodology previously described in Tavio et al. 30 The outer membrane proteins (OMPs) were visualized in a sodium dodecyl sulfate–polyacrylamide gel electrophoresis at 12% in the following conditions: 150V and 40 mA. OMPs in which differences were observed in acrylamide gels, were recovered and sent for sequencing (University of Barcelona) and their expression levels were determined by Real-Time PCR (RT-PCR). Thus, overnight cultures were diluted 1/100 into 15 ml LB medium and were grown at 37°C until strains reached OD600 values of 0.4 to 0.6. Then, 9 ml were treated with 12 ml of RNAprotect Bacteria reagent and total RNA was extracted with the RNeasy Mini Kit (Qiagen, Hilden, Germany). RNA samples were treated with DNA-free DNase (Ambion, Austin, TX) according to the manufacturer's instructions until RNA samples were totally DNA free. Reverse transcription was performed using the Superscript III First-Strand synthesis system (Eugene, OR) with an input of 3.5 μg of total RNA to a final volume of 30 μl under standard reverse transcription PCR conditions following the manufacturer's instruction. The relative copy number of each target gene was determined in an ABI Prism 7500 Real-Time system (Applied Biosystems, Warrington, United Kingdom), using primers targeting ompA and ompW genes and the rpoB gene, the housekeeping gene used as internal control (Table 1). Reactions were performed in duplicate in a final volume of 20 μl, including 5 μl of cDNA and 10 μl of Power Syber Green Master Mix (Applied Biosystems, Warrington, United Kingdom). The nontemplate control was performed in duplicate for each gene. The cycling conditions were 50°C for 2 min, 95°C for 10 min, and 40 cycles at 95°C for 15 sec and 60°C for 1 min. Data acquisition was done at the end of the elongation stage. Data were analyzed using the 7500 System SDS software (version 1.4; Applied Biosystems).
Results
All the selected mutant strains shown the same REP-PCR pattern as their parental isolate (data not shown), being considered as reliably derived AZM-resistant mutants.
The rate of mutation ranged between < 2.70×10−10 and 2.17×10−7 for E. coli and < 9.58×10−10 to 1.05×10−8 for S. boydii isolates (Table 2). E. coli mutants shown an increase in the AZM-MIC with respect to the parental isolates of 1- to more than sixfold, with one strain (Ec 3.5) achieving an MIC > 256 mg/l (Table 3). The S. boydii mutants only presented increases of a maximum of twofold in MIC levels when compared with parental isolates.
Inoculum total=no plates×inoculum on each plate.
Concentration of AZM (mg/l) in selection plates.
An example for E. coli 2 [AZM]=32 mg/l:
C, confluent growth; - Not performed; E. coli, Escherichia coli; S. boydii, Shigella boydii; AZM, azithromycin; MIC, minimal inhibitory concentration.
The MIC of parental isolates is indicated for comparison purposes.
MIC1, MIC in the first passage; PAβN, MIC performed in presence of PAβN; MIC20, MIC in the passage number 20; PAβN, Phe-Arg-β-Naphthylamide; OMP, outer membrane protein.
Despite showing a final high level of AZM resistance, the mutant Ec 3.5 presented a decrease in the MIC value higher than twofold (from > 256 mg/l to 128 mg/l). The remaining strains shown stable AZM resistance (Table 3).
The viability of microorganisms was not altered by the presence of 20 mg/l of PAβN. No differences in the AZM-MIC levels of the parental and Shigella spp. mutants were observed when PAβN was added, while in the E. coli mutants no MIC returned to the parental levels.
Regarding the other antimicrobial agents tested, no differences were observed in the MIC levels between the parental isolates and their respective derived mutants (Table 4).
In all cases, no differences in the MIC levels among parental isolates and its derived mutants were observed.
AMP, ampicillin; NA, nalidixic acid; C, chloramphenicol; TE, tetracycline.
None of the strains studied presented mutations in the rplV, rplD, and rrlH genes.
No differences were observed in the OMP profiles among the mutant strains derived from E. coli 2 and E. coli 3 and their respective parental isolates, whereas some differences were observed among the parental isolates E. coli 1 and their derived mutants in which the OmpA protein (37.2 kDa) was overexpressed. Thus, levels of expression of OmpA in Ec 1.1, Ec 1.2, and Ec 1.3 were 2-fold, 1.6-fold, and 2.4-fold higher than in the parental strain, respectively. On the other hand, a 1.7-fold increase in the expression level of OmpW (22.9 kDa) was observed in one S. boydii-derived mutant (Sb1) compared with the parental strain (Fig. 1).
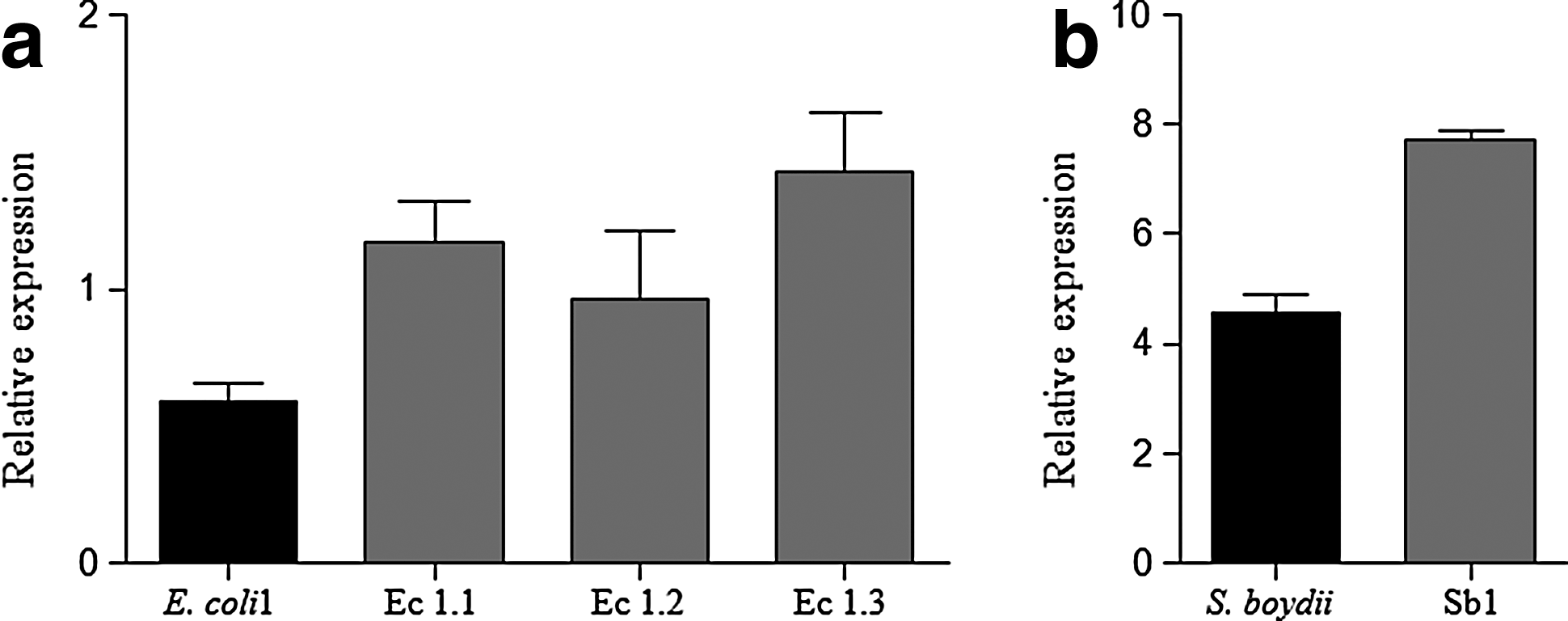
Relative expression of ompA
Discussion
Antibiotic resistance is a relevant factor that affects the usefulness and may diminish the useful life of antimicrobial agents. Taking this into account, studies have been designed to analyze and characterize clinical isolates and in vitro macrolide- resistant mutants of different microorganisms.8,14,32,33
In this study, the ability of AZM to select resistant mutants of S. boydii and E. coli was analyzed. Similar mutation rates were found between the species studied. However, the results show that highly resistant mutants were selected in a single step more easily in E. coli than in Shigella. Although the presence of mutations in the rplV and rplD genes might be a possible explanation, 4 no mutation was detected.
Similarly, specific amino acid substitutions in 23S rRNA have been related to the development of resistance to different antimicrobial agents, such as macrolide, linezolid, or chloramphenicol. However, in the present study, no mutation was found in this gene. The absence of mutations may be related to the presence of multiple copies of the rrlH gene in the chromosome of E. coli, which makes the acquisition of macrolide resistance mediated by this mechanism difficult. 7
PAβN is an efflux pump inhibitor, which has been extensively used to study the role of efflux pumps in the acquisition of antibiotic resistance. PAβN acts as an efflux pump substrate competing with antibacterial agents for its specific binding point. Thus, PAβN does not act against all antibiotics pumped out by the same efflux pump because the antibiotics have different binding points. Macrolides are one of the antibacterial agents that compete with PAβN. 19 Accordingly the effect of PAβN-inhibitible efflux pumps was detected both in parental and mutant E. coli and Shigella spp. strains. However, this did not explain the acquired resistance levels in the E. coli mutants, and the decrease in the MIC of AZM in the presence of PAβN may be attributable to the basal effect of this kind of efflux pumps as presented in the parental isolates. Furthermore, the MIC levels of the E. coli mutant to AZM did not return to parental levels in the presence of PAβN. This fact, together with the stability of AZM-resistant E. coli mutants after 20 passages in media free of antibiotic pressure, supports the possible presence of mutations in other targets, such as stable deregulation of efflux pumps unable to be inhibited by PAβN and which do not affect any of the other drugs investigated.
In some mutants, the initial MIC was slightly lower than the AZM concentration present in the selection plates. This may be attributable to the simultaneous effect of high inoculum used to generate mutants that may allow the bacteria to grow slightly over their real MIC and to express a possible unstable mechanism of resistance (i.e. hyper expression of efflux pumps or permeability alterations) that may revert after the mutant colony is picked up from the selection plate and recultured to perform the MIC.
The mutant strain, Ec 3.2, presents an intriguing AZM resistance phenotype, with a slight increase in the MIC levels after 20 passages in antibiotic-free media. Interestingly, in E. coli, it has recently been described that in the absence of selective pressure, mutations producing low levels of fluoroquinolone resistance are associated with high-fitness cost, but favor the selection of further mutations accounting for a significantly increased fitness, while dramatically decreasing drug susceptibility. 21 Studies to establish if a similar phenomenon might explain the increase of MIC levels from 8 to 32 mg/l in mutant Ec 3.2 are under development.
An increase in the expression level of OmpA was observed in the E. coli 1-derived mutants. To our knowledge, no information about relation between the increase in the levels of this porin and resistance to AZM has been described; so, further studies are needed to determine a possible relationship.
Although differences in the expression levels of OmpW has been related with alterations in the susceptibility levels to different agents,12,17 to our knowledge, this is the first report relating alterations in the expression of OmpW with variations in the susceptibility levels of AZM. However, further studies designed to elucidate the exact role of OmpW in the acquisition of AZM are necessary.
Although studies involving a larger number of strains are necessary, the low rate of selection of AZM-resistant mutants, together with the lack of cross resistance with unrelated antimicrobial agents favor the use of AZM as an alternative treatment for Shigellosis, similar to what has been proposed by other authors. 9 Currently, in Peru, AZM is among the antibiotics recommended for the treatment of children's diarrhea due to Shigella, 13 despite descriptions of highly resistant AZM Enterobacteriaceae clinical isolates. 24 This latter fact is probably due to the presence of AZM plasmid-mediated mechanisms of resistance in the area, as has been described for other resistance determinants.23,25
In summary, the obtained results suggest the selection of a stable unusual mechanism of resistance, and then further studies are being designed to establish it in the present mutants. Besides, it has described the different effects of AZM in the selection of resistant mutants of E. coli and S. boydii. Although AZM is a promising alternative to current treatment schedules for diarrhea in children, periodic surveillance is required to detect the possible appearance of clinical isolates exhibiting resistance. Further studies to elucidate the mechanisms of resistance are under development.
Footnotes
Acknowledgments
We thank Laura Puyol and Diana Barrios for their support in the laboratory management. This work was supported by the Agencia Española de Cooperacion Internacional al Desarrollo (AECID) grants number: D/024648/09, D/030509/10, and A1/035720/11.
JR is supported by program I3 (CES11/012) from Ministerio de Economia and Competitividad (Spain).
Disclosure Statement
The authors have no competing interests to disclose.